Constitutive and Cold Acclimation-Regulated Protein Expression Profiles of Scots Pine Seedlings Reveal Potential for Adaptive Capacity of Geographically Distant Populations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
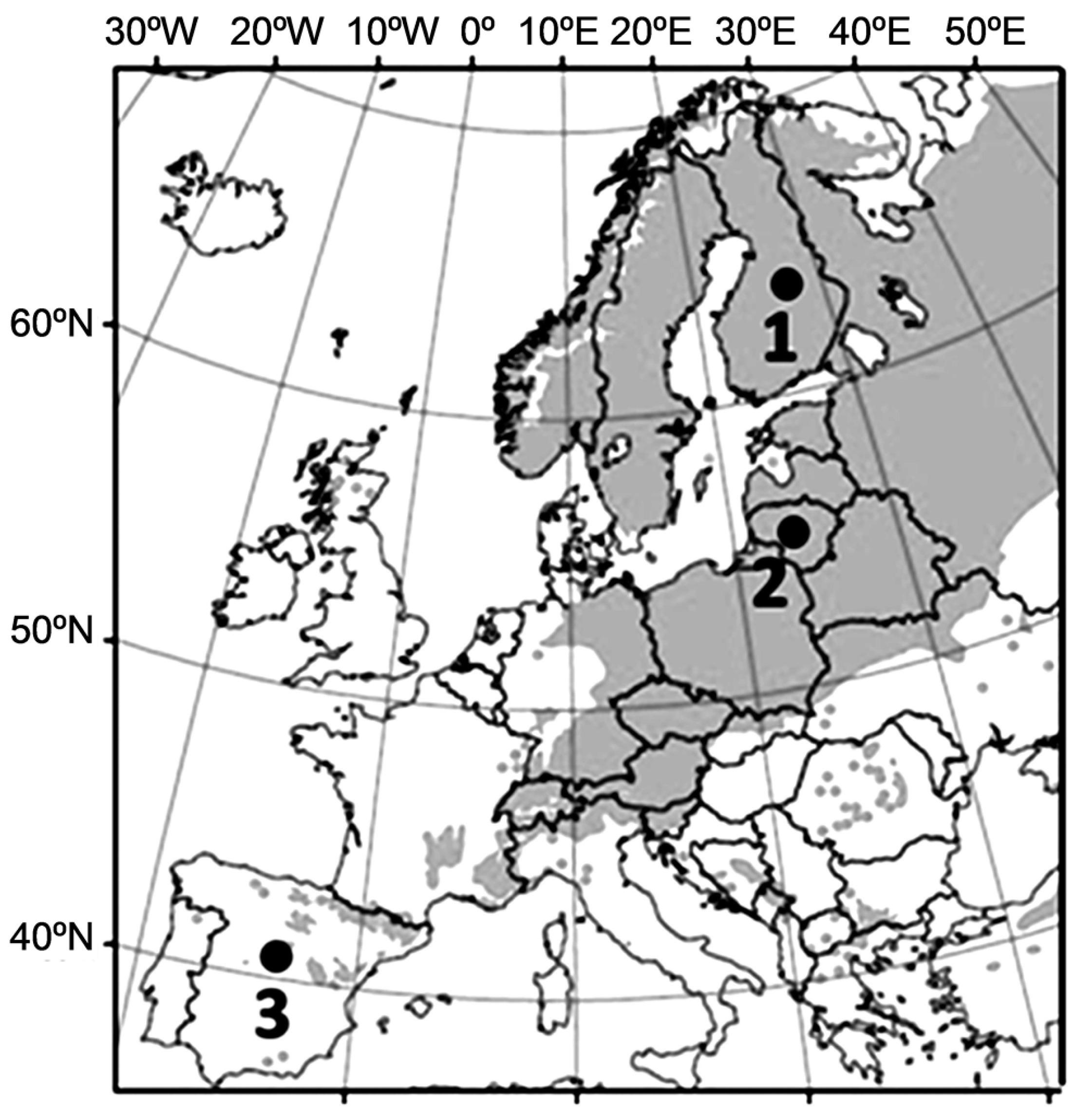
2.1. Seedling Production and Cold Acclimation Treatment
2.2. Seedling Morphology Measurements
2.3. Two-Dimensional Gel Electrophoresis and Protein Identification
2.4. Data Analysis
3. Results
3.1. Seedling Morphology
3.2. Differentially Expressed Proteins
3.3. Protein Expression Patterns
4. Discussion
4.1. Stress Response Proteins Upregulated in the Northern Populations under Active Growth Conditions
4.2. Stress Response Proteins Upregulated in the Lithuanian and/or Spanish Population under Active Growth Conditions
4.3. Effect of Acclimation Treatment on Protein Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Accession | Mascot Score /p. no. /SC, % | Thr. M.W. /pI | Sequence Description | Protein Expression Profile |
|---|---|---|---|---|---|
| 5 | PITA_000033197-RA a | 413 /7 /35.6 | 17.8 /6.2 | pathogenesis-related 10 (PR-10) protein |  |
| 6 | PITA_000003453-RA a | 725 /8 /28.3 | 32.3 /4.8 | 29 kda ribonucleoprotein (CP29) chloroplastic-like |  |
| 9 | PITA_000000569-RA | 1258 /19 /32.8 | 64.4 /5.2 | heat shock cognate 70 kda protein 2 (HSP 70-2) |  |
| 10 | PITA_000048817-RA | 1513 /23 /42.6 | 55.8 /4.8 | protein disulfide-isomerase (PDI) |  |
| 15 | PITA_000069617-RA a | 231 /5 /35.8 | 16.3 /5.9 | nucleoside diphosphate kinase 1 (NDK 1) |  |
| 20 | PITA_000066768-RA a | 348 /5 /33.6 | 25.1 /5.0 | thaumatin-like partial |  |
| 27 | PITA_000060113-RA a | 479 /9 /25.9 | 49.8 /5.7 | calreticulin-3-like isoform x1 |  |
| 29 | PITA_000042977-RA a | 828 /12 /35.9 | 37.7 /5.8 | malate dehydrogenase (MDH) |  |
| 32 | PITA_000037611-RA a | 802 /11 /41.0 | 30.7 /6.2 | proteasome subunit beta type-5 (PSMB5) |  |
| 36 | PITA_000004184-RA | 601 /8 /17.6 | 57.6 /7.2 | alcohol dehydrogenase partial |  |
| 37 | PITA_000055561-RA a | 926 /15 /36.1 | 34.9 /5.0 | 40s ribosomal protein sa-like |  |
| 39 | PITA_000052060-RA | 1085 /18 /24.0 | 101.2 /5.7 | chaperone protein 1 |  |
| 42 | PITA_000009823-RA a | 429 /8 /28.4 | 35.1 /4.9 | Ser/Thr-protein phosphatase pp2a (PP2A) catalytic subunit |  |
| 48 | PITA_000019887-RA a | 477 /10 /28.9 | 44.9 /5.6 | guanosine-diphosphate (GDP)-mannose -epimerase 2 |  |
| 52 | PITA_000011094-RA a | 673 /10 /15.1 | 105 /6.0 | probable linoleate 9s-lipoxygenase (LOX) 5 |  |
| 53 | PITA_000020461-RA a | 912 /19 /20.0 | 111.7 /6.4 | aconitase cytoplasmic |  |
| 55 | PITA_000016807-RA a | 619 /11 /40.8 | 29.9 /4.2 | tubulin beta chain-like |  |
| 60 | PITA_000006003-RA a | 599 /12 /25.0 | 32.4 /6.7 | tau class glutathione s-transferase (GST) |  |
| 61 | PITA_000059572-RA a | 1289 /18 /41.0 | 56.2 /5.4 | protein Tic62 chloroplastic |  |
| 64 | PITA_000047327-RA a | 383 /7 /23.5 | 43.5 /6.0 | oxygen-evolving enhancer (OEE) protein chloroplastic |  |
| 65 | PITA_000055142-RA a | 566 /9 /31.2 | 30.5 /8.8 | nicotinamide adenine dinucleotide (NAD)-binding Rossmann-fold superfamily protein |  |
| 67 | PITA_000008754-RA a | 328.69 /4 /21.9 | 23.6 /5.1 | abscisic acid water deficit stress and ripening (ASR) inducible-like partial |  |
| 68 | PITA_000022327-RA | 446 /5 /50.0 | 17.6 /5.6 | water deficit inducible lipoprotein 3 (LP3)-like partial |  |
| 73 | PITA_000023770-RA a | 448 /9 /23.9 | 32.6 /5.0 | lactoylglutathione lyase |  |
| 75 | PITA_000008205-RA a | 768 /14 /58.0 | 22.8 /8.8 | PR-10 protein |  |
| 76 | PITA_000077110-RA a | 157 /3 /22.6 | 10.6 /5.1 | cyanate hydratase |  |
References
- Paoletti, E.; Bytnerowicz, A.; Andersen, C.; Augustaitis, A.; Ferretti, M.; Grulke, N.; Gunthardt-Goerg, M.S.; Innes, J.; Johnson, D.; Karnosky, D.; et al. Impacts of air pollution and climate change on forest ecosystems—Emerging research needs. Sci. World J. 2007, 7 (Suppl. 1), 1–8. [Google Scholar] [CrossRef]
- Perie, C.; de Blois, S. Dominant forest tree species are potentially vulnerable to climate change over large portions of their range even at high latitudes. PeerJ 2016, 4, e2218. [Google Scholar] [CrossRef] [Green Version]
- Aspinwall, M.J.; Loik, M.E.; Resco de Dios, V.; Tjoelker, M.G.; Payton, P.R.; Tissue, D.T. Utilizing intraspecific variation in phenotypic plasticity to bolster agricultural and forest productivity under climate change. Plant Cell Environ. 2014, 38, 1752–1764. [Google Scholar] [CrossRef] [PubMed]
- Neale, D.B.; Ingvarsson, P.K. Population, quantitative and comparative genomics of adaptation in forest trees. Curr. Opin. Plant Biol. 2008, 11, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Breda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Kremer, A.; Ronce, O.; Robledo-Arnuncio, J.J.; Guillaume, F.; Bohrer, G.; Nathan, R.; Bridle, J.R.; Gomulkiewicz, R.; Klein, E.K.; Ritland, K.; et al. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [Green Version]
- Petit, R.J.; Hampe, A. Some evolutionary consequences of being a tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Eriksson, O. Evolutionary forces influencing variation among populations of Pinus sylvestris. Silva Fennica 1998, 32, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Diz, A.P.; Martinez-Fernandez, M.; Rolan-Alvarez, E. Proteomics in evolutionary ecology: Linking the genotype with the phenotype. Mol. Ecol. 2012, 21, 1060–1080. [Google Scholar] [CrossRef] [PubMed]
- Karr, T.L. Application of proteomics to ecology and population biology. Heredity 2008, 100, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockman, M.V.; Kruglyak, L. Genetics of global gene expression. Nat. Rev. Genet. 2006, 7, 862–872. [Google Scholar] [CrossRef]
- Thiellement, H.; Zivy, M.; Plomion, C. Combining proteomic and genetic studies in plants. J. Chromatogr. B 2002, 782, 137–149. [Google Scholar] [CrossRef]
- Chevalier, F.; Martin, O.; Rofidal, V.; Devauchelle, A.D.; Barteau, S.; Sommerer, N.; Rossignol, M. Proteomic investigation of natural variation between Arabidopsis ecotypes. Proteomics 2004, 4, 1372–1381. [Google Scholar] [CrossRef]
- Bedon, F.; Villar, E.; Vincent, D.; Dupuy, J.W.; Lomenech, A.M.; Mabialangoma, A.; Chaumeil, P.; Barre, A.; Plomion, C.; Gion, J.M. Proteomic plasticity of two Eucalyptus genotypes under contrasted water regimes in the field. Plant Cell Environ. 2012, 35, 790–805. [Google Scholar] [CrossRef]
- Valcu, C.M.; Lalanne, C.; Plomion, C.; Schlink, K. Heat induced changes in protein expression profiles of Norway spruce (Picea abies) ecotypes from different elevations. Proteomics 2008, 8, 4287–4302. [Google Scholar] [CrossRef]
- Valcu, C.M.; Lalanne, C.; Muller-Starck, G.; Plomion, C.; Schlink, K. Protein polymorphism between 2 Picea abies populations revealed by 2-dimensional gel rlectrophoresis and tandem mass spectrometry. J. Hered. 2008, 99, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Cheddadi, R.; Vendramin, G.G.; Litt, T.; Francois, L.; Kageyama, M.; Lorentz, S.; Laurent, J.M.; De Beaulieu, J.L.; Sadori, L.; Jost, A.; et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Glob. Ecol. Biogeogr. 2006, 15, 271–282. [Google Scholar] [CrossRef]
- Sinclair, W.T.; Morman, J.D.; Ennos, R. The postglacial history of Scots pine (Pinus sylvestris L.) in Western Europe: Evidence from mitochondrial DNA variation. Mol. Ecol. 1999, 8, 83–88. [Google Scholar] [CrossRef]
- Soranzo, N.; Alia, R.; Provan, J.; Powell, W. Patterns of variation at a mitochondrial sequence-tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations. Mol. Ecol. 2000, 9, 1205–1211. [Google Scholar] [CrossRef]
- Dvornyk, V.; Sirvio, A.; Mikkonen, M.; Savolainen, O. Low nucleotide diversity at the pal1 locus in the widely distributed Pinus sylvestris. Mol. Biol. Evol. 2002, 19, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gil, M.R.; Mikkonen, M.; Savolainen, O. Nucleotide diversity at two phytochrome loci along a latitudinal cline in Pinus sylvestris. Mol. Ecol. 2003, 12, 1195–1206. [Google Scholar] [CrossRef]
- Naydenov, K.; Senneville, S.; Beaulieu, J.; Tremblay, F.; Bousquet, J. Glacial vicariance in Eurasia: Mitochondrial DNA evidence from Scots pine for a complex heritage involving genetically distinct refugia at mid-northern latitudes and in Asia Minor. BMC Evol. Biol. 2007, 7, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyhajarvi, T.; Salmela, M.J.; Savolainen, O. Colonization routes of Pinus sylvestris inferred from distribution of mitochondrial DNA variation. Tree Genet. Genomes 2008, 4, 247–254. [Google Scholar] [CrossRef]
- Willis, K.J.; van Andel, T.H. Trees or no trees? The environments of central and eastern Europe during the last glaciation. Quat. Sci. Rev. 2004, 23, 2369–2387. [Google Scholar] [CrossRef]
- Scalfi, M.; Piotti, A.; Rossi, M.; Piovani, P. Genetic variability of Italian southern Scots pine (Pinus sylvestris L.) populations: The rear edge of the range. Eur. J. For. Res. 2009, 128, 377. [Google Scholar] [CrossRef]
- Hannerz, M. Genetic and Seasonal Variation in Hardiness and Growth Rhythm in Boreal and Temperate Conifers—A Review and Annotated Bibliography; The Forestry Research Institute of Sweden Report; SkogForsk: Uppsala, Sweden, 1998; pp. 1–140. [Google Scholar]
- Wachowiak, W.; Perry, A.; Donnelly, K.; Cavers, S. Early phenology and growth trait variation in closely related European pine species. Ecol. Evol. 2018, 8, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Dietrichson, J. A summary of studies on genetic variation in forest trees grown in Scandinavia with special reference to the adaptation problem. Norway Skogforsoksv Medd 1971, 29, 25–56. [Google Scholar]
- Eriksson, G.; Andersson, S.; Eiche, V.; Ifver, J.; Persson, A. Severity index and transfer effects on survival and volume production of Pinus sylvestris in northern Sweden. Stud. For. Suecica 1980, 156, 1–32. [Google Scholar]
- Strimbeck, G.R.; Schaberg, P.G.; Fossdal, C.G.; Schroder, W.P.; Kjellsen, T.D. Extreme low temperature tolerance in woody plants. Front. Plant Sci. 2015, 6, 884. [Google Scholar] [CrossRef] [PubMed]
- Dormling, I. Bud dormancy, frost hardiness and frost drought in seedlings of Pinus sylvestris and Picea abies. In Advances in Cold Hardiness; Li, P.H., Christersson, L., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 285–298. [Google Scholar]
- Dormling, I. Influence of light intensity and temperature on photoperiodic response of Norway spruce provenances. In Proceedings of the IUFRO Meeting of WP Norway Spruce Provenances (S 2.03.11) and Norway Spruce Breeding (S 2.02.11), Bucharest, Romania, 24 September–1 October 1979; pp. 398–407. [Google Scholar]
- Repo, T. Seasonal changes in frost hardiness in Picea abies and Pinus sylvestris in Finland. Can. J. For. Res. 1992, 22, 1949–1957. [Google Scholar] [CrossRef]
- Wisniewski, M.; Nassuth, A.; Arora, R. Cold hardiness in trees: A mini-review. Front. Plant Sci. 2018, 9, 1394. [Google Scholar] [CrossRef]
- Kontunen-Soppela, S.; Lankila, J.; Lähdesmäki, L.; Laine, K. Response of protein and carbohydrate metabolism of Scots pine seedlings to low temperature. J. Plant Physiol. 2002, 159, 175–180. [Google Scholar] [CrossRef]
- Pukacki, P.M.; Kamińska-Rożek, E. Differential effects of spring reacclimation and deacclimation on cell membranes of Norway spruce seedlings. Acta Soc. Bot. Pol. 2013, 82, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Vogg, G.; Heim, R.; Hansen, J.; Schäfer, C.; Beck, E. Frost hardening and photosynthetic performance of Scots pine (Pinus sylvestris L.) needles. I. Seasonal changes in the photosynthetic apparatus and its function. Planta 1998, 204, 193–200. [Google Scholar] [CrossRef]
- Vogg, G.; Heim, R.; Gotschy, B.; Beck, E.; Hansen, J. Frost hardening and photosynthetic performance of Scots pine (Pinus sylvestris L.) needles. II. Seasonal changes in the fluidity of thylakoid membranes. Planta 1998, 204, 201–206. [Google Scholar] [CrossRef]
- Pukacki, P.M.; Kamińska-Rożek, E. Reactive species, antioxidants and cold tolerance during deacclimation of Picea abies populations. Acta Physiol. Plant. 2013, 35, 129–138. [Google Scholar] [CrossRef]
- Bae, J.J.; Choo, Y.S.; Ono, K.; Sumida, A.; Hara, T. Photoprotective mechanisms in cold-acclimated and nonacclimated needles of Picea glehnii. Photosynthetica 2010, 48, 110–116. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, Q.; Xu, M.; Chi, Y.; Shen, R.; Li, P.; Dai, H. Responses of Pinus massoniana and Pinus taeda to freezing in temperate forests in central China. Scand. J. For. Res. 2012, 27, 520–531. [Google Scholar] [CrossRef]
- Korotaeva, N.; Romanenko, A.; Suvorova, G.; Ivanova, M.V.; Lomovatskaya, L.; Borovskii, G.; Voinikov, V. Seasonal changes in the content of dehydrins in mesophyll cells of common pine needles. Photosynth. Res. 2015, 124, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Basset, C.I.; Gusta, L.V. An overview of cold hardiness in woody plants: Seeing the forest through the trees. Hortscience 2003, 38, 952–959. [Google Scholar] [CrossRef]
- Danusevicius, D.; Gabrilavicius, R. Variation in juvenile growth rhythm among Picea abies provenances from the Baltic states and adjacent regions. Scand. J. For. Res. 2001, 16, 305–317. [Google Scholar] [CrossRef]
- Isaacson, T.; Damasceno, C.M.; Saravanan, R.S.; He, Y.; Catala, C.; Saladie, M.; Rose, J.K. Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat. Protoc. 2006, 1, 769–774. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tamosiune, I.; Staniene, G.; Haimi, P.; Stanys, V.; Rugienius, R.; Baniulis, D. Endophytic Bacillus and Pseudomonas spp. modulate apple shoot growth, cellular redox balance, and protein expression under in vitro conditions. Front. Plant Sci. 2018, 9, 889. [Google Scholar] [CrossRef] [Green Version]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Neale, D.B.; Wegrzyn, J.L.; Stevens, K.A.; Zimin, A.V.; Puiu, D.; Crepeau, M.W.; Cardeno, C.; Koriabine, M.; Holtz-Morris, A.E.; Liechty, J.D.; et al. Decoding the massive genome of loblolly pine using haploid DNA and novel assembly strategies. Genome Biol. 2014, 15, R59. [Google Scholar] [CrossRef] [Green Version]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, K.; Fischer, B.B.; Madureira, D.J.; Pillai, S. Transcriptomics in ecotoxicology. Anal. Bioanal. Chem. 2010, 397, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigras, F.J.; Ryyppo, A.; Lindstrom, A.; Stattin, E. Cold acclimation and deacclimation of shoots and roots of conifer seedlings. In Conifer Cold Hardiness; Bigras, F.J., Colombo, S.J., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 57–88. [Google Scholar]
- Agarwal, M.; Katiyar-Agarwal, S.; Grover, A. Plant Hsp100 proteins: Structure, function and regulation. Plant Sci. 2002, 163, 397–405. [Google Scholar] [CrossRef]
- Singla, S.L.; Pareek, A.; Grover, A. Yeast HSP 104 homologue rice HSP 110 is developmentally- and stress-regulated. Plant Sci. 1997, 125, 211–219. [Google Scholar] [CrossRef]
- Hong, S.W.; Vierling, E. Hsp101 is necessary for heat tolerance but dispensable for development and germination in the absence of stress. Plant J. 2001, 27, 25–35. [Google Scholar] [CrossRef]
- Thelin, L.; Mutwil, M.; Sommarin, M.; Persson, S. Diverging functions among calreticulin isoforms in higher plants. Plant Signal Behav. 2011, 6, 905–910. [Google Scholar] [CrossRef] [Green Version]
- Ndimba, B.K.; Chivasa, S.; Simon, W.J.; Slabas, A.R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 4185–4196. [Google Scholar] [CrossRef]
- Komatsu, S.; Yang, G.; Khan, M.; Onodera, H.; Toki, S.; Yamaguchi, M. Over-expression of calcium-dependent protein kinase 13 and calreticulin interacting protein 1 confers cold tolerance on rice plants. Mol. Genet. Genom. 2007, 277, 713–723. [Google Scholar] [CrossRef]
- Stone, S.L. The role of ubiquitin and the 26S proteasome in plant abiotic stress signaling. Front. Plant Sci. 2014, 5, 135. [Google Scholar] [CrossRef]
- Garcia-Hernandez, M.; Davies, E.; Baskin, T.I.; Staswick, P.E. Association of plant p40 protein with ribosomes is enhanced when polyribosomes form during periods of active tissue growth. Plant Physiol. 1996, 111, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Omidbakhshfard, M.A.; Omranian, N.; Ahmadi, F.S.; Nikoloski, Z.; Mueller-Roeber, B. Effect of salt stress on genes encoding translation-associated proteins in Arabidopsis thaliana. Plant Signal Behav. 2012, 7, 1095–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzlovar, S.; Dermastia, M. The comparative analysis of osmotins and osmotin-like PR-5 proteins. Plant Biol. 2003, 5, 116–124. [Google Scholar] [CrossRef]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef]
- Zhu, B.; Chen, T.H.; Li, P.H. Expression of an ABA-responsive osmotin-like gene during the induction of freezing tolerance in Solanum commersonii. Plant Mol. Biol. 1993, 21, 729–735. [Google Scholar] [CrossRef]
- Liu, J.J.; Ekramoddoullah, A.K.M. The family 10 of plant pathogenesis-related proteins: Their structure, regulation, and function in response to biotic and abiotic stresses. Physiol. Mol. Plant Pathol. 2006, 68, 3–13. [Google Scholar] [CrossRef]
- Ukaji, N.; Kuwabara, C.; Takezawa, D.; Arakawa, K.; Fujikawa, S. Accumulation of pathogenesis-related (PR) 10/Bet v 1 protein homologues in mulberry (Morus bombycis Koidz.) tree during winter. Plant Cell Environ. 2004, 27, 1112–1121. [Google Scholar] [CrossRef]
- Christie, P.J.; Hahn, M.; Walbot, V. Low-temperature accumulation of alcohol dehydrogenase-1 mRNA and protein activity in maize and rice seedlings. Plant Physiol. 1991, 95, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Jarillo, J.A.; Leyva, A.; Salinas, J.; Martinez-Zapater, J.M. Low temperature induces the accumulation of alcohol dehydrogenase mRNA in Arabidopsis thaliana, a chilling-tolerant plant. Plant Physiol. 1993, 101, 833–837. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, M.; Temple, S.J.; Allan, D.L.; Vance, C.P.; Samac, D.A. Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiol. 2001, 127, 1836–1844. [Google Scholar] [CrossRef]
- Yao, Y.X.; Dong, Q.L.; Zhai, H.; You, C.X.; Hao, Y.J. The functions of an apple cytosolic malate dehydrogenase gene in growth and tolerance to cold and salt stresses. Plant Physiol. Biochem. 2011, 49, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Nveawiah-Yoho, P.; Zhou, J.; Palmer, M.; Sauve, R.; Zhou, S. Identification of proteins for salt tolerance using a comparative proteomics analysis of tomato accessions with contrasting salt tolerance. J. Am. Soc. Hortic. Sci. 2013, 138, 382–394. [Google Scholar] [CrossRef] [Green Version]
- Askari, H.; Edqvist, J.; Hajheidari, M.; Kafi, M.; Salekdeh, G.H. Effects of salinity levels on proteome of Suaeda aegyptiaca leaves. Proteomics 2006, 6, 2542–2554. [Google Scholar] [CrossRef]
- Miller, J.M.; Conn, E.E. Metabolism of hydrogen cyanide by higher plants. Plant Physiol. 1980, 65, 1199–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiser, G.D.; Wang, T.T.; Hoffman, N.E.; Yang, S.F.; Liu, H.W.; Walsh, C.T. Formation of cyanide from carbon 1 of 1-aminocyclopropane-1-carboxylic acid during its conversion to ethylene. Proc. Natl. Acad. Sci. USA 1984, 81, 3059–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conklin, P.L.; Norris, S.R.; Wheeler, G.L.; Williams, E.H.; Smirnoff, N.; Last, R.L. Genetic evidence for the role of GDP-mannose in plant ascorbic acid (vitamin C) biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 4198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid—A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Liu, Z.; Fan, M.; Chen, Y.; Tian, H.; Wu, D.; Gao, H.; Ren, C.; Song, S.; Xie, D. GDP-D-mannose epimerase regulates male gametophyte development, plant growth and leaf senescence in Arabidopsis. Sci. Rep. 2017, 7, 10309. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Shen, G.; Yan, J.; He, C.; Zhang, H. AtCHIP functions as an E3 ubiquitin ligase of protein phosphatase 2A subunits and alters plant response to abscisic acid treatment. Plant J. 2006, 46, 649–657. [Google Scholar] [CrossRef]
- Monroy, A.; Sangwan, V.; Dhindsa, R.S. Low temperature signal transduction during cold acclimation: Protein phosphatase 2A as an early target for cold-inactivation. Plant J. 1998, 13, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Pais, S.M.; Gonzalez, M.A.; Tellez-Inon, M.T.; Capiati, D.A. Characterization of potato (Solanum tuberosum) and tomato (Solanum lycopersicum) protein phosphatases type 2A catalytic subunits and their involvement in stress responses. Planta 2009, 230, 13–25. [Google Scholar] [CrossRef]
- Kalapos, M.P. The tandem of free radicals and methylglyoxal. Chem. Biol. Interact. 2008, 171, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Droog, F. Plant glutathione S-transferase, a tale of theta and tau. J. Plant Growth Regul. 1997, 16, 95–107. [Google Scholar] [CrossRef]
- Roxas, V.P.; Smith, R.K., Jr.; Allen, E.R.; Allen, R.D. Overexpression of glutathione S-transferase/glutathione peroxidase enhances the growth of transgenic tobacco seedlings during stress. Nat. Biotechnol. 1997, 15, 988–991. [Google Scholar] [CrossRef]
- Livanos, P.; Galatis, B.; Apostolakos, P. The interplay between ROS and tubulin cytoskeleton in plants. Plant Signal Behav. 2014, 9, e28069. [Google Scholar] [CrossRef]
- Golan, I.; Dominguez, P.G.; Konrad, Z.; Shkolnik-Inbar, D.; Carrari, F.; Bar-Zvi, D. Tomato ABSCISIC ACID STRESS RIPENING (ASR) gene family revisited. PLoS ONE 2014, 9, e107117. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.F.; Yu, S.C.; Yang, C.Y.; Wang, C.S. Lily ASR protein-conferred cold and freezing resistance in Arabidopsis. Plant Physiol. Biochem. 2011, 49, 937–945. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, S.C.; Hong, S.K.; An, K.; An, G.; Kim, S.R. Ectopic expression of a cold-responsive OsAsr1 cDNA gives enhanced cold tolerance in transgenic rice plants. Mol. Cells 2009, 27, 449–458. [Google Scholar] [CrossRef]
- Padmanabhan, V.; Dias, D.M.; Newton, R.J. Expression analysis of a gene family in loblolly pine (Pinus taeda L.) induced by water deficit stress. Plant Mol. Biol. 1997, 35, 801–807. [Google Scholar] [CrossRef]
- Seidler, A.; Rutherford, A.W.; Michel, H. On the role of the N-terminus of the extrinsic 33 kDa protein of Photosystem II. Plant Mol. Biol. 1996, 31, 183–188. [Google Scholar] [CrossRef]
- Riccardi, F.; Gazeau, P.; Jacquemot, M.P.; Vincent, D.; Zivy, M. Deciphering genetic variations of proteome responses to water deficit in maize leaves. Plant Physiol. Biochem. 2004, 42, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, K.; Hanagata, N.; Dubinsky, Z.; Baba, S.; Karube, I. Molecular characterization of cDNA encoding oxygen evolving enhancer protein 1 increased by salt treatment in the mangrove Bruguiera gymnorrhiza. Plant Cell Physiol. 2000, 41, 1279–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Wilkinson, B.; Gilbert, H.F. Protein disulfide isomerase. Biochim. Biophys. Acta 2004, 1699, 35–44. [Google Scholar] [CrossRef]
- Kupsch, C.; Ruwe, H.; Gusewski, S.; Tillich, M.; Small, I.; Schmitz-Linneweber, C. Arabidopsis chloroplast RNA binding proteins CP31A and CP29A associate with large transcript pools and confer cold stress tolerance by influencing multiple chloroplast RNA processing steps. Plant Cell 2012, 24, 4266–4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, M.A.; Ray, N.B.; Olcott, M.C.; Hendricks, S.P.; Mathews, C.K. Metabolic functions of microbial nucleoside diphosphate kinases. J. Bioenerg. Biomembr. 2000, 32, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Ahsan, N.; Lee, S.H.; Kang, K.Y.; Lee, J.J.; Lee, B.H. An approach to identify cold-induced low-abundant proteins in rice leaf. C. R. Biol. 2007, 330, 215–225. [Google Scholar] [CrossRef]
- Krol, M.; Hurry, V.M.; Maxwell, D.P.; Malek, L.; Ivanov, A.G.; Huner, N.P.A. Low growth temperature inhibition of photosynthesis in cotyledons of jack pine seedlings (Pinus banksianna) is due to impaired chloroplast development. Can. J. Bot. 2002, 80, 1042–1051. [Google Scholar] [CrossRef]
- Oquist, G.; Huner, N.P. Photosynthesis of overwintering evergreen plants. Annu. Rev. Plant Biol. 2003, 54, 329–355. [Google Scholar] [CrossRef]




| Trait | Population | ||
|---|---|---|---|
| Spanish | Lithuanian | Finnish | |
| Number of cotyledons (4 weeks after germination) | 7.0 ± 0.12 a | 6.0 ± 0.13 b | 6.0 ± 0.13 b |
| Seedling height before the acclimation treatment (12 weeks after seeding), mm | 58.6 ± 0.23 a | 63.6 ± 0.23 a | 31.9 ± 0.11 b |
| Length of hypocotyl before the acclimation treatment (12 weeks after seeding), mm | 30.2 ± 0.16 a | 30.8 ± 0.16 a | 17.0 ± 0.06 b |
| Needle weight before the acclimation treatment (12 weeks after seeding), g | 4.2 ± 0.51 a | 3.2 ± 0.26 b | 1.5 ± 0.06 c |
| Needle weight at the end of acclimation treatment (14 weeks after seeding), g | 4.5 ± 0.16 a | 4.7 ± 0.14 a | 2.2 ± 0.34 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baniulis, D.; Sirgėdienė, M.; Haimi, P.; Tamošiūnė, I.; Danusevičius, D. Constitutive and Cold Acclimation-Regulated Protein Expression Profiles of Scots Pine Seedlings Reveal Potential for Adaptive Capacity of Geographically Distant Populations. Forests 2020, 11, 89. https://doi.org/10.3390/f11010089
Baniulis D, Sirgėdienė M, Haimi P, Tamošiūnė I, Danusevičius D. Constitutive and Cold Acclimation-Regulated Protein Expression Profiles of Scots Pine Seedlings Reveal Potential for Adaptive Capacity of Geographically Distant Populations. Forests. 2020; 11(1):89. https://doi.org/10.3390/f11010089
Chicago/Turabian StyleBaniulis, Danas, Monika Sirgėdienė, Perttu Haimi, Inga Tamošiūnė, and Darius Danusevičius. 2020. "Constitutive and Cold Acclimation-Regulated Protein Expression Profiles of Scots Pine Seedlings Reveal Potential for Adaptive Capacity of Geographically Distant Populations" Forests 11, no. 1: 89. https://doi.org/10.3390/f11010089
APA StyleBaniulis, D., Sirgėdienė, M., Haimi, P., Tamošiūnė, I., & Danusevičius, D. (2020). Constitutive and Cold Acclimation-Regulated Protein Expression Profiles of Scots Pine Seedlings Reveal Potential for Adaptive Capacity of Geographically Distant Populations. Forests, 11(1), 89. https://doi.org/10.3390/f11010089