To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
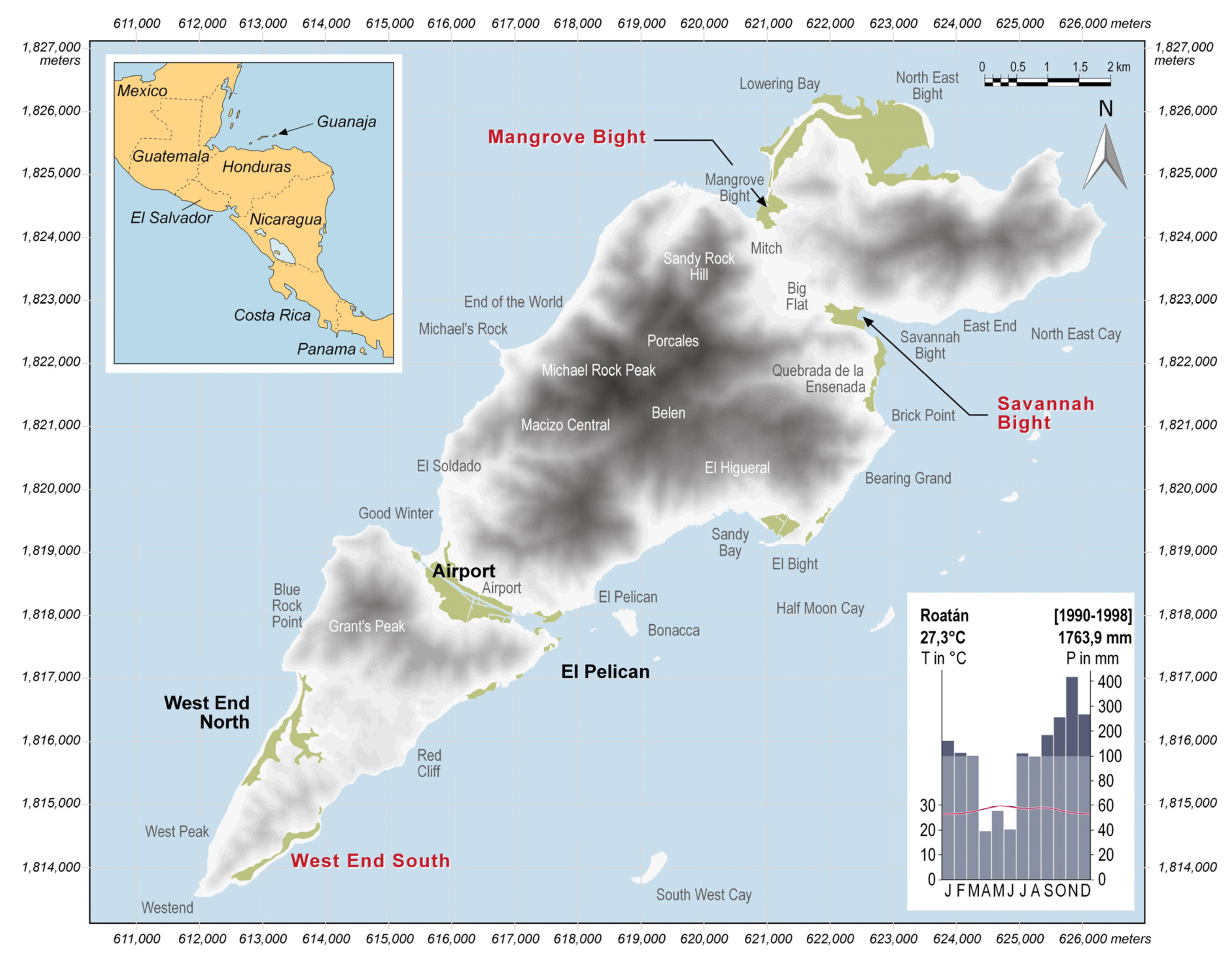
2.1. Study Area
2.2. Field Sampling
2.3. Data Analysis
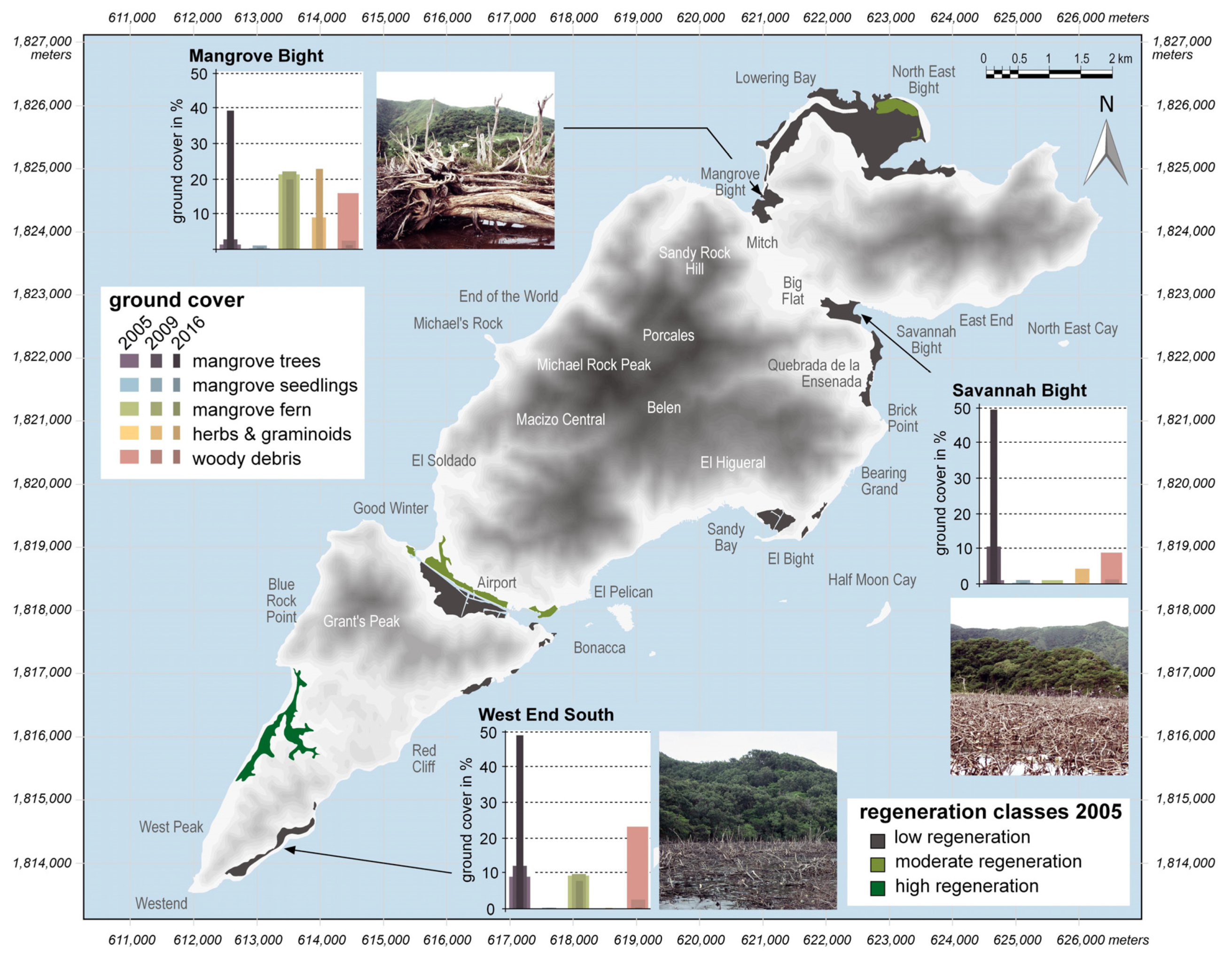
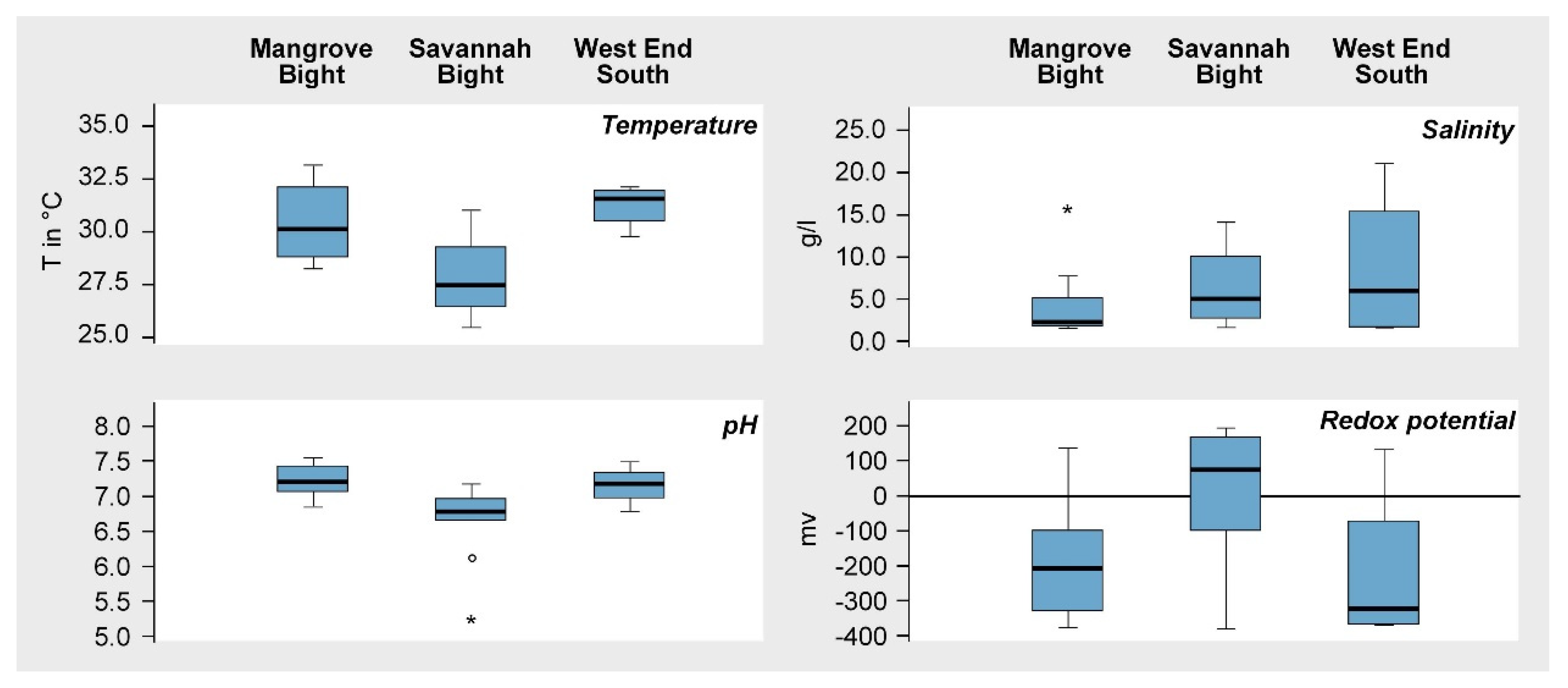
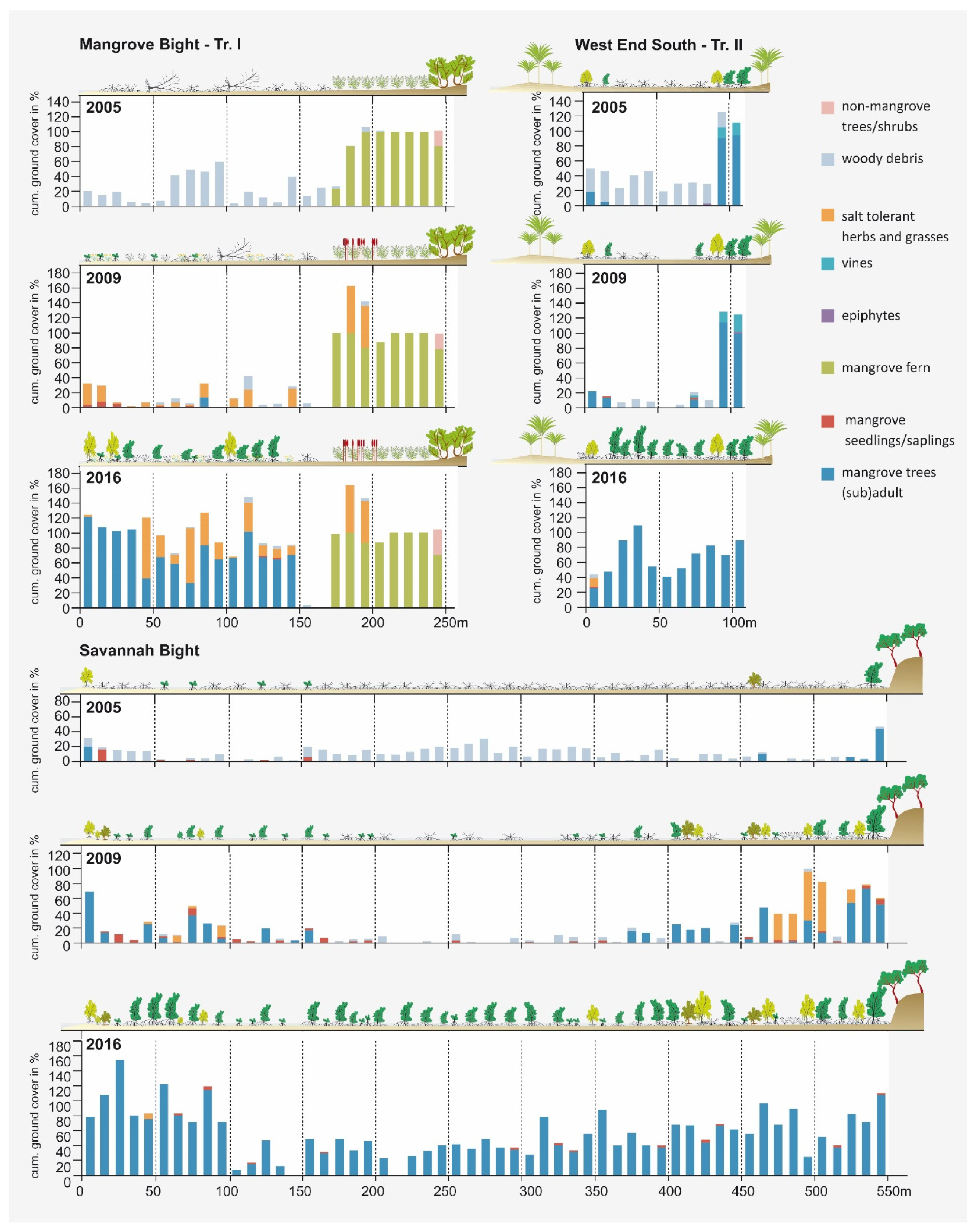
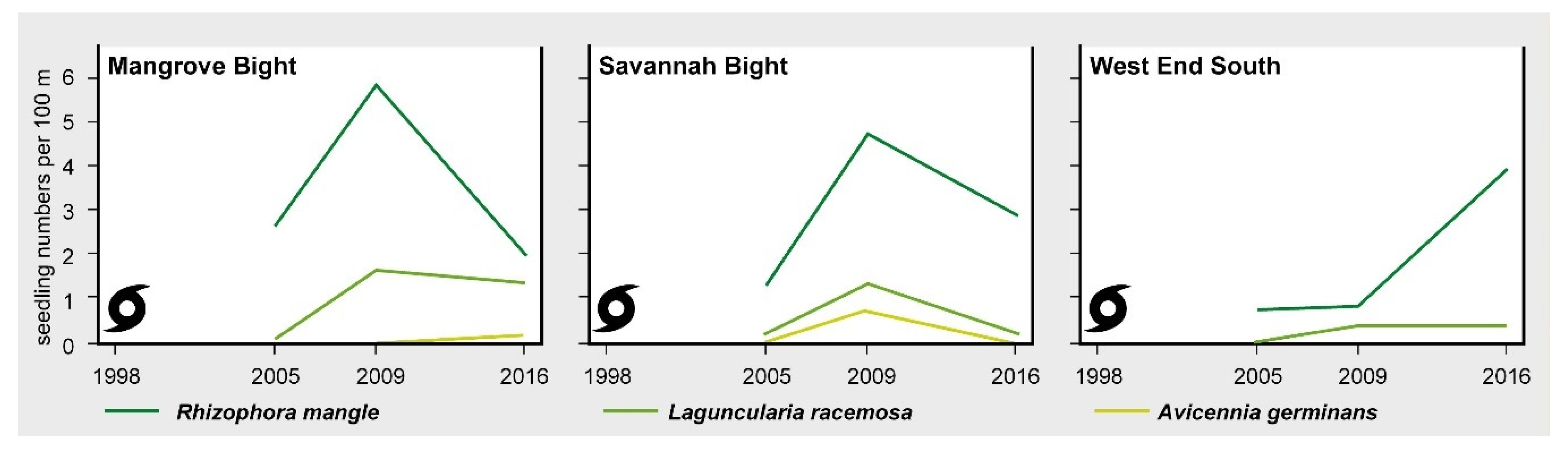
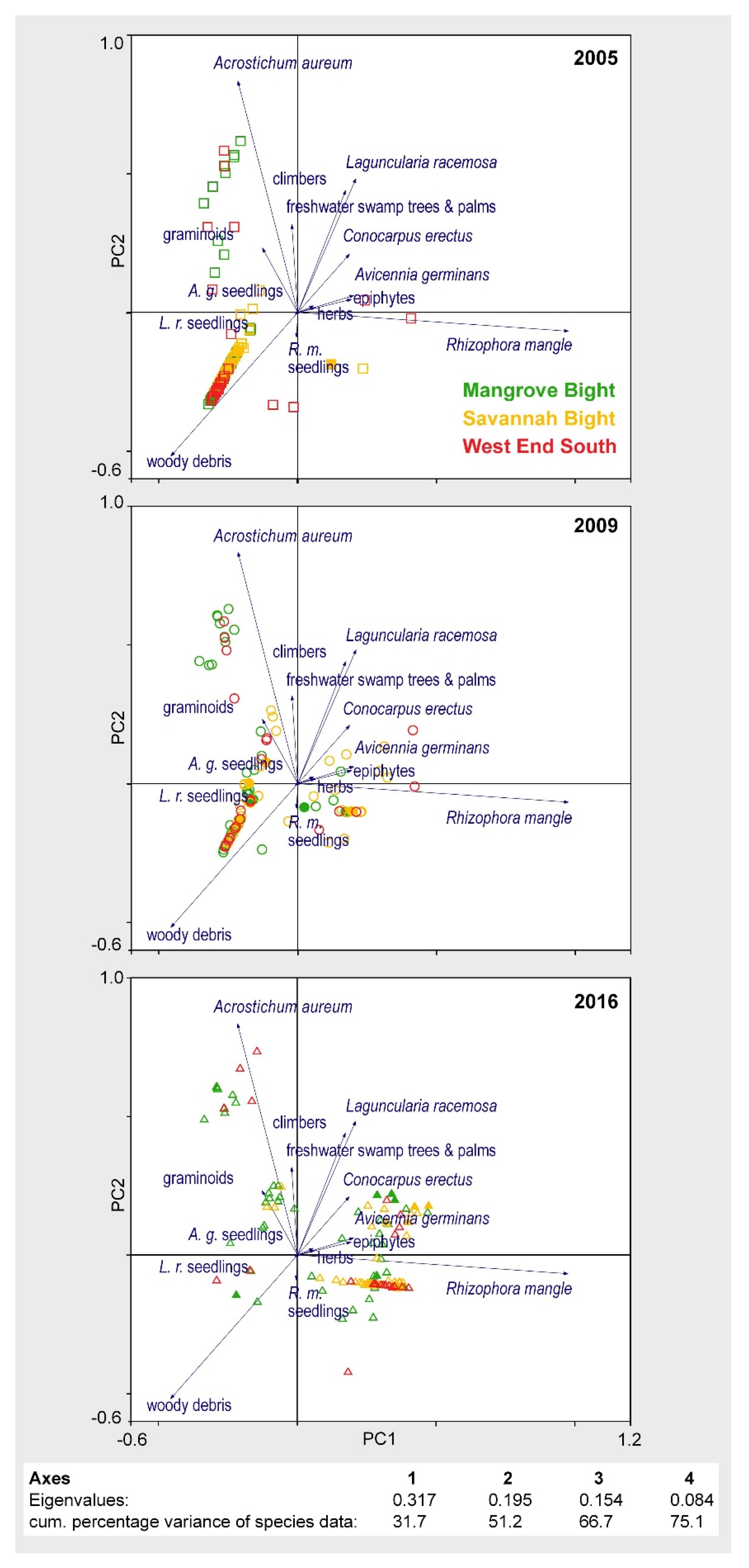
3. Results
4. Discussion
4.1. Mangrove Regeneration and Restoration on Guanaja after Hurricane Mitch
- (1)
- whether no reforestation measures were undertaken, as in WES,
- (2)
- whether some planting was successful as in parts of SB and MB, or
- (3)
- whether an interim herbaceous phase was involved, as in MB.
4.2. Mangrove Regeneration and Restoration—A More Global Perspective
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Ewel, K.C.; Twilley, R.R.; Ong, J.E. Different kinds of mangrove forests provide different goods and services. Glob. Ecol. Biogeogr. Lett. 1998, 7, 83–94. [Google Scholar] [CrossRef]
- Walters, B.B.; Rönnbäck, P.; Kovacs, J.M.; Crona, B.; Hussain, S.A.; Badola, R.; Primavera, J.H.; Barbier, E.; Dahdouh-Guebas, F. Ethnobiology, socioeconomics and management of mangrove forests: A review. Aquat. Bot. 2008, 89, 220–236. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.; Kainuma, M.; Collins, L. World Atlas of Mangroves; Earthscan: London, UK; Washington, DC, USA, 2010. [Google Scholar]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Salem, M.E.; Mercer, D.E. The economic value of mangroves: A meta-analysis. Sustainability 2012, 4, 359–383. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.L.; et al. Ecological role and services of tropical mangrove ecosystems: A reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- UNEP. The Importance of Mangroves to People: A Call to Action; van Bochove, J., Sullivan, E., Nakamura, T., Eds.; United Nations Environment Programme World Conservation Monitoring Centre: Cambridge, UK, 2014. [Google Scholar]
- Cummings, A.R.; Shah, M. Mangroves in the global climate and environmental mix. Geogr. Compass 2017, e12353. [Google Scholar] [CrossRef]
- Huxham, M.; Dencer-Brown, A.; Diele, K.; Kathiresan, K.; Nagelkerken, I.; Wanjiru, C. Mangroves and people: Local ecosystem Services in a changing climate. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 245–274. [Google Scholar]
- Barbier, E.B.; Koch, E.M.; Silliman, B.R.; Hacker, S.D.; Wolanski, E.; Primavera, J.; Granek, E.F.; Polasky, S.; Aswani, S.; Cramer, L.A.; et al. Coastal Ecosystem-Based Management with Nonlinear Ecological Functions and Values. Science 2008, 319, 321–323. [Google Scholar] [CrossRef]
- Costanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Marois, D.E.; Mitsch, W.J. Coastal protection from tsunamis and cyclones provided by mangrove wetlands—A review. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 71–83. [Google Scholar] [CrossRef]
- Danielsen, F.; Sørensen, M.K.; Olwig, M.F.; Selvam, V.; Parish, F.; Burgess, N.D.; Hiraishi, T.; Karunagaran, V.M.; Rasmussen, M.S.; Hansen, L.B.; et al. The Asian Tsunami: A protective role for coastal vegetation. Science 2005, 310, 643. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, K.; Rajendran, N. Coastal mangrove forests mitigated tsunami. Estuar. Coast. Shelf Sci. 2005, 65, 601–606. [Google Scholar] [CrossRef]
- Barbier, E.B. Natural barriers to natural disasters: Replanting mangroves after the tsunami. Front. Ecol. Environ. 2006, 4, 124–131. [Google Scholar] [CrossRef]
- Barbier, E.B. The protective service of mangrove ecosystems: A review of valuation methods. Mar. Pollut. Bull. 2016, 109, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Vincent, J.R. Mangroves protected villages and reduced death toll during Indian super cyclone. PNAS 2009, 106, 7357–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandilyan, S.; Kathiresan, K. Mangroves as bioshield: An undisputable fact. Ocean Coast. Manag. 2005, 103, 94–96. [Google Scholar] [CrossRef]
- Hochard, J.P.; Hamilton, S.; Barbier, E.B. Mangroves shelter coastal economic activity from cyclones. PNAS 2019, 116, 12232–12237. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.; McIvor, A.; Tonneijck, F.H.; Tol, S.; van Eijk, P. Mangroves for coastal defence. In Guidelines for Coastal Managers & Policy Makers; Wetlands International, Wageningen, Neherlands, and The Nature Conservancy: Arlington, TX, USA, 2014; p. 42. [Google Scholar]
- Krauss, K.W.; Osland, M.J. Tropical cyclones and the organization of mangrove forests: A review. Ann. Bot. 2020, 125, 213–234. [Google Scholar] [CrossRef]
- Walton, M.; Samonte-Tan, G.; Primavera, J.; Edwards-Jones, G.; Le Vay, L. Are mangroves worth replanting? The direct economic benefits of a community-based reforestation project. Environ. Conserv. 2006, 33, 335–343. [Google Scholar] [CrossRef]
- Primavera, J.H.; Rollon, R.N.; Samson, M.S. The pressing challenges of mangrove rehabilitation: Pond reversion and coastal protection. In Ecohydrology and Restoration; Chicharo, L., Zalewski, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 217–244. [Google Scholar]
- Andradi-Brown, D.A.; Howe, C.; Mace, G.M.; Knight, A.T. Do mangrove forest restoration or rehabilitation activities return biodiversity to pre-impact levels? Environ. Evid. 2013, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- López-Portillo, J.; Lewis III, R.R.; Saenger, P.; Rovai, A.; Koedam, N.; Dahdouh-Guebas, F.; Agraz-Hernández, C.; Rivera-Monroy, V.H. Mangrove Forest Restoration and Rehabilitation. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 301–345. [Google Scholar]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [Green Version]
- Ellison, A.M.; Felson, A.J.; Friess, D.A. Mangrove Rehabilitation and Restoration as Experimental Adaptive Management. Front. Mar. Sci. 2020, 7, 327. [Google Scholar] [CrossRef]
- Ellison, A.M. Mangrove Restoration: Do we know enough? Restor. Ecol. 2000, 8, 219–229. [Google Scholar] [CrossRef]
- Iftekhar, M.S. Functions and development of reforested mangrove areas: A review. Int. J. Biodivers. Sci. Manag. 2008, 4, 1–14. [Google Scholar] [CrossRef]
- IUCN Mass Mangrove Restoration: Driven by Good Intentions But Offering Limited Results. 2017. Available online: https://www.iucn.org/news/forests/201702/mass-mangrove-restoration-driven-good-intentions-offering-limited-results (accessed on 10 May 2020).
- Kodikara, K.A.S.; Mukherjee, N.; Jayatissa, L.P.; Dahdouh-Guebas, F.; Koedam, N. Have mangrove restoration projects worked? An in-depth study in Sri Lanka. Restor. Ecol. 2017, 25, 705–716. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hamilton, S.; Barbier, E.B.; Primavera, J.; Lewis, R.R. Better restoration policies are needed to conserve mangrove ecosystems. Nat. Ecol. Evol. 2019, 3, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.S.; Ruiz, P.L.; Telesnicki, G.J.; Meeder, J.F. Estimating above-ground biomass and production in mangrove communities of Biscayne national Park, Florida (USA). Wetl. Ecol. Manag. 2001, 9, 27–37. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Onrizal, O.; Ahmad, A.G.; Mansour, M. Assessment of Natural Regeneration of Mangrove Species at Tsunami Affected Areas in Indonesia and Malaysia. IOP Conf. Ser. Mater. Sci. Eng. 2017, 180, 012045. [Google Scholar] [CrossRef]
- Bayraktarov, E.; Saunders, M.I.; Abdullah, S.; Mills, M.; Beher, J.; Possingham, H.P.; Mumby, P.J.; Lovelock, C.E. The cost and feasibility of marine coastal restoration. Ecol. Appl. 2016, 26, 1055–1074. [Google Scholar] [CrossRef]
- Fickert, T. Better resilient than resistant—Regeneration Dynamics of storm-disturbed mangrove forests on the Bay Island of Guanaja (Honduras) during the first two decades after Hurricane Mitch (October 1998). Diversity 2018, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Pasch, R.J.; Avila, L.A.; Guiney, J.L. Atlantic Hurricane Season of 1998. Mon. Weather Rev. 2001, 129, 3085–3123. [Google Scholar] [CrossRef] [Green Version]
- Cahoon, D.R.; Hensel, P.; Rybczyk, J.; McKee, K.; Proffitt, C.E.; Perez, B. Mass tree mortality leads to mangrove peat collapse at Bay Islands, Honduras after Hurricane Mitch. J. Ecol. 2003, 91, 1093–1105. [Google Scholar] [CrossRef]
- Guiney, J.L.; Lawrence, M.B. Preliminary Report: Hurricane Mitch 22 October–5 November 1998; National Hurricane Center: Miami, FL, USA, 1999. [Google Scholar]
- Vanselow, K.A.; Kolb, M.; Fickert, T. Destruction and Regeneration of Terrestrial, Littoral and Marine Ecosystems on the Island of Guanaja/Honduras seven years after Hurricane Mitch. Erdkunde 2007, 61, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Fickert, T.; Grüninger, F. Floristic zonation, vegetation structure, and plant diversity patterns within a Caribbean mangrove and swamp forest on the Bay Island of Utila (Honduras). Ecotropica 2010, 16, 73–92. [Google Scholar]
- Valade, I.; Grelot, J.; Laurent, J.M. Esquema Director de Manejo Ambietal de las Islas de la Bahía, Honduras; Repiublica de Honduras: Islas de La Bahia, Honduras, 2002. [Google Scholar]
- Doyle, T.W.; Michot, T.C.; Roetker, F.; Sullivan, J.; Melder, M.; Handley, B.; Balmat, J. Hurricane Mitch: Landscape Analysis of Damaged Forest Resources of the Bay Islands and Caribbean Coast of Honduras, USGS Open File Report 03-175. 2002.
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Cintrón, G.; Schaeffer-Novelli, Y. Methods for studying mangrove structure. In The Mangrove Ecosystem: Research Methods; Snedaker, S.C., Snedaker, J.G., Eds.; UNESCO: Paris, France, 1984; pp. 91–113. [Google Scholar]
- Balick, M.J.; Nee, M.H.; Daniel, E.A. Checklist of the Vascular Plants of Belize; Memoirs of the New York Botanical Garden 85: New York, NY, USA, 2000. [Google Scholar]
- Ter Braak, C.J.F.; Šmilaur, P. CANOCO for Windows Version 4.5; Biometris, Plant Research International: Wageningen, The Netherlands, 2002. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Saenger, P. Mangrove Ecology, Silviculture and Conservation; Kluwer: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2002. [Google Scholar]
- Vanselow, K.A. Die Ruhe nach dem Sturm—Eine Zeitreihenanalyse zur Entwicklung der Vegetationsbedeckung auf der Karibikinsel Guanaja nach Hurrikan Mitch. Mitt. Fränkischen Geogr. Ges. 2018, 63, 111–121. [Google Scholar]
- Winograd, M. Sustainability and vulnerability indicators for decision making: Lessons learned from Honduras. Int. J. Sustain. Dev. 2007, 10, 93–105. [Google Scholar] [CrossRef]
- Wanless, H.R.; Parkinson, R.W.; Tedesco, L.P. Sea level control on stability of Everglades wetlands. In Everglades: The Ecosystem and Its Restoration; Davis, S.M., Ogden, J.C., Eds.; St. Lucie Press: Boca Raton, FL, USA, 1994; pp. 199–223. [Google Scholar]
- Bandeira, S.; Balidy, H. Limpopo Estuary Mangrove Transformation, Rehabilitation and Management. In Estuaries: A Lifeline of Ecosystem Services in the Western Indian Ocean; Diop, S., Scheren, P., Machiwa, J., Eds.; Springer: Heidelberg/Berlin, Germany, 2016; pp. 227–237. [Google Scholar]
- Mbense, S.; Rajkaran, A.; Bolosha, U.; Adams, J. Rapid colonization of degraded mangrove habitat by succulent salt marsh. S. Afr. J. Bot. 2016, 107, 129–136. [Google Scholar] [CrossRef]
- Asbridge, E.; Lucas, R.; Rogers, K.; Accad, A. The extent of mangrove change and potential for recovery following severe Tropical Cyclone Yasi, Hinchinbrook Island, Queensland, Australia. Ecol. Evol. 2018, 8, 10416–10434. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer-Novelli, Y.; Soriano-Sierra, E.; Vale, C.; Bernini, E.; Rovai, A.; Pinheiro, M.; Schmidt, A.; de Almeida, R.; Júnior, C.; Menghini, R.; et al. Climate changes in mangrove forests and salt marshes. Braz. J. Oceanogr. 2016, 64, 37–52. [Google Scholar] [CrossRef] [Green Version]
- McKee, K.L.; McGinnis II, T.E. Hurricane Mitch: Effects on Mangrove Soil Characteristics and Root Contributions to Soil Stabilization; USGS Open File Report 03-178: Lafayette, IN, USA, 2002. [Google Scholar]
- Lacerda, L.D. Mangrove Ecosystems: Function and Management; Springer: Berlin, Germany, 2002. [Google Scholar]
- Hossain, M.D.; Nuruddin, A.A. Soil and mangrove: A Review. J. Environ. Sci. Technol. 2016, 9, 198–207. [Google Scholar] [CrossRef] [Green Version]
- McDonald, K.O.; Webber, D.F.; Webber, M.K. Mangrove forest structure under varying environmental conditions. Bull. Mar. Sci. 2003, 73, 491–505. [Google Scholar]
- Hseu, Z.Y.; Chen, Z.S. Monitoring the changes of redox potential, pH and electric conductivity of the mangrove soils in Northern Taiwan. Proc. Natl. Sci. Counc. Repub. China Part B 2000, 24, 143–150. [Google Scholar]
- Imbert, D.; Rousteau, A.; Scherrer, P. Ecology of Mangrove Growth and Recovery in the Lesser Antilles: State of Knowledge and Basis for Restoration Projects. Restor. Ecol. 2000, 8, 230–236. [Google Scholar] [CrossRef]
- Milbrandt, E.C.; Tinsley, M.N. The role of saltwort (Batis maritima L.) in regeneration of degraded mangroive forests. Hydrobiologia 2006, 568, 369–377. [Google Scholar] [CrossRef]
- McKee, K.L.; Rooth, J.E.; Feller, I.C. Mangrove recruitment after forest disturbance is facilitated by herbaceous species in the Caribbean. Ecol. Appl. 2007, 17, 1678–1693. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Urrego, L.E.; Martínez, J.I.; Polanía, J.; Yokoyama, Y. Mangrove dynamics in the southwestern Caribbean since the ‘Little Ice Age’: A history of human and natural disturbances. Holocene 2010, 20, 849–861. [Google Scholar] [CrossRef]
- Biswas, S.R.; Khan, M.S.I.; Mallik, A.U. Invaders’ control on post-disturbance succession in coastal mangroves. J. Plant Ecol. 2012, 5, 157–166. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove extinction Risk and Geographic Areas of global concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Chowdhury, R.R.; Uchida, E.; Chen, L.; Osorio, V.; Yoder, L. Anthropogenic Drivers of Mangrove Loss. Patterns and Implications for Livelihoods. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 275–300. [Google Scholar]
- Feller, I.C.; Friess, D.A.; Krauss, K.W.; Lewis III, R.R. The state of the world’s mangroves in the 21st century under climate change. Hydrobiologia 2017, 803, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.; Lucas, R.; Bunting, P.; Hardy, A.; Rosenqvist, A.; Simard, M. Distribution and drivers of global mangrove forest change, 1996–2010. PLoS ONE 2017, 12, e0179302. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.R. Ecological engineering for successful management and restoration of mangrove forests. Ecol. Eng. 2005, 24, 403–418. [Google Scholar] [CrossRef]
- Dale, P.E.R.; Knight, J.M.; Dwyer, P.G. Mangrove rehabilitation: A review focusing on ecological and institutional issues. Wetl. Ecol. Manag. 2014, 22, 587–604. [Google Scholar] [CrossRef] [Green Version]
- Rey, J.R.; Carlson, D.B.; Brockmeyer Jr., R. E. Coastal wetland management in Florida: Environmental concerns and human health. Wetl. Ecol. Manag. 2012, 20, 197–211. [Google Scholar] [CrossRef]
- Brown, B.; Fadilla, R.; Nurdin, Y.; Soulsby, I.; Ahmad, R. Community based ecological mangrove rehabilitation (CBEMR) in Indonesia. Sapiens 2014, 7, 53–64. [Google Scholar]
- Ferreira, A.C.; Ganade, G.; de Attayde, J.L. Restoration versus natural regeneration in a neotropical mangrove: Effects on plant biomass and crab communities. Ocean Coast. Manag. 2015, 110, 38–45. [Google Scholar] [CrossRef]
- Bosire, J.O.; Dahdouh-Guebas, F.; Walton, M.; Crona, B.I.; Lewis, R.R.; Field, C.; Kairo, J.G.; Koedam, N. Functionality of restored mangroves: A review. Aquat. Bot. 2008, 89, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Primavera, J.; Esteban, J. A review of mangrove rehabilitation in the Philippines: Successes, failures and future prospects. Wetl. Ecol. Manag. 2008, 16, 345–358. [Google Scholar] [CrossRef]
- Barnuevo, A.; Asaeda, T.; Sanjaya, K.; Kanesaka, Y.; Fortes, M. Drawbacks of mangrove rehabilitation schemes: Lessons learned from the large-scale mangrove plantations. Estuar. Coast. Shelf Sci. 2017, 198, 432–437. [Google Scholar] [CrossRef]
- Rovai, A.; Soriano-Sierra, E.; Pagliosa, P.; Cintrón, G.; Schaeffer-Novelli, Y.; Menghini, R.; Coelho-Jr, C.; Horta, P.; Lewis, R.; Simonassi, J.; et al. Secondary succession impairment in restored mangroves. Wetl. Ecol. Manag. 2012, 20, 447–459. [Google Scholar] [CrossRef]
- Granado, R.; Pinto Neta, L.C.; Nunes-Freitas, A.F.; Voloch, C.M.; Lira, C. Assessing Genetic Diversity after Mangrove Restoration in Brazil: Why Is It So Important? Diversity 2018, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, R.B.; Yeo-Chang, Y.; Park, M.S.; Chun, J.N. Local People’s Participation in Mangrove Restoration Projects and Impacts on Social Capital and Livelihood: A Case Study in the Philippines. Forests 2020, 11, 580. [Google Scholar] [CrossRef]
- Babak, K.; Hashim, R. Mangrove restoration without planting. Ecol. Eng. 2011, 37, 387–391. [Google Scholar]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönig, D. Resilience-Based Restoration of Mangrove Social-Ecological Systems—A Case Study from the Gulf of Guayaquil, Ecuador. Master’s Thesis, Master thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Forest Ecology and Management, University Freiburg, Freiburg, Germany, 2014. [Google Scholar]
- Romañach, S.; Deangelis, D.; Koh, H.; Li, Y.; Teh, S.; Sulaiman, R.; Zhai, L. Conservation and restoration of mangroves: Global status, perspectives, and prognosis. Ocean Coast. Manag. 2018, 154, 72–82. [Google Scholar] [CrossRef]






© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fickert, T. To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras). Forests 2020, 11, 1068. https://doi.org/10.3390/f11101068
Fickert T. To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras). Forests. 2020; 11(10):1068. https://doi.org/10.3390/f11101068
Chicago/Turabian StyleFickert, Thomas. 2020. "To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras)" Forests 11, no. 10: 1068. https://doi.org/10.3390/f11101068
APA StyleFickert, T. (2020). To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras). Forests, 11(10), 1068. https://doi.org/10.3390/f11101068