Selection and Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis in Needles of Larix olgensis under Abiotic Stresses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA Extraction and cDNA Preparation
2.3. Selection of Candidate Reference Genes and Primer Design
2.4. Reverse Transcription Quantitative PCR (qRT-PCR) Analysis
2.5. Gene Expression Stability Analysis
2.6. Validation of RGs by qRT-PCR
3. Results
3.1. Selection of Candidate Reference Genes
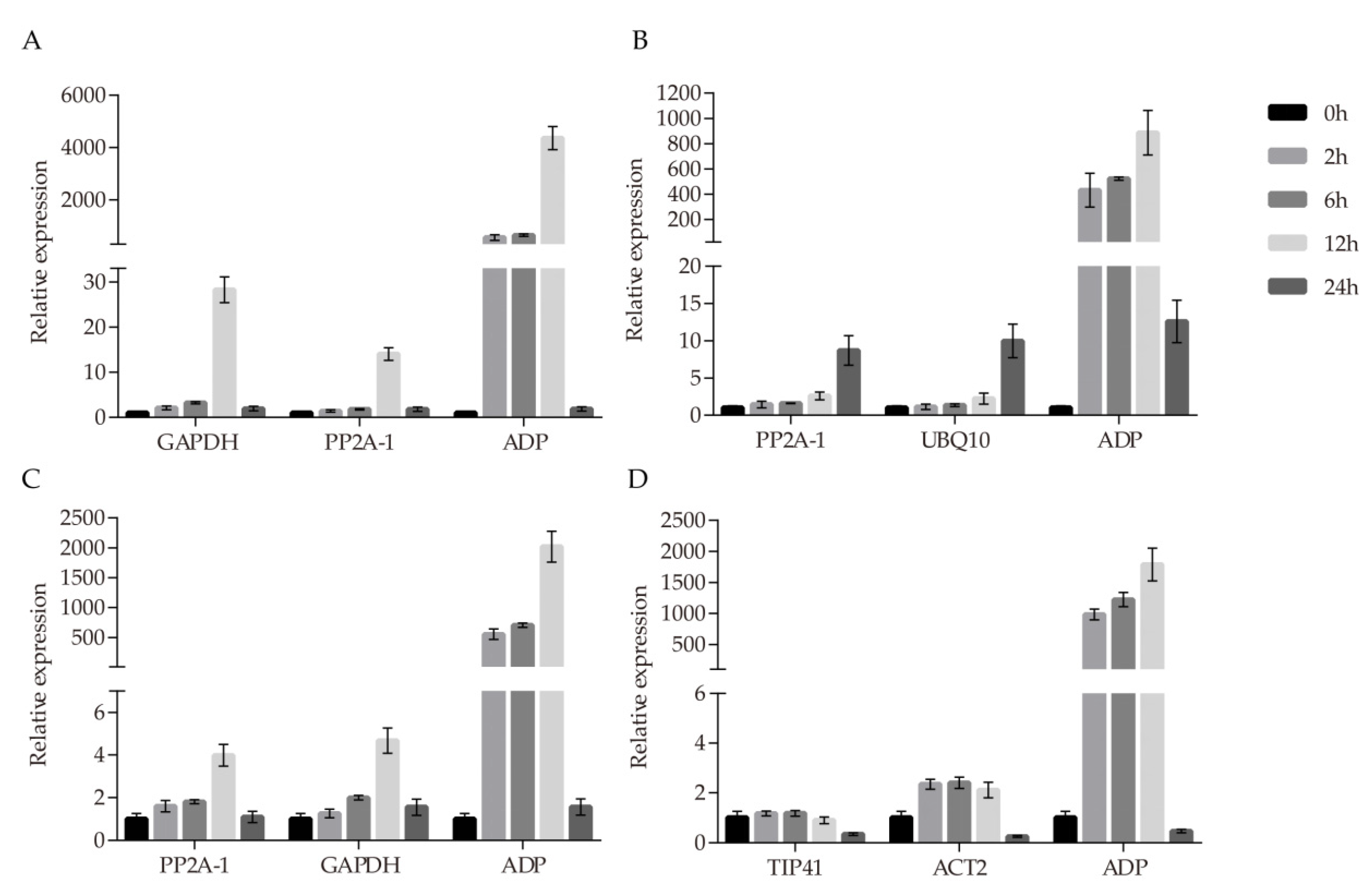
3.2. Expression Profiles of Candidate RGs
3.3. Analysis of Gene Expression Stability
3.4. Comprehensive Analysis
3.5. Validation of CAT Reference Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miao, L.; Qin, X.; Gao, L.; Li, Q.; Li, S.; He, C.; Li, Y.; Yu, X. Selection of reference genes for quantitative real-time PCR analysis in cucumber (Cucumis sativus L.), pumpkin (Cucurbita moschata Duch.) and cucumber-pumpkin grafted plants. PeerJ 2019, 7, e6536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 532–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dheda, K.; Huggett, J.F.; Bustin, S.A.; Johnson, M.A.; Rook, G.; Zumla, A. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques 2004, 37, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Z. Expression profiling of ABA pathway transcripts indicates crosstalk between abiotic and biotic stress responses in Arabidopsis. Genomics 2012, 100, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Mascia, T.; Santovito, E.; Gallitelli, D.; Cillo, F. Evaluation of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in infected tomato plants. Mol. Plant Pathol. 2010, 11, 805–816. [Google Scholar] [CrossRef]
- Zhou, Z.; Cong, P.; Tian, Y.; Zhu, Y. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots. PLoS ONE 2017, 12, e185288. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, B.; Su, X.; Zhang, S.; Huang, M. Reference gene selection for quantitative real-time polymerase chain reaction in Populus. Anal. Biochem. 2011, 408, 337–339. [Google Scholar] [CrossRef]
- Guenin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: the necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.T.; Poolakkalody, N.J.; Shah, J.M. Plant reference genes for development and stress response studies. J. Biosci. 2018, 43, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Sgamma, T.; Pape, J.; Massiah, A.; Jackson, S. Selection of reference genes for diurnal and developmental time-course real-time PCR expression analyses in lettuce. Plant Methods 2016, 12, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Song, X.; Wang, F.; Xiong, A.S. Suitable reference genes for accurate gene expression analysis in parsley (Petroselinum crispum) for abiotic stresses and hormone stimuli. Front. Plant Sci. 2016, 7, 1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Han, X.; Chen, S.; Zheng, L.; He, X.; Liu, M.; Qiao, G.; Wang, Y.; Zhuo, R. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Liu, J.X.; Xing, G.M.; Sun, S.; Li, S.; Duan, A.Q.; Wang, F.; Li, M.Y.; Xu, Z.S.; Xiong, A.S. Selection of appropriate reference genes for RT-qPCR analysis under abiotic stress and hormone treatment in celery. PeerJ 2019, 7, e7925. [Google Scholar] [CrossRef]
- Ma, S.; Niu, H.; Liu, C.; Zhang, J.; Hou, C.; Wang, D. Expression stabilities of candidate reference genes for RT-qPCR under different stress conditions in soybean. PLoS ONE 2013, 8, e0075271. [Google Scholar] [CrossRef] [Green Version]
- Wan, Q.; Chen, S.; Shan, Z.; Yang, Z.; Chen, L.; Zhang, C.; Yuan, S.; Hao, Q.; Zhang, X.; Qiu, D.; et al. Stability evaluation of reference genes for gene expression analysis by RT-qPCR in soybean under different conditions. PLoS ONE 2017, 12, e0189405. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Papierniak, A. Identification of suitable reference genes for studying gene expression in cucumber plants subjected to abiotic stress and growth regulators. Mol. Breed. 2011, 28, 343–357. [Google Scholar] [CrossRef]
- Vasyutkina, E.A.; Reunova, G.D.; Tupikin, A.E.; Zhuravlev, Y.N. Mitochondrial DNA variation in Olga Bay larch (Larix olgensis A. Henry) from Primorsky Krai of Russia. Genetika 2014, 50, 291–298. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, K.; Yan, Q.; Liu, Z.; Yu, L.; Wang, H. Feasibility of implementing thinning in even-aged Larix olgensis plantations to develop uneven-aged larch-broadleaved mixed forests. J. For. Res. 2010, 15, 71–80. [Google Scholar] [CrossRef]
- Shen, C.; Wang, L.; Li, M. The altitudinal variability and temporal instability of the climate-tree-ring growth relationships for Changbai larch (Larix olgensis Henry) in the Changbai mountains area, Jilin, Northeastern China. Trees 2016, 30, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Zhang, H.; Zhang, L.; Li, X.; Deng, J.; Jiang, T. Construction of genetic linkage maps of larch (Larix kaempferi × Larix gmelini) by rapd markers and mapping of QTLs for larch. Biotechnol. Biotechnol. Equip. 2011, 25, 2197–2202. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Yang, J.; Li, C. Transcriptomic response to nitric oxide treatment in Larix olgensis Henry. Int. J. Mol. Sci. 2015, 16, 28582–28597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Sun, X.; Xie, Y.; Feng, J.; Zhang, S. Transcriptome and proteome profiling of adventitious root development in hybrid larch (Lari kaempferi × Lari olgensis). BMC Plant Biol. 2014, 14, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Xiong, H.; Wang, J.; Zhang, H.; Zhang, L. Mining Myb transcription factors related to wood development in Larix olgensis. J. For. Res. 2019. [Google Scholar] [CrossRef]
- Li, D.; Hu, X.; Li, C. Overexpression of the LoMYB29 gene of Larix olgensis contributes to the regulation of proanthocyanidin biosynthesis in Arabidopsis thaliana. J. For. Res. 2019, 30, 1793–1804. [Google Scholar] [CrossRef]
- Hu, X.; Xu, X.; Li, C. Ectopic expression of the LoERF017 transcription factor from Larix olgensis Henry enhances salt and osmotic-stress tolerance in Arabidopsis thaliana. Plant Biotechnol. Rep. 2018, 12, 93–104. [Google Scholar] [CrossRef]
- Chen, R.; Chen, W.; Tigabu, M.; Zhong, W.; Li, Y.; Ma, X.; Li, M. Screening and evaluation of stable reference genes for quantitative real-time polymerase chain reaction (qRT-PCR) analysis in Chinese fir roots under water, phosphorus, and nitrogen stresses. Forests 2019, 10, 1087. [Google Scholar] [CrossRef] [Green Version]
- Paschalidis, K.; Tsaniklidis, G.; Wang, B.; Delis, C.; Trantas, E.; Loulakakis, K.; Makky, M.; Sarris, P.F.; Ververidis, F.; Liu, J. The interplay among polyamines and nitrogen in plant stress responses. Plants 2019, 8, 315. [Google Scholar] [CrossRef] [Green Version]
- Nie, Q.; Gao, G.L.; Fan, Q.J.; Qiao, G.; Wen, X.P.; Liu, T.; Peng, Z.J.; Cai, Y.Q. Isolation and characterization of a catalase gene “HuCAT3” from pitaya (Hylocereus undatus) and its expression under abiotic stress. Gene 2015, 563, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Jiang, Q.; Wang, F.; Wang, G.L.; Xu, Z.S.; Xiong, A.S. Selection of suitable reference genes for qPCR normalization under abiotic stresses and hormone stimuli in carrot leaves. PLoS ONE 2015, 10, e117569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaakola, L.; Pirttila, A.M.; Halonen, M.; Hohtola, A. Isolation of high quality RNA from bilberry (Vaccinium myrtillus L.) fruit. Mol. Biotechnol. 2001, 19, 201–203. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Li, M.Y.; Wang, F.; Jiang, Q.; Wang, G.L.; Tian, C.; Xiong, A.S. Validation and comparison of reference genes for qPCR normalization of celery (Apium graveolens) at different development stages. Front. Plant Sci. 2016, 7, 313. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Qi, J.; Chen, M.; Zhang, G.; Tao, A.; Fang, P.; Xu, J.; Onyedinma, S.A.; Su, J. Reference genes selection for transcript normalization in kenaf (Hibiscus cannabinus L.) under salinity and drought stress. PeerJ 2015, 3, e1347. [Google Scholar] [CrossRef] [Green Version]
- VanGuilder, H.D.; Vrana, K.E.; Freeman, W.M. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques 2008, 44, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Qi, J.; Zhang, G.; Xu, J.; Tao, A.; Fang, P.; Su, J. Selection of reliable reference genes for quantitative real-time PCR gene expression analysis in Jute (Corchorus capsularis) under stress treatments. Front. Plant Sci. 2015, 6, 848. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.M.; Zhao, J.; Scandalios, J.G. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J. 2000, 22, 87–95. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.Y.; Wang, P.C.; Chen, J.; Song, C.P. Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Bogdanovic, M.D.; Dragicevic, M.B.; Tanic, N.T.; Todorovic, S.I.; Misic, D.M.; Zivkovic, S.T.; Tissier, A.; Simonovic, A.D. Reverse transcription of 18S rRNA with poly (dT) (18) and other homopolymers. Plant Mol. Biol. Rep. 2013, 31, 55–63. [Google Scholar] [CrossRef]
- Zhu, P.; Ma, Y.; Zhu, L.; Chen, Y.; Li, R.; Ji, K. Selection of suitable reference genes in Pinus massoniana Lamb. under different abiotic stresses for qPCR normalization. Forests 2019, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Gao, Z.; Li, H.; Li, Q.; Zhang, C.; Xu, W.; Song, S.; Ma, C.; Wang, S. Selection of reference genes for miRNA qRT-PCR under abiotic stress in grapevine. Sci. Rep. 2018, 8, 4444. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, J.; Li, G.; Qi, L.; Yang, F.; Wang, R.; Li, G. Reference gene selection for qRT-PCR in Caragana korshinskii Kom. under different stress conditions. Mol. Biol. Rep. 2014, 41, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Chen, M.; Huang, X.; Chen, H.; Tao, A.; Xu, J.; Qi, J. Reference gene selection for qRT-PCR normalization analysis in kenaf (Hibiscus cannabinus L.) under abiotic stress and hormonal stimuli. Front. Plant Sci. 2017, 8, 771. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Blumwald, E. Assessing reference genes for accurate transcript normalization using quantitative real-time PCR in pearl millet [Pennisetum glaucum (L.) R. Br.]. PLoS ONE 2014, 9, e0106308. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, B.; Tan, Z.; Liu, J.; Yang, Z.; Li, Z.; Huang, B. Selection of reference genes for quantitative real-time PCR normalization in creeping bentgrass involved in four abiotic stresses. Plant Cell Rep. 2015, 34, 1825–1834. [Google Scholar] [CrossRef]
- Wang, H.L.; Chen, J.; Tian, Q.; Wang, S.; Xia, X.; Yin, W. Identification and validation of reference genes for Populus euphratica gene expression analysis during abiotic stresses by quantitative real-time PCR. Physiol. Plant 2014, 152, 529–545. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, J.; Liu, Y.; Chen, J.; Lin, H.; Huang, Y.; Bian, X.; Zhao, Y. Selection and validation of appropriate reference genes for real-time quantitative PCR analysis in Momordica charantia. Phytochemistry 2019, 164, 1–11. [Google Scholar] [CrossRef]
- Barsalobres-Cavallari, C.F.; Severino, F.E.; Maluf, M.P.; Maia, I.G. Identification of suitable internal control genes for expression studies in Coffea arabica under different experimental conditions. BMC Mol. Biol. 2009, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhakar, R.P.; Srinivas, R.D.; Sivasakthi, K.; Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Evaluation of sorghum [Sorghum bicolor (L.)] reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Luo, J.; Xu, S.; Wang, W.; Liu, T.; Han, C.; Chen, Y.; Kong, L. Selection of reference genes for gene expression normalization in Peucedanum praeruptorum Dunn under abiotic stresses, hormone treatments and different tissues. PLoS ONE 2016, 11, e152356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of reference genes for quantitative gene expression studies in Platycladus orientalis (Cupressaceae) using real-time PCR. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, B.; Liu, B.; Bi, Y.; Hou, W.; Wu, C.; Han, T. Validation of internal control for gene expression study in soybean by quantitative real-time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Lu, H.; He, Z.; Wang, C.; Wang, Y.; Ji, X. Selection of appropriate reference genes for quantitative real-time reverse transcription PCR in Betula platyphylla under salt and osmotic stress conditions. PLoS ONE 2019, 14, e225926. [Google Scholar] [CrossRef]
- Huang, L.; Yan, H.; Jiang, X.; Zhang, X.; Zhang, Y.; Huang, X.; Zhang, Y.; Miao, J.; Xu, B.; Frazier, T.; et al. Evaluation of candidate reference genes for normalization of quantitative RT-PCR in switchgrass under various abiotic stress conditions. BioEnergy Res. 2014, 7, 1201–1211. [Google Scholar] [CrossRef]
- Gimenez, M.J.; Piston, F.; Atienza, S.G. Identification of suitable reference genes for normalization of qPCR data in comparative transcriptomics analyses in the Triticeae. Planta 2011, 233, 163–173. [Google Scholar] [CrossRef]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Ciaffi, M. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol. 2009, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, K.; de Campos, M.K.F.; Pereira, L.F.P.; Vieira, L.G.E. Reference gene selection for real-time quantitative polymerase chain reaction normalization in “Swingle” citrumelo under drought stress. Anal. Biochem. 2010, 402, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Long, X.Y.; Wang, J.R.; Ouellet, T.; Rocheleau, H.; Wei, Y.M.; Pu, Z.E.; Jiang, Q.T.; Lan, X.J.; Zheng, Y.L. Genome-wide identification and evaluation of novel internal control genes for Q-PCR based transcript normalization in wheat. Plant Mol. Biol. 2010, 74, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadzillah, N.M.; Gill, V.; Finch, R.P.; Burdon, R.H. Chilling, oxidative stress and antioxidant responses in shoot cultures of rice. Planta 1996, 199, 552–556. [Google Scholar] [CrossRef]
- Purev, M.; Kim, Y.; Kim, M.K.; Pulla, R.K.; Yang, D. Isolation of a novel catalase (Cat1) gene from Panax ginseng and analysis of the response of this gene to various stresses. Plant Physiol. Biochem. 2010, 48, 451–460. [Google Scholar] [CrossRef]
- Xing, Y.; Jia, W.; Zhang, J. AtMEK1 mediates stress-induced gene expression of CAT1 catalase by triggering H2O2 production in Arabidopsis. J. Exp. Bot. 2007, 58, 2969–2981. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Description | Arabidopsis Homolog Locus | E-Value | Identities |
|---|---|---|---|---|---|
| GAPDH | MN905721 | Glyceraldehyde 3-phosphate dehydrogenase | AT1G13440 | 4 ×10−81 | 80% |
| 18S | MN905722 | 18S ribosomal RNA | AT2G47420 | 0.005 | 88% |
| ACT2 | MN905723 | Actin 2 | AT3G18780 | 4 ×10-141 | 80% |
| TIP41 | MN905724 | TIP41-like protein | AT4G34270 | 4×10-15 | 84% |
| EF-1a | MN905725 | Elongation factor 1-alpha | AT5G60390 | 4×10-101 | 81% |
| TUB | MN905726 | Tubulin beta-6 | AT5G12250 | 0 | 82% |
| eIF-4α | MN905727 | Eukaryotic translation initiation factor 4α-1 | AT3G13920 | 0 | 81% |
| PP2A-1 | MN905728 | Protein phosphatase 2A-1 | AT1G59830 | 2×10-31 | 82% |
| UBC9 | MN905729 | Ubiquitin-conjugating enzyme 9 | AT4G2796 | 6×10-60 | 81% |
| UBQ10 | MN905730 | polyubiquitin 10 | AT4G05320 | 10-164 | 80% |
| PTBP1 | MN905731 | Polypyrimidine tractbinding protein | AT3G01150 | 7×10-18 | 80% |
| TUA | MN905732 | Tubulin alpha-2 | AT1G50010 | 0 | 99% |
| ADP | MN905733 | ADP-ribosylation factor | AT1G02430 | 10-102 | 84% |
| HIS | MN905734 | Histone | AT5G10980 | 2×10-52 | 81% |
| UBQ7 | MN905735 | Ubiquitin-like protein RUB2 | AT2G35635 | 3×10-33 | 79% |
| ACT12 | MN905736 | Actin protein coding 12 | AT3G46520 | 10-124 | 80% |
| Target gene | |||||
| CAT | MN905737 | catalase | AT4G21120 | 0.14 | 85% |
| Gene Symbol | Primer Sequence (5′–3′) Forward/Reverse | Amplicon Length (bp) | R2 | PCR Efficiency (%) | SD | CV |
|---|---|---|---|---|---|---|
| GAPDH | ATTGGAAGACTCGTCGCTCG/ACCGAAAACAGCCACAGGTT | 201 | 0.9949 | 104.13 | 1.10 | 5.11 |
| 18S | CAGCGCCATCAAGGAGGAAT/ACCATGCGAGGATCCAACC | 209 | 0.9997 | 103.43 | 0.92 | 3.18 |
| ACT2 | TGAGCTACGAGTTGCTCCAG/GGCGACATACATTGCAGGTG | 130 | 0.9982 | 100.53 | 0.64 | 2.56 |
| TIP41 | ATGCCCGTCAAGAATGGGAG/TCAACGGGTGGTAAGGCTTC | 166 | 0.9922 | 99.79 | 0.83 | 3.06 |
| EF-1a | TGTGTTGGACTGCCACACTT/TGGGTTTGGAGGGCATCATC | 152 | 0.9972 | 103.43 | 1.40 | 6.12 |
| TUB | TGGTACCATGGATAGCGTGC/TGCCCCTTAGCCCAATTGTT | 105 | 0.9924 | 100.07 | 1.23 | 4.36 |
| eIF-4α | GCTCTTTGCAAGCTATGATG/CACATCAAGACCCTTGCAGA | 151 | 0.9958 | 99.93 | 1.40 | 6.12 |
| PP2A-1 | GGAGACATCCATGGGCAGTT/ACGACACGGTCTCAACAGG | 130 | 0.9933 | 99.98 | 0.66 | 2.47 |
| UBC9 | TCCCTATGCAGGGGGTGTAT/GGATCCGTCAACAAGGAGCA | 210 | 0.9918 | 98.35 | 1.15 | 4.95 |
| UBQ10 | GATGGACGTACTCTCGCTGA/AAAATCGCCACCACGAAGAC | 81 | 0.9992 | 99.29 | 1.04 | 4.55 |
| PTBP1 | CCCGTCGAAGGTTTTGCATC/AGCCTGGTTATGGTTGGCTC | 130 | 0.9994 | 101.12 | 0.71 | 2.78 |
| TUA | ATAAGACAGTTGGCGGTGGG/TGCTCTGGGTGAAAGAGCTG | 157 | 0.9992 | 105.62 | 1.23 | 4.98 |
| ADP | ACCAAGCTCTTTCAGCGTCT/GGTCGTCTTACCAGCAGCAT | 81 | 0.9957 | 108.72 | 1.33 | 5.94 |
| HIS | CGAGGCTTACCTTGTAGGGC/CCCTTTCACCGCGAATCCTT | 116 | 0.9978 | 99.5 | 1.06 | 4.77 |
| UBQ7 | CTCCTGTGCAACAGAGGCTT/TAATGACCACCACGTAGGGC | 124 | 0.9978 | 99.66 | 0.86 | 3.54 |
| ACT12 | CTTGCCGGTCGGGATTTAAC/TTCCAGGGAGGAACTGGTCT | 174 | 0.9963 | 96.85 | 0.89 | 3.70 |
| Target gene | ||||||
| CAT | TGCTCACCGTGCTGCATCTA/GCGGCATTGAACACCCCATT | 148 | 0.9918 | 100.38 |
| Rank | Drought | Salt | Cold | Heat | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | GAPDH | 0.25 | PP2A-1 | 0.14 | PP2A-1 | 0.20 | TIP41 | 0.48 | PP2A-1 | 0.47 |
| 2 | 18S | 0.25 | UBQ10 | 0.14 | GAPDH | 0.20 | ACT2 | 0.48 | ACT2 | 0.47 |
| 3 | PP2A-1 | 0.31 | UBC9 | 0.27 | TIP41 | 0.22 | PTBP1 | 0.51 | TIP41 | 0.53 |
| 4 | UBQ10 | 0.35 | 18S | 0.29 | PTBP1 | 0.24 | eIF-4α | 0.66 | PTBP1 | 0.57 |
| 5 | UBC9 | 0.39 | TIP41 | 0.31 | 18S | 0.26 | 18S | 0.73 | 18S | 0.61 |
| 6 | TIP41 | 0.41 | eIF-4α | 0.34 | ACT2 | 0.28 | HIS | 0.79 | eIF-4α | 0.63 |
| 7 | EF-1α | 0.42 | EF-1α | 0.37 | UBQ10 | 0.29 | UBC9 | 0.86 | UBC9 | 0.67 |
| 8 | eIF-4α | 0.45 | TUB | 0.40 | ACT12 | 0.33 | UBQ10 | 0.92 | UBQ10 | 0.71 |
| 9 | ACT12 | 0.48 | PTBP1 | 0.42 | eIF-4α | 0.36 | TUB | 0.96 | GAPDH | 0.73 |
| 10 | ACT2 | 0.50 | ACT2 | 0.45 | UBC9 | 0.40 | PP2A-1 | 1.03 | HIS | 0.75 |
| 11 | PTBP1 | 0.53 | GAPDH | 0.47 | EF-1α | 0.45 | UBQ7 | 1.07 | ACT12 | 0.77 |
| 12 | TUB | 0.56 | UBQ7 | 0.50 | UBQ7 | 0.51 | GAPDH | 1.12 | UBQ7 | 0.80 |
| 13 | TUA | 0.61 | ACT12 | 0.53 | HIS | 0.55 | TUA | 1.17 | TUA | 0.84 |
| 14 | HIS | 0.67 | TUA | 0.59 | TUA | 0.66 | ACT12 | 1.22 | EF-1α | 0.88 |
| 15 | UBQ7 | 0.72 | HIS | 0.66 | TUB | 0.75 | EF-1α | 1.28 | TUB | 0.93 |
| 16 | ADP | 1.11 | ADP | 1.06 | ADP | 1.15 | ADP | 1.64 | ADP | 1.10 |
| Rank | Drought | Salt | Cold | Heat | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | PP2A-1 | 0.08 | ACT2 | 0.06 | GAPDH | 0.07 | 18S | 0.20 | 18S | 0.15 |
| 2 | 18S | 0.09 | 18S | 0.07 | 18S | 0.10 | PTBP1 | 0.24 | UBC9 | 0.27 |
| 3 | GAPDH | 0.09 | UBC9 | 0.07 | UBC9 | 0.12 | ACT2 | 0.41 | PTBP1 | 0.28 |
| 4 | eIF-4α | 0.13 | PP2A-1 | 0.07 | PP2A-1 | 0.12 | UBC9 | 0.41 | TIP41 | 0.38 |
| 5 | UBQ10 | 0.17 | UBQ10 | 0.14 | ACT2 | 0.18 | TUB | 0.45 | eIF-4α | 0.39 |
| 6 | UBC9 | 0.22 | eIF-4α | 0.15 | UBQ10 | 0.23 | eIF-4α | 0.48 | UBQ10 | 0.39 |
| 7 | TIP41 | 0.25 | GAPDH | 0.16 | TIP41 | 0.24 | TIP41 | 0.60 | ACT2 | 0.39 |
| 8 | ACT2 | 0.29 | TIP41 | 0.20 | eIF-4α | 0.25 | UBQ10 | 0.61 | PP2A-1 | 0.42 |
| 9 | EF-1α | 0.31 | EF-1α | 0.28 | PTBP1 | 0.28 | HIS | 0.68 | GAPDH | 0.46 |
| 10 | TUA | 0.41 | TUB | 0.31 | EF-1α | 0.35 | TUA | 0.68 | HIS | 0.47 |
| 11 | ACT12 | 0.42 | PTBP1 | 0.39 | ACT12 | 0.44 | GAPDH | 0.76 | ACT12 | 0.57 |
| 12 | TUB | 0.43 | ACT12 | 0.48 | UBQ7 | 0.65 | PP2A-1 | 0.79 | TUA | 0.59 |
| 13 | PTBP1 | 0.44 | UBQ7 | 0.49 | HIS | 0.71 | UBQ7 | 0.95 | UBQ7 | 0.61 |
| 14 | UBQ7 | 0.72 | TUA | 0.55 | TUA | 0.73 | EF-1α | 1.04 | EF-1α | 0.66 |
| 15 | HIS | 0.79 | HIS | 0.87 | TUB | 0.74 | ACT12 | 1.05 | TUB | 0.71 |
| 16 | ADP | 2.68 | ADP | 2.70 | ADP | 2.75 | ADP | 2.84 | ADP | 1.50 |
| Rank | Drought | Salt | Cold | Heat | Total | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | |
| 1 | GADPH | 0.17 | 0.79 | PP2A-1 | 0.17 | 0.65 | PP2A-1 | 0.40 | 1.51 | UBQ7 | 0.29 | 1.22 | ACT2 | 0.54 | 2.17 |
| 2 | PTBP1 | 0.63 | 2.43 | UBQ10 | 0.19 | 0.84 | ACT12 | 0.42 | 1.74 | TIP41 | 0.43 | 1.60 | PP2A-1 | 0.58 | 2.18 |
| 3 | HIS | 0.66 | 2.90 | PTBP1 | 0.23 | 0.93 | PTBP1 | 0.44 | 1.72 | ACT2 | 0.60 | 2.42 | PTBP1 | 0.67 | 2.60 |
| 4 | ACT2 | 0.69 | 2.74 | TIP41 | 0.23 | 0.88 | ACT2 | 0.46 | 1.84 | PP2A-1 | 0.66 | 2.52 | TIP41 | 0.77 | 2.84 |
| 5 | ACT12 | 0.85 | 3.43 | ACT12 | 0.30 | 1.24 | TIP41 | 0.47 | 1.73 | PTBP1 | 0.81 | 3.14 | UBQ7 | 0.79 | 3.23 |
| 6 | TUB | 0.87 | 3.12 | UBC9 | 0.35 | 1.54 | 18S | 0.53 | 1.88 | GAPDH | 0.93 | 4.51 | ACT12 | 0.85 | 3.54 |
| 7 | PP2A-1 | 0.88 | 3.26 | 18S | 0.36 | 1.29 | eIF-4α | 0.54 | 2.23 | eIF-4α | 0.99 | 4.13 | GAPDH | 0.86 | 4.02 |
| 8 | UBQ10 | 0.90 | 3.91 | UBQ7 | 0.36 | 1.52 | UBQ10 | 0.56 | 2.49 | ACT12 | 1.05 | 4.43 | 18S | 0.90 | 3.11 |
| 9 | 18S | 1.09 | 3.74 | EF-1α | 0.42 | 1.90 | GAPDH | 0.60 | 2.89 | HIS | 1.06 | 4.73 | eIF-4α | 0.90 | 3.70 |
| 10 | UBQ7 | 1.13 | 4.46 | eIF-4α | 0.42 | 1.77 | UBQ7 | 0.76 | 3.11 | TUA | 1.14 | 4.65 | HIS | 1.02 | 4.59 |
| 11 | EF-1α | 1.15 | 4.96 | TUB | 0.52 | 1.93 | HIS | 0.76 | 3.40 | TUB | 1.19 | 4.14 | UBQ10 | 1.02 | 4.48 |
| 12 | eIF-4α | 1.16 | 4.60 | ACT2 | 0.53 | 2.14 | UBC9 | 0.94 | 4.09 | 18S | 1.32 | 4.59 | UBC9 | 1.15 | 4.94 |
| 13 | TIP41 | 1.18 | 4.24 | HIS | 0.57 | 2.67 | EF-1α | 0.95 | 4.24 | UBC9 | 1.55 | 6.67 | TUB | 1.20 | 4.25 |
| 14 | UBC9 | 1.20 | 5.06 | GAPDH | 0.60 | 2.84 | TUA | 1.07 | 4.48 | UBQ10 | 1.73 | 7.48 | TUA | 1.22 | 4.93 |
| 15 | TUA | 1.39 | 5.56 | TUA | 0.87 | 3.67 | TUB | 1.65 | 5.86 | EF-1α | 2.15 | 9.18 | ADP | 1.26 | 5.62 |
| 16 | ADP | 3.28 | 15.30 | ADP | 2.87 | 14.06 | ADP | 3.10 | 14.77 | ADP | 3.25 | 15.22 | EF-1α | 1.39 | 6.07 |
| Rank | Drought | Salt | Cold | Heat | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | GAPDH | 1.44 | PP2A-1 | 2.00 | PP2A-1 | 1.59 | TIP41 | 2.41 | ACT2 | 2.41 |
| 2 | PP2A-1 | 2.76 | UBQ10 | 2.15 | GAPDH | 2.62 | ACT2 | 2.62 | PP2A-1 | 2.52 |
| 3 | 18S | 3.30 | UBC9 | 3.78 | 18S | 3.91 | PTBP1 | 3.11 | PTBP1 | 3.30 |
| 4 | UBQ10 | 5.43 | 18S | 3.83 | TIP41 | 4.72 | 18S | 3.91 | 18S | 3.42 |
| 5 | PTBP1 | 6.59 | ACT2 | 4.93 | PTBP1 | 4.76 | UBQ7 | 5.23 | TIP41 | 3.63 |
| 6 | ACT2 | 6.84 | TIP41 | 5.43 | ACT2 | 4.93 | eIF-4α | 5.52 | UBC9 | 5.52 |
| 7 | eIF-4α | 7.27 | PTBP1 | 6.67 | ACT12 | 5.60 | UBC9 | 7.14 | eIF-4α | 6.46 |
| 8 | UBC9 | 7.49 | eIF-4α | 7.11 | UBQ10 | 6.95 | PP2A-1 | 7.83 | UBQ10 | 8.08 |
| 9 | ACT12 | 7.91 | EF-1α | 8.28 | UBC9 | 7.11 | HIS | 7.86 | GAPDH | 8.28 |
| 10 | TIP41 | 8.17 | ACT12 | 9.21 | eIF-4α | 7.96 | TUB | 7.91 | ACT12 | 8.99 |
| 11 | HIS | 8.57 | TUB | 9.58 | EF-1α | 11.27 | GAPDH | 9.25 | UBQ7 | 9.21 |
| 12 | EF-1α | 8.85 | GAPDH | 10.25 | UBQ7 | 11.29 | UBQ10 | 9.64 | HIS | 10.00 |
| 13 | TUB | 9.52 | UBQ7 | 10.77 | HIS | 12.30 | TUA | 10.91 | TUA | 12.97 |
| 14 | TUA | 12.49 | HIS | 14.30 | TUA | 14.00 | ACT12 | 11.89 | TUB | 14.30 |
| 15 | UBQ7 | 12.81 | TUA | 14.33 | TUB | 15.00 | EF-1α | 14.66 | EF-1α | 14.64 |
| 16 | ADP | 16.00 | ADP | 16.00 | ADP | 16.00 | ADP | 16.00 | ADP | 15.66 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Yu, S.; Zeng, M.; Liu, X.; Yang, J.; Li, C. Selection and Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis in Needles of Larix olgensis under Abiotic Stresses. Forests 2020, 11, 193. https://doi.org/10.3390/f11020193
Li D, Yu S, Zeng M, Liu X, Yang J, Li C. Selection and Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis in Needles of Larix olgensis under Abiotic Stresses. Forests. 2020; 11(2):193. https://doi.org/10.3390/f11020193
Chicago/Turabian StyleLi, Dandan, Sen Yu, Minzhen Zeng, Xiao Liu, Jia Yang, and Chenghao Li. 2020. "Selection and Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis in Needles of Larix olgensis under Abiotic Stresses" Forests 11, no. 2: 193. https://doi.org/10.3390/f11020193
APA StyleLi, D., Yu, S., Zeng, M., Liu, X., Yang, J., & Li, C. (2020). Selection and Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis in Needles of Larix olgensis under Abiotic Stresses. Forests, 11(2), 193. https://doi.org/10.3390/f11020193