Responses of Parameters for Electrical Impedance Spectroscopy and Pressure–Volume Curves to Drought Stress in Pinus bungeana Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Drought Treatments
2.2. Electrical Impedance Spectroscopy
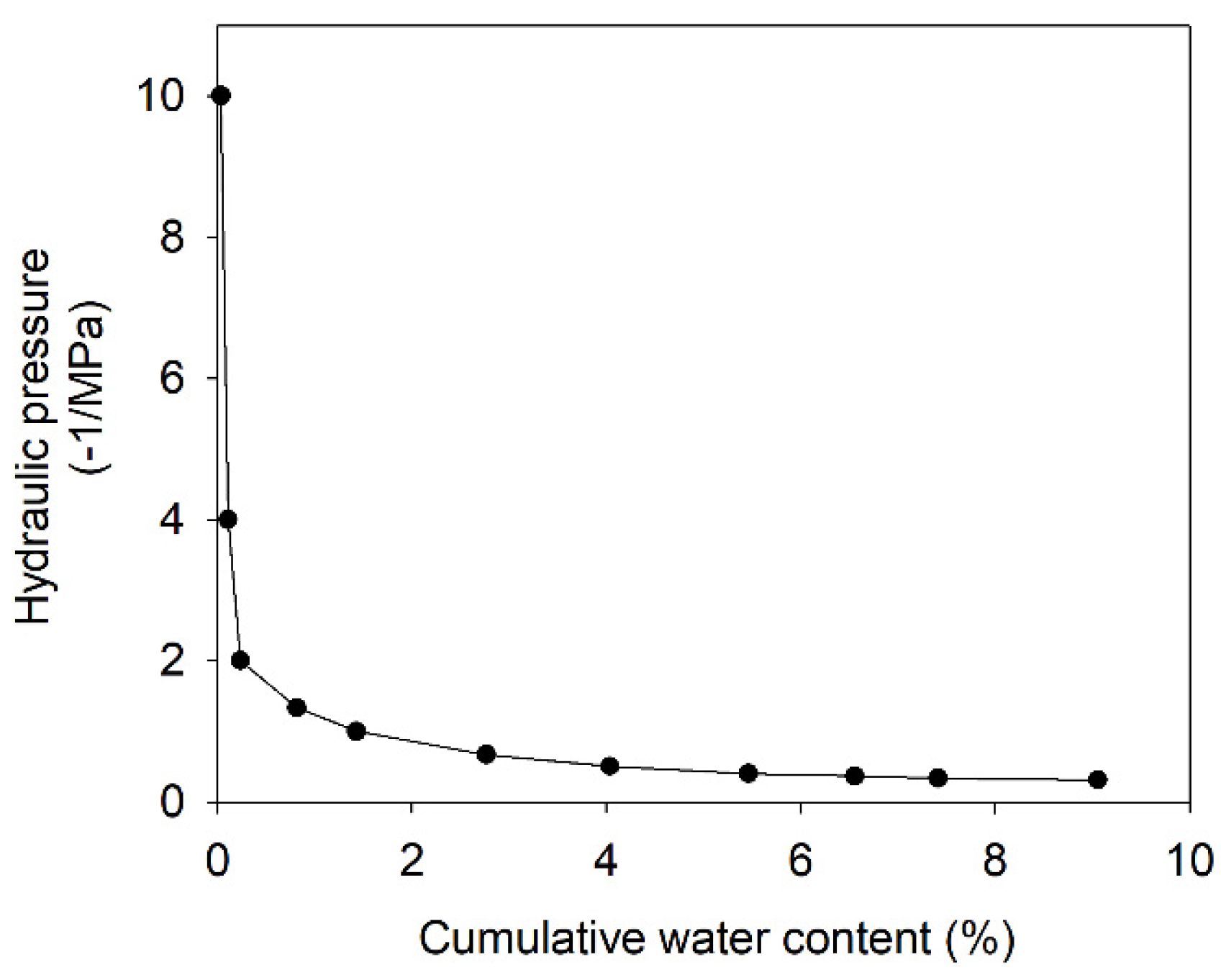
2.3. Pressure–Volume (PV) Curves and Their Parameters
2.4. Statistical Analyses
3. Results
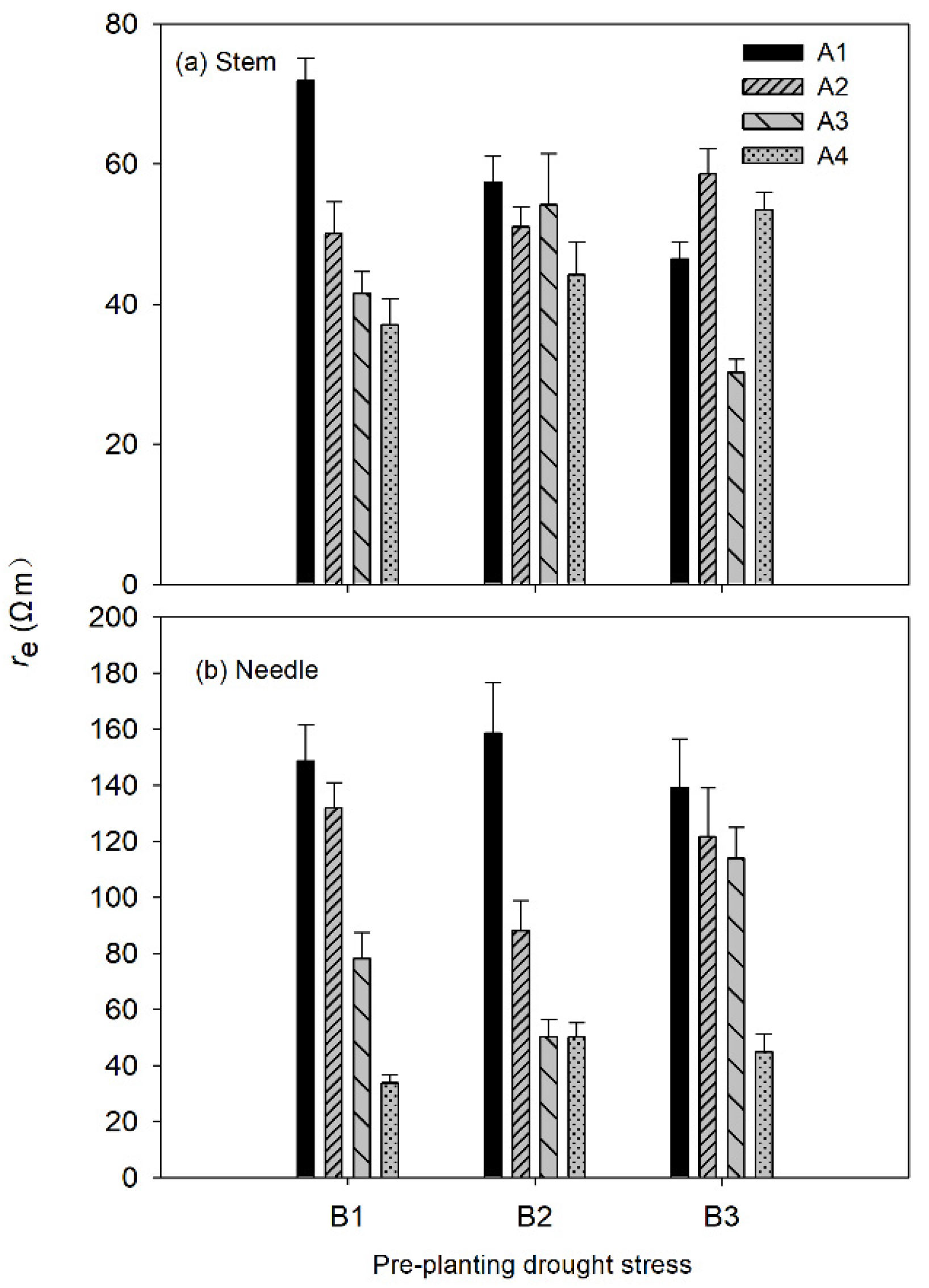
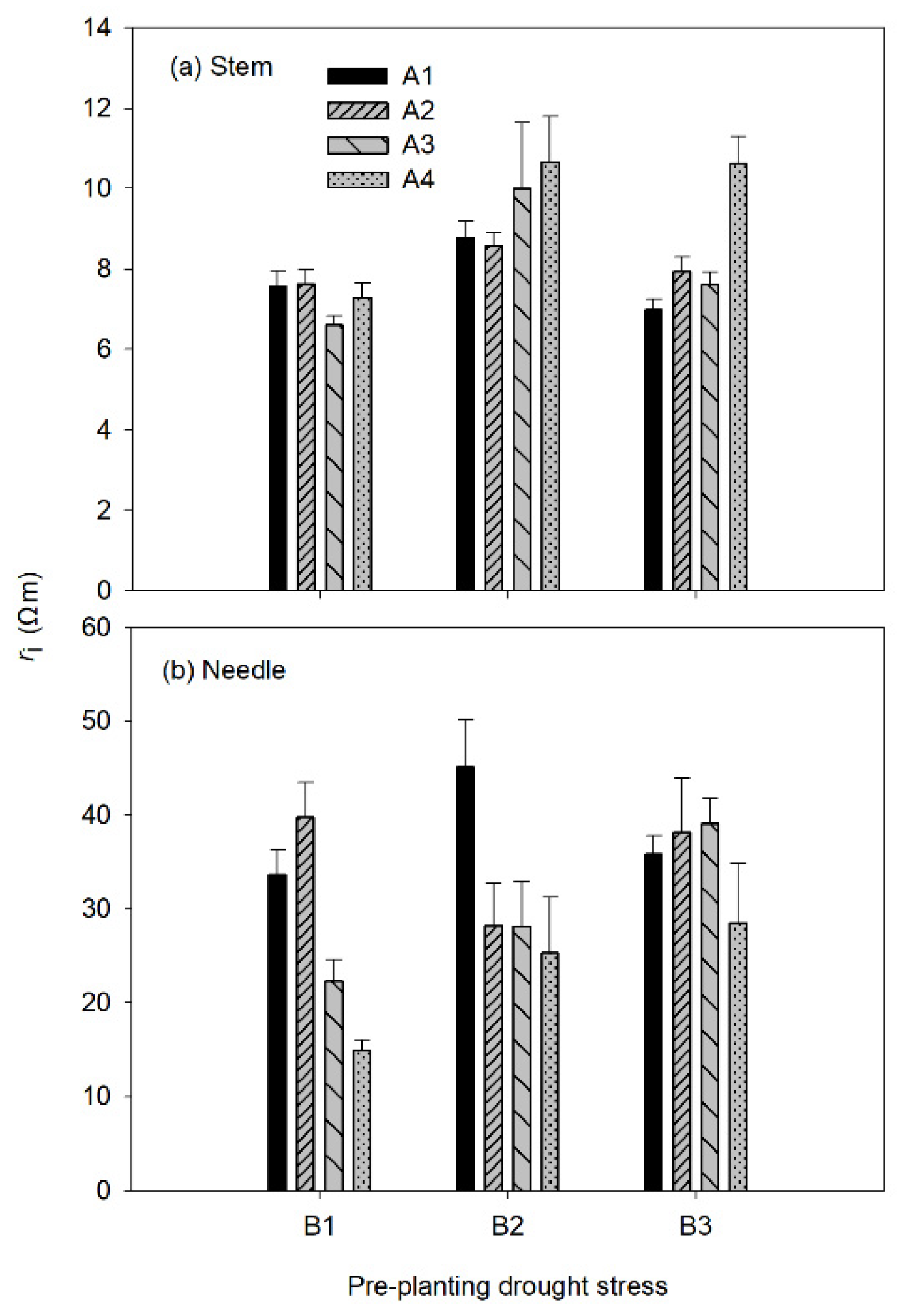
3.1. Electrical Impedance Spectroscopy (EIS) Parameters
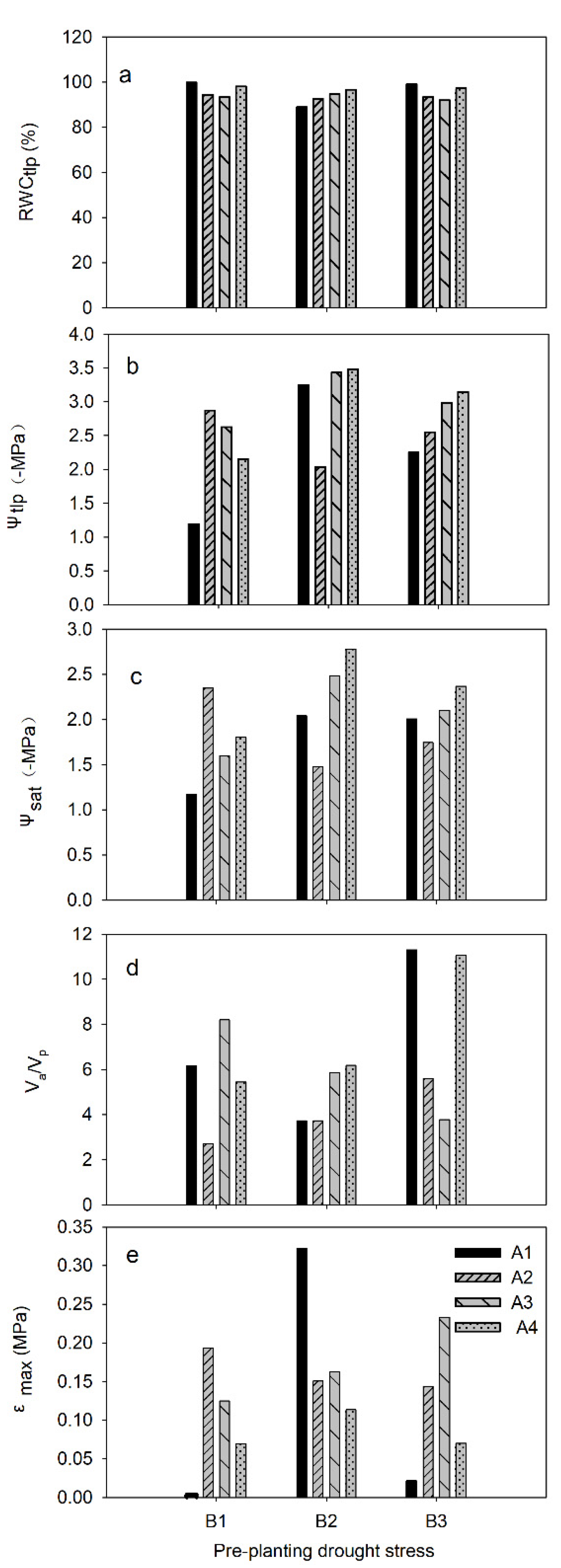
3.2. Hydraulic Parameters of Shoots by PV Curves After Four Weeks Post-Planting Growth
3.3. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, L.; Xiao, J.; Zhou, Y.; Zheng, Y.; Li, J.; Xiao, H. Drought events and their effects on vegetation productivity in China. Ecosphere 2016, 7, e01591. [Google Scholar] [CrossRef]
- Dickman, L.T.; McDowell, N.G.; Sevanto, S.; Pangle, R.E.; Pockman, W.T. Carbohydrate dynamics and mortality in a piñon-juniper woodland under three future precipitation scenarios. Plant Cell Environ. 2015, 38, 729–739. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Harayama, H.; Ikeda, T.; Ishida, A.; Yamamoto, S.I. Seasonal variations in water relations in current-year leaves of evergreen trees with delayed greening. Tree Physiol. 2006, 26, 1025–1033. [Google Scholar] [CrossRef] [Green Version]
- Zolfaghar, S.; Villalobos-Vega, R.; Cleverly, J.; Eamus, D. Co-ordination among leaf water relations and xylem vulnerability to embolism of Eucalyptus trees growing along a depth-to-groundwater gradient. Tree Physiol. 2015, 35, 732–743. [Google Scholar] [CrossRef] [Green Version]
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in vascular plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Bowman, W.D.; Roberts, S.W. Seasonal changes in tissue elasticity in chaparral shrubs. Plant Physiol. 1985, 65, 233–236. [Google Scholar] [CrossRef]
- Pallardy, S.G. Transpiration and Plant Water Balance. In Physiology of Woody Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 325–366. [Google Scholar]
- Tyree, M.T.; Hammel, H.T. The measurement of the turgor pressure and water relations of plants by the pressure-bomb technique. J. Exp. Bot. 1972, 23, 267–282. [Google Scholar] [CrossRef]
- Rascio, A.; Platani, C.; Di Fonzo, N.; Wittmer, G. Bound water in durum wheat under drought stress. Plant Physiol. 1992, 98, 908–912. [Google Scholar] [CrossRef] [Green Version]
- Arabzadeh, N.; Emadian, S.F. Effect of water (drought) stress on water relations of Haloxylon Aphyllum and H. Persicum. Iran. J. Sci. Technol. 2010, 34, 245–255. [Google Scholar]
- Bartlett, M.K.; Zhang, Y.; Kreidler, N.; Sun, S.; Ardy, R.C.; Cao, K.; Sack, L. Global analysis of plasticity in turgor loss point, a key drought tolerance trait. Ecol. Lett. 2014, 17, 1580–1590. [Google Scholar] [CrossRef]
- Banks, J.M.; Percival, G.C.; Rose, G. Variations in seasonal drought tolerance rankings. Trees 2019, 33, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.C.; Jones, M.M. Turgor Maintenance by Osmotic Adjustment: AReview and Evaluation. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; John Wiley and Sons: New York, NY, USA, 1980; pp. 87–103. [Google Scholar]
- Di, X.Y.; Wang, M.B.; Chen, J.W.; Zhang, W.F. Study on water parameters with PV curves in eight poplar clones. Acta Bot. Boreali Occident. Sin. 2007, 27, 98–103, (In Chinese with English abstract). [Google Scholar]
- Martínez, J.P.; Silva, H.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- MacDonald, J.R. Emphasizing Solid Materials and Systems. In Impedance Spectroscopy; John Wiley & Sons Inc.: New York, NY, USA, 1987. [Google Scholar]
- Grimnes, S.; Martinsen, O.G. Bioimpedance and Bioelectricity Basics; Academic Press: San Diego, CA, USA, 2000; p. 359. [Google Scholar]
- Repo, T.; Oksanen, E.; Vapaavuori, E. Effects of elevated concentrations of ozone and carbon dioxide on the electrical impedance of leaves of silver birch (Betula pendula) clones. Tree Physiol. 2004, 24, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Repo, T.; Zhang, M.I.N.; Ryyppö, A.; Vapaavuori, E.; Sutinen, S. Effects of freeze-thaw injury on parameters of distributed electrical circuits of stems and needles of Scots pine seedlings at different stages of acclimation. J. Exp. Bot. 1994, 45, 823–833. [Google Scholar] [CrossRef]
- Zhang, M.I.N.; Willison, J.H.M.; Cox, M.A. Measurement of heat injury in plant tissue by using electrical impedance analysis. Can. J. Bot. 1993, 71, 1605–1611. [Google Scholar] [CrossRef]
- Repo, T.; Zhang, G.; Ryyppö, A.; Rikala, R.; Vuorinen, M. The relation between growth cessation and frost hardening in Scots pines of different origins. Trees 2000, 14, 456–464. [Google Scholar] [CrossRef]
- Repo, T.; Zhang, G.; Ryyppö, A.; Rikala, R. The electrical impedance spectroscopy of Scots pine (Pinus sylvestris L.) shoots in relation to cold acclimation. J. Exp. Bot. 2000, 51, 2095–2107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Ryyppö, A.; Repo, T. The electrical impedance spectroscopy of Scots pine needles during cold acclimation. Physiol. Plant. 2002, 115, 385–392. [Google Scholar] [CrossRef]
- Mancuso, S.; Rinaldelli, E. Response of young mycorrhizal and non-mycorrhizal plants of olive tree (Olea europaea L) to saline conditions. II. Dynamics of electrical impedance parameters of shoots and leaves. Adv. Hortic. Sci. 1996, 10, 135–145. [Google Scholar]
- Wang, A.F.; Roitto, M.; Lehto, T.; Zwiazek, J.J.; Calvo-Polanco, M.; Repo, T. Waterlogging under simulated late-winter conditions had little impact on the physiology and growth of Norway spruce seedlings. Ann. For. Sci. 2013, 70, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Ozier-Lafontaine, H.; Bajazet, T. Analysis of root growth by impedance spectroscopy (EIS). Plant Soil 2005, 277, 299–313. [Google Scholar] [CrossRef]
- Čermák, J.; Ulrich, R.; Staněk, Z.; Koller, J.; Aubrecht, L. Electrical measurement of tree root absorbing surfaces by the earth impedance method: 2. Verification based on allometric relationships and root severing experiments. Tree Physiol. 2006, 26, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Ellis, T.W.; Murray, W.; Paul, K.; Kavalieris, L.; Brophy, J.; Williams, C.; Maass, M. Electrical capacitance as a rapid and non-invasive indicator of root length. Tree Physiol. 2012, 33, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Repo, T.; Korhonen, A.; Laukkanen, M.; Lehto, T.; Silvennoinen, R. Detecting mycorrhizal colonisation in Scots pine roots using electrical impedance spectra. Biosyst. Eng. 2014, 121, 139–149. [Google Scholar] [CrossRef]
- Repo, T.; Korhonen, A.; Lehto, T.; Silvennoinen, R. Assessment of frost damage in mycorrhizal and non-mycorrhizal roots of Scots pine seedlings using classification analysis of their electrical impedance spectra. Trees 2016, 30, 483–495. [Google Scholar] [CrossRef]
- Cseresnyés, I.; Takács, T.; Sepovics, B.; Kovács, R.; Füzy, A.; Parádi, I.; Rajkai, K. Electrical characterization of the root system: A noninvasive approach to study plant stress responses. Acta Physiol. Plant. 2019, 41, 169. [Google Scholar] [CrossRef] [Green Version]
- Repo, T.; Paine, D.H.; Taylor, A.G. Electrical impedance spectroscopy in relation to seed viability and moisture content in snap bean (Phaseolus vulgaris L.). Seed Sci. Res. 2002, 12, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Tiitta, M.; Repo, T.; Viitanen, H. Effect of soft rot and bacteria on electrical impedance of wood at low moisture content. Mater. Org. 1999, 33, 261–287. [Google Scholar]
- Colombo, S.J.; Blumwald, E. Electrical impedance of white spruce shoots in relation to pressure-volume analysis and free sugar content. Plant Cell Environ. 1992, 15, 837–842. [Google Scholar] [CrossRef]
- Repo, T. Influence of different electrodes and tissues on the impedance spectra of Scots pine shoots. Electro Magn. 1994, 13, 1–14. [Google Scholar] [CrossRef]
- Zhang, M.I.N.; Repo, T.; Willison, J.H.M.; Sutinen, S. Electrical impedance analysis in plant tissues: On the biological meaning of Cole-Cole α in Scots pine needles. Eur. Biophys. J. 1995, 24, 99–106. [Google Scholar] [CrossRef]
- Sun, Z.H.; Wang, Q.C. The drought resistance of four broad-leaved species in the north of China with PV technique. Sci. Silv. Sin. 2003, 39, 33–38, (In Chinese with English abstract). [Google Scholar]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Inoue, Y.; Kenzo, T.; Tamai, S.; Yamamoto, F.; Yamanaka, N.; Ichie, T. Drought tolerance of three fabaceous seedlings grown under different soil water conditions in semi-arid land of South America. J. Jpn. Soc. Reveg. Technol. 2018, 43, 499–508. [Google Scholar] [CrossRef]
- Wang, A.F.; Zhang, G. Effects of drought on electrical impedance spectroscopy parameters in stems of Pinus bungeana Zucc. seedlings. Front. Agric. China 2010, 4, 468–474. [Google Scholar] [CrossRef]
- Wang, A.F.; Zhang, G.; Qie, Y.X.; Xiang, D.Y.; Di, B.; Chen, Z.P. Effects of drought on physiological characteristics of needles of Pinus bungeana Zucc. seedlings. Plant Physiol. J. 2012, 48, 189–196, (In Chinese with English abstract). [Google Scholar]
- Wang, A.F.; Zhang, G.; Wei, S.C.; Cui, T.X. Relation between frost hardiness and parameters of electrical impedance spectroscopy in saplings of different development stage of Pinus sylvestris L. var. mongolica Litv. Acta Ecol. Sin. 2008, 28, 5741–5749, (In Chinese with English abstract). [Google Scholar]
- Ryyppö, A.; Repo, T.; Vapaavuori, E. Development of freezing tolerance in roots and shoots of scots pine seedlings at nonfreezing temperatures. Can. J. For. Res. 1998, 28, 557–565. [Google Scholar] [CrossRef]
- Repo, T.; Hiekkala, P.; Hietala, T.; Tahvanainen, L. Intracellular resistance correlates with initial stage of frost hardening in willow (Salix viminalis). Physiol. Plant. 1997, 101, 627–634. [Google Scholar] [CrossRef]
- Kandiko, R.A.; Timmins, R.; Worrall, J. Pressure-volume curves of shoots and roots of normal and drought conditioned western hemlock seedlings. Can. J. For. Res. 1980, 10, 10–16. [Google Scholar] [CrossRef]
- Pita, P.; Pardos, J.A. Growth, leaf morphology, water use and tissue water relations of Eucalyptus globulus clones in response to water deficit. Tree Physiol. 2001, 21, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Patakas, A.; Noitsakis, B. Cell wall elasticity as a mechanism to maintain favorable water relations during leaf ontogeny in grapevines. Am. J. Enol. Vitic. 1997, 48, 352–356. [Google Scholar]
- Patakas, A.; Nikolaou, N.; Zioziou, E.; Radoglou, K.; Noitsakis, B. The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines. Plant Sci. 2002, 163, 361–367. [Google Scholar] [CrossRef]
- Dichio, B.; Xiloyannis, C.; Angelopoulos, K.; Nuzzo, V.; Bufo, A.B.; Celano, G. Drought-induced variations of water relations parameters in Olea europea. Plant Soil 2003, 257, 381–389. [Google Scholar] [CrossRef]
- Tyree, M.T.; Karamanos, A.J. Water Stress as an Ecological Factor. In Plants and Their Atmospheric Environment; Grace, J.B., Ford, E.D., Jarvis, P.G., Eds.; Blackwell Scientific Publications: Oxford, UK, 1981; pp. 237–261. [Google Scholar]
- Niinemets, U. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Lenz, T.I.; Wright, I.J.; Westoby, M. Interrelations among pressure–volume curve traits across species and water availability gradients. Physiol. Plant 2006, 127, 423–433. [Google Scholar] [CrossRef]
- Xu, H.L.; Xu, Q.C.; Qin, F.F.; Tian, C.M.; Wang, R. Applications of xeropytophysiology in plant production—Tomato fruit yield and quality improved by restricted irrigations in soil-based greenhouses. Acta Hortic. 2011, 893. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Data | p Values | ||||||
|---|---|---|---|---|---|---|---|---|
| t | A | B | A × B | t × A | t × B | t × A × B | ||
| re | Stem | 0.000 | 0.000 | 0.000 | 0.009 | 0.000 | 0.000 | 0.000 |
| ri | Stem | 0.000 | 0.661 | 0.436 | 0.002 | 0.000 | 0.000 | 0.000 |
| τ1 | Stem | 0.310 | 0.640 | 0.660 | 0.012 | 0.000 | 0.020 | 0.000 |
| τ2 | Stem | 0.000 | 0.340 | 0.008 | 0.835 | 0.000 | 0.011 | 0.000 |
| ψ1 | Stem | 0.494 | 0.110 | 0.946 | 0.109 | 0.264 | 0.400 | 0.059 |
| ψ2 | Stem | 0.000 | 0.110 | 0.111 | 0.003 | 0.004 | 0.004 | 0.000 |
| re | Needle | 0.000 | 0.000 | 0.227 | 0.001 | 0.000 | 0.136 | 0.000 |
| ri | Needle | 0.000 | 0.000 | 0.080 | 0.010 | 0.000 | 0.000 | 0.000 |
| τm | Needle | 0.000 | 0.000 | 0.003 | 0.058 | 0.000 | 0.005 | 0.000 |
| β | Needle | 0.000 | 0.000 | 0.005 | 0.251 | 0.000 | 0.025 | 0.000 |
| Stem | Needle | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| re | ri | τ1 | τ2 | ψ1 | ψ2 | re | ri | τm | β | |
| RWCtlp | 0.24 | −0.03 | −0.03 | 0.18 | 0.09 | −0.18 | 0.24 | −0.43 | −0.18 | −0.51 * |
| ψtlp | 0.46 | −0.56 * | −0.24 | −0.30 | 0.12 | −0.14 | 0.29 | −0.32 | 0.24 | −0.02 |
| ψsat | 0.43 | −0.58 * | −0.09 | −0.17 | 0.07 | −0.11 | 0.36 | −0.19 | 0.28 | 0.18 |
| Va/Vp | 0.05 | 0.08 | 0.41 | 0.12 | −0.07 | −0.03 | −0.16 | −0.07 | −0.09 | −0.38 |
| εmax | −0.37 | 0.40 | 0.32 | 0.34 | −0.1 | 0.10 | −0.18 | 0.35 | −0.17 | 0.19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, A.-F.; Di, B.; Repo, T.; Roitto, M.; Zhang, G. Responses of Parameters for Electrical Impedance Spectroscopy and Pressure–Volume Curves to Drought Stress in Pinus bungeana Seedlings. Forests 2020, 11, 359. https://doi.org/10.3390/f11030359
Wang A-F, Di B, Repo T, Roitto M, Zhang G. Responses of Parameters for Electrical Impedance Spectroscopy and Pressure–Volume Curves to Drought Stress in Pinus bungeana Seedlings. Forests. 2020; 11(3):359. https://doi.org/10.3390/f11030359
Chicago/Turabian StyleWang, Ai-Fang, Bao Di, Tapani Repo, Marja Roitto, and Gang Zhang. 2020. "Responses of Parameters for Electrical Impedance Spectroscopy and Pressure–Volume Curves to Drought Stress in Pinus bungeana Seedlings" Forests 11, no. 3: 359. https://doi.org/10.3390/f11030359
APA StyleWang, A. -F., Di, B., Repo, T., Roitto, M., & Zhang, G. (2020). Responses of Parameters for Electrical Impedance Spectroscopy and Pressure–Volume Curves to Drought Stress in Pinus bungeana Seedlings. Forests, 11(3), 359. https://doi.org/10.3390/f11030359