Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
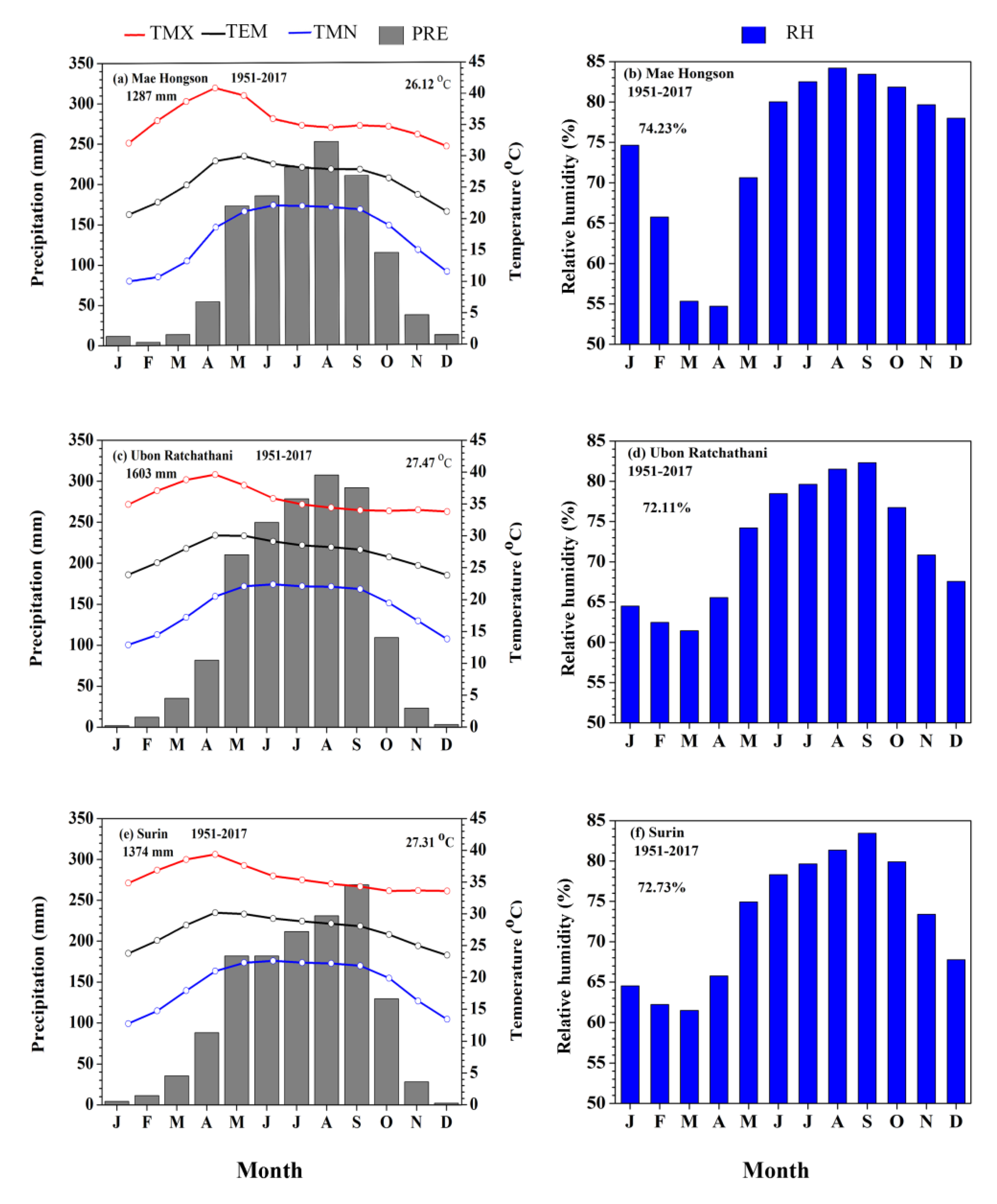
2.1. Study Area and Climate
2.2. Tree-Ring Sampling and Chronology Development
2.3. Climate-Growth Responses
2.4. Long-Term Growth Trends
3. Results
3.1. Tree-Ring Chronology Statistics
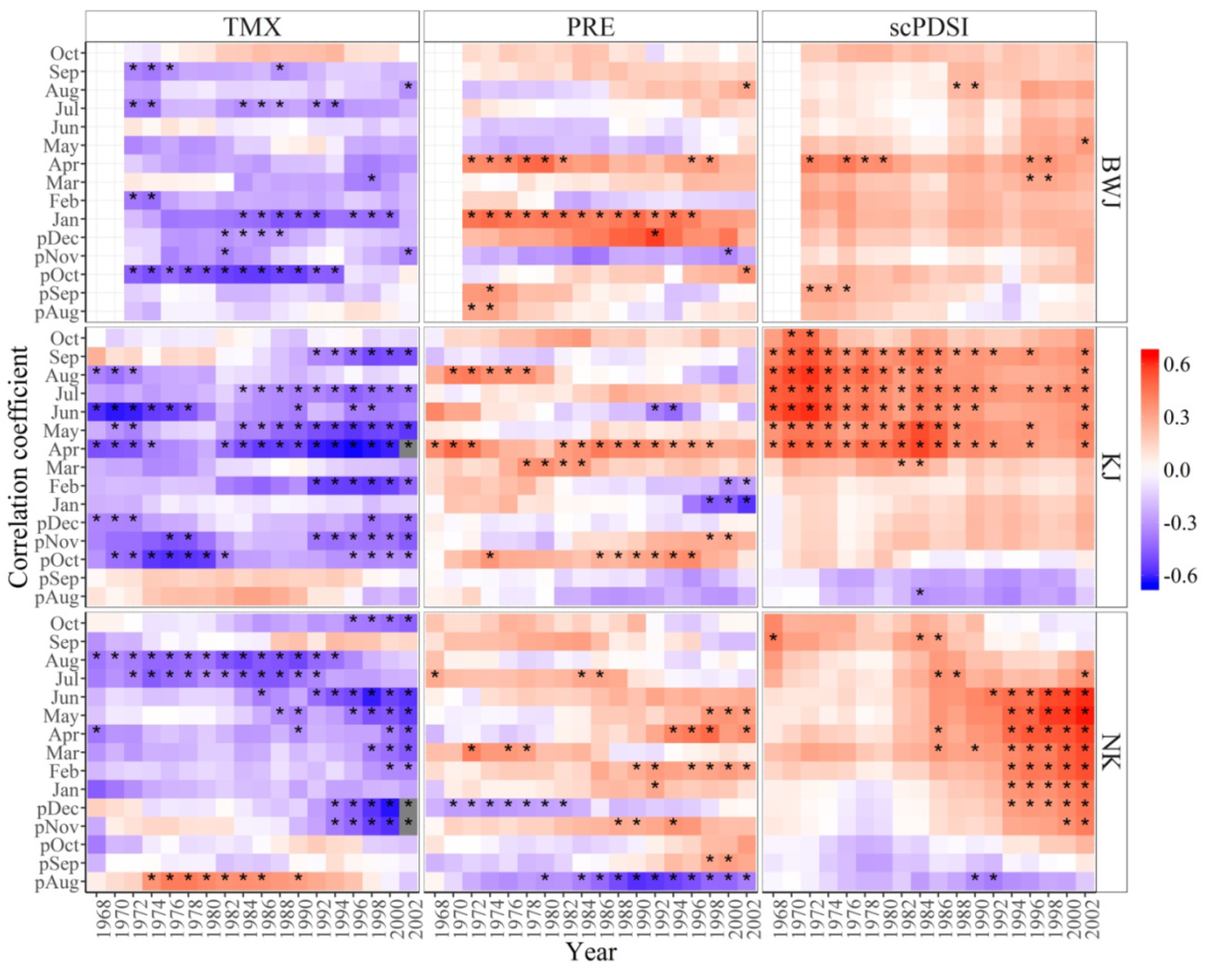
3.2. Growth-Climate Relationships
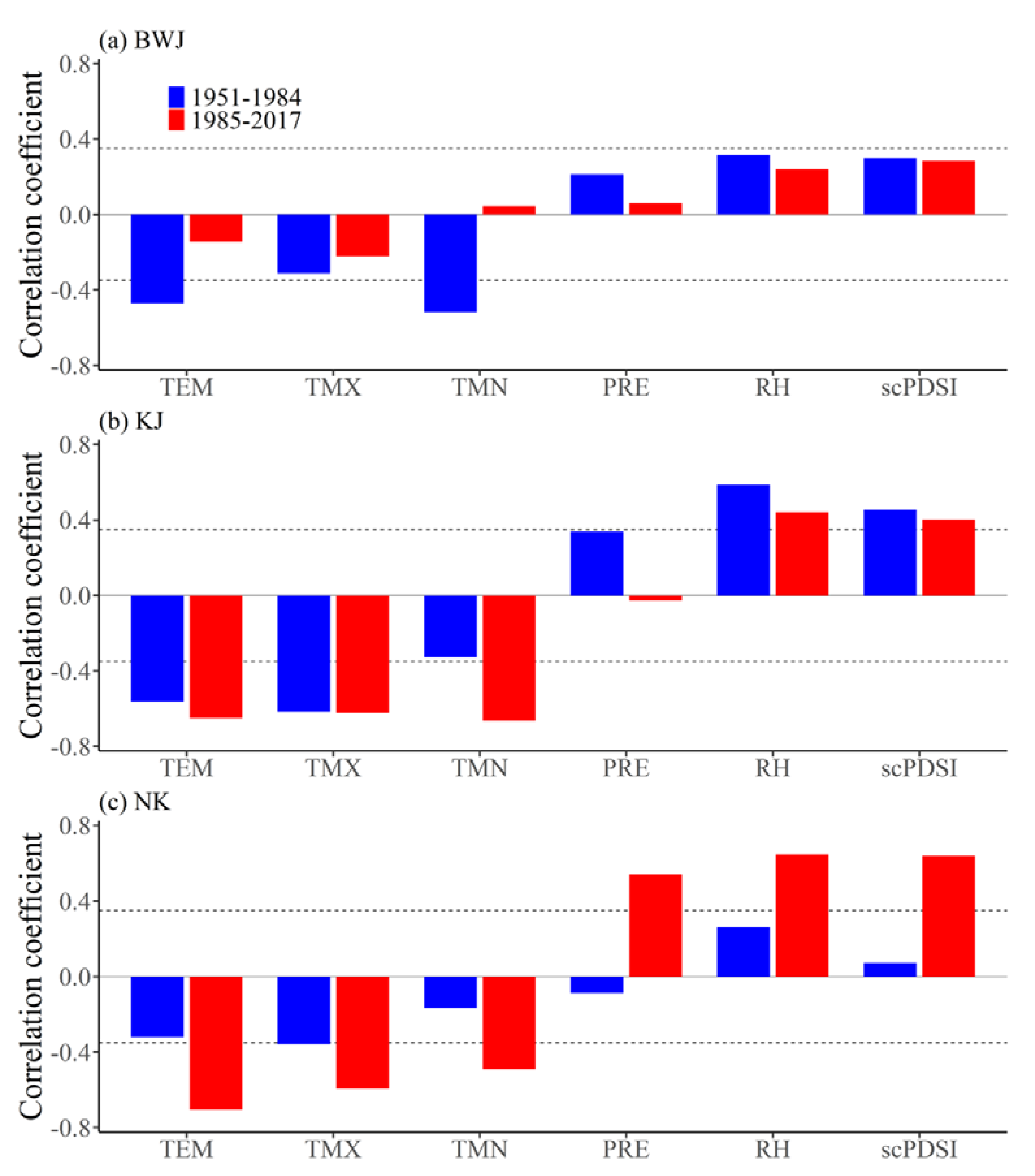
3.3. Temporal Stability of the Climate-Growth Relationships
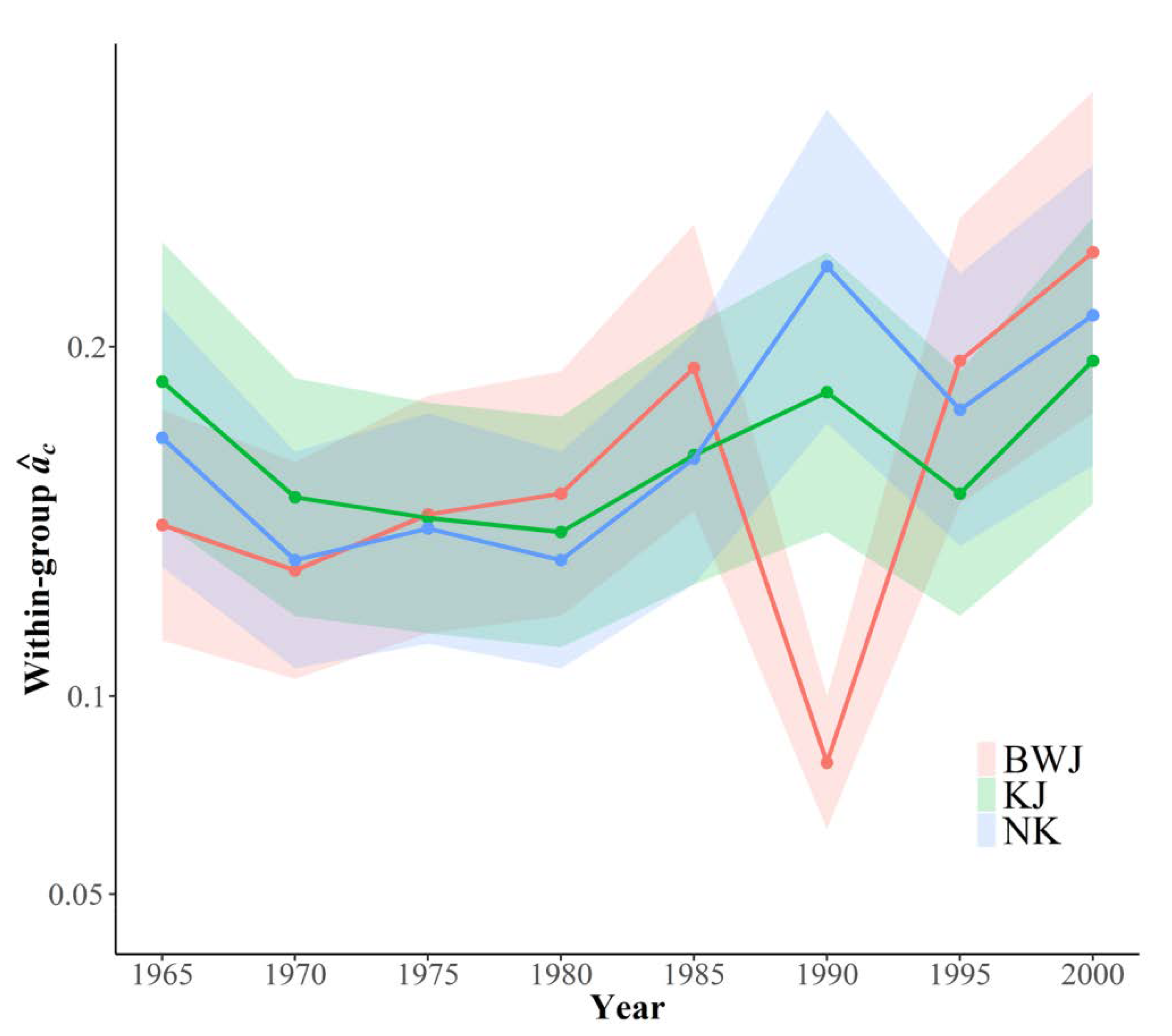
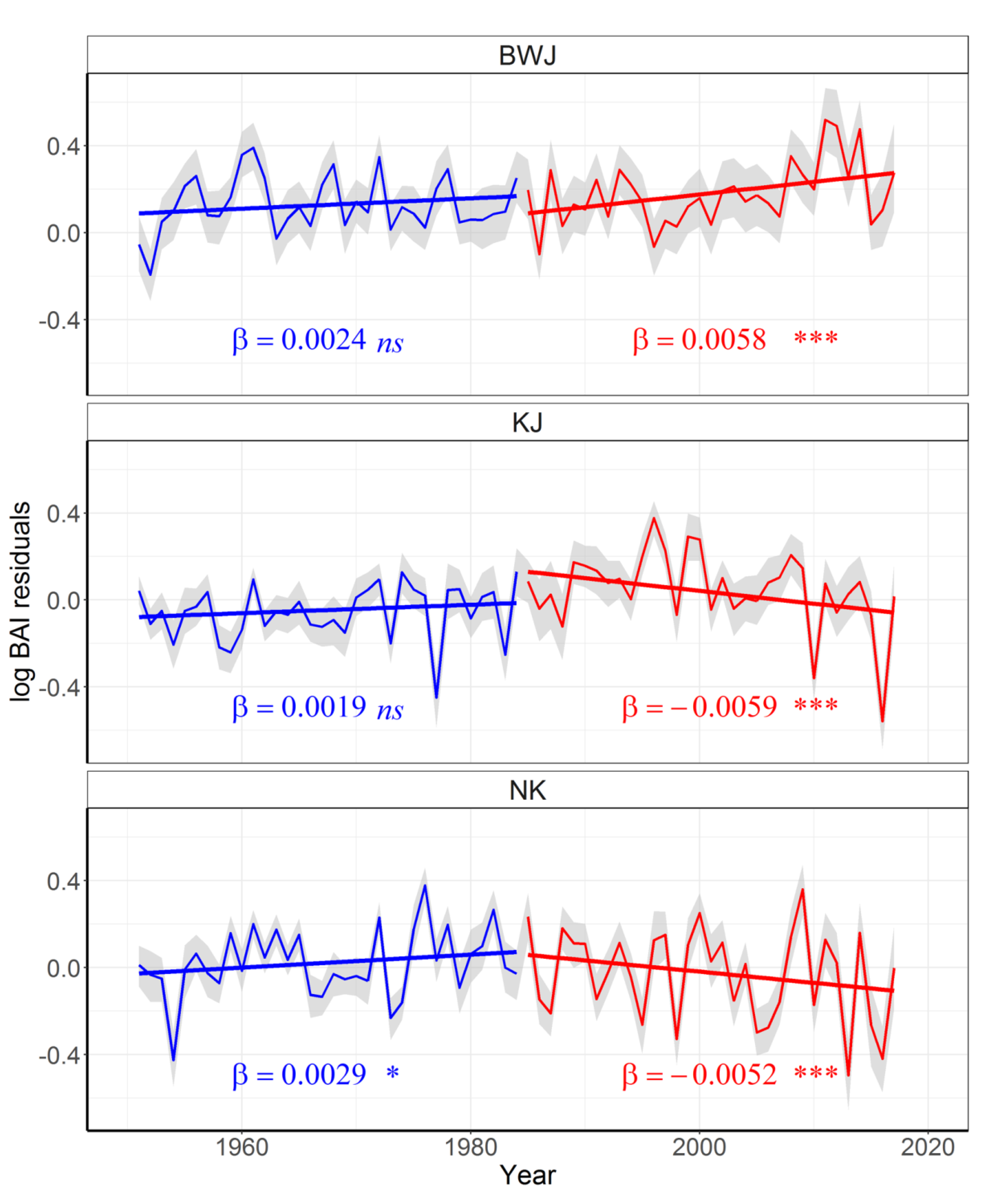
3.4. Growth Synchrony and Long-Term Trends
4. Discussion
4.1. Dominant Climate-Growth Relationships
4.2. Temporal Shift of Growth-Climate Sensitivity
4.3. Growth Synchrony and Long-Term Trends
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clark, D.B.; Clark, D.A.; Oberbauer, S.F. Annual wood production in a tropical rain forest in NE Costa Rica linked to climatic variation but not to increasing CO2. Glob. Chang. Biol. 2010, 16, 747–759. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Corlett, R.T. Impacts of warming on tropical lowland rainforests. Trends Ecol. Evol. 2011, 26, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.C.; McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.J. Larger trees suffer most during drought in forests worldwide. Nat. Plants 2015, 1, 15139. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.; Julio Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A. Forest growth responses to drought at short- and long-term scales in Spain: Squeezing the stress memory from tree rings. Front. Ecol. Evol. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Vlam, M.; Baker, P.J.; Bunyavejchewin, S.; Zuidema, P.A. Temperature and rainfall strongly drive temporal growth variation in Asian tropical forest trees. Oecologia 2014, 174, 1449–1461. [Google Scholar] [CrossRef]
- Groenendijk, P.; van der Sleen, P.; Vlam, M.; Bunyavejchewin, S.; Bongers, F.; Zuidema, P.A. No evidence for consistent long-term growth stimulation of 13 tropical tree species: Results from tree-ring analysis. Glob. Chang. Biol. 2015, 21, 3762–3776. [Google Scholar] [CrossRef]
- Zuidema, P.A.; Baker, P.J.; Groenendijk, P.; Schippers, P.; van der Sleen, P.; Vlam, M.; Sterck, F. Tropical forests and global change: Filling knowledge gaps. Trends Plant Sci. 2013, 18, 413–419. [Google Scholar] [CrossRef]
- Pumijumnong, N. Dendrochronology in Southeast Asia. Trees 2013, 27, 343–358. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Park, W.-K. Teak vessel chronologies as an indicator of Southeast Asia premonsoon temperature. Palaeobot 2001, 50, 21–26. [Google Scholar]
- Buckley, B.M.; Barbetti, M.; Watanasak, M.; D’Arrigo, R.; Boonchirdchoo, S.; Sarutanon, S. Dendrochronological Investigations in Thailand. IAWA J. 1995, 16, 393–409. [Google Scholar] [CrossRef] [Green Version]
- D’Arrigo, R.; Palmer, J.; Ummenhofer, C.C.; Kyaw, N.N.; Krusic, P. Three centuries of Myanmar monsoon climate variability inferred from teak tree rings. Geophys. Res. Lett. 2011, 38, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Buckley, B.M.; Duangsathaporn, K.; Palakit, K.; Butler, S.; Syhapanya, V.; Xaybouangeun, N. Analyses of growth rings of Pinus merkusii from Lao P.D.R. For. Ecol. Manag. 2007, 253, 120–127. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Watanasak, M.; Boonchirdchoo, S.; Krusic, P.; Sarutanon, S.; Barbett, M.; Buckley, B. Progress in Dendroclimatic Studies of Mountain Pine in Northern Thailand. IAWA J. 1997, 18, 433–444. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Wanyaphet, T. Seasonal cambial activity and tree-ring formation of Pinus merkusii and Pinus kesiya in Northern Thailand in dependence on climate. For. Ecol. Manag. 2006, 226, 279–289. [Google Scholar] [CrossRef]
- Shah, S.K.; Mehrotra, N. Tree–ring studies of Toona ciliata from subtropical wet hill forests of Kalimpong, eastern Himalaya. Dendrochronologia 2017, 46, 46–55. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, M.; Wernicke, J.; Bräuning, A. Changes in sensitivity of tree-ring widths to climate in a tropical moist forest tree in Bangladesh. Forests 2018, 9, 761. [Google Scholar] [CrossRef] [Green Version]
- Lumyai, P.; Duangsathaporn, K. Climate reconstruction on the growth of teak in Umphang wildlife sanctuary, Thailand. Environ. Nat. Res. J. 2018, 16, 21–30. [Google Scholar]
- Sano, M.; Buckley, B.M.; Sweda, T. Tree-ring based hydroclimate reconstruction over northern Vietnam from Fokienia hodginsi: Eighteenth century mega-drought and tropical Pacific influence. Clim. Dyn. 2009, 33, 331–340. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Eckstein, D. Reconstruction of pre-monsoon weather conditions in northwestern Thailand from the tree-ring widths of Pinus merkusii and Pinus kesiya. Trees 2011, 25, 125–132. [Google Scholar] [CrossRef]
- Zimmer, H.; Baker, P. Climate and historical stand dynamics in the tropical pine forests of northern Thailand. For. Ecol. Manag. 2009, 257, 190–198. [Google Scholar] [CrossRef]
- Briffa, K.R.; Schweingruber, F.H.; Jones, P.D.; Osborn, T.J.; Shiyatov, S.G.; Vaganov, E.A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 1998, 391, 678–682. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Long-term change in the sensitivity of tree-ring growth to climate forcing in Larix decidua. New Phytol. 2006, 170, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Babst, F.; Bouriaud, O.; Poulter, B.; Trouet, V.; Girardin, M.P.; Frank, D.C. Twentieth century redistribution in climatic drivers of global tree growth. Sci. Adv. 2019, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, J.E.; Smijanić, M.; Scharnweber, T.; Buras, A.; Cedro, A.; Cruz-Garciá, R.; Drobyshev, I.; Janecka, K.; Jansons, Ā.; Kaczka, R.; et al. Tree growth influenced by warming winter climate and summer moisture availability in northern temperate forests. Glob. Chang. Biol. 2019, 26, 2505–2518. [Google Scholar] [CrossRef]
- Wilmking, M.; Juday, G.P.; Barber, V.A.; Zald, H.S.J. Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Glob. Chang. Biol. 2004, 10, 1724–1736. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Gutiérrez, E.; Voltas, J. A roadmap to disentangling ecogeographical patterns of spatial synchrony in dendrosciences. Trees 2018, 32, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Averyanov, L.V.; Nguyen, T.H.; Sinh, K.N.; Pham, T.; Van Lamxay, V.; Bounphanmy, S.; Chantthavongsa, K. Gymnosperms of Laos. Nord. J. Bot. 2014, 32, 765–805. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; p. 414. [Google Scholar]
- Rinn, F. TASP-Win User Reference Mannual; Rinntech: Heidelberg, Germany, 2003. [Google Scholar]
- Holmes, R.L. Computer-assisted Quality Control in Tree-ring Dating and Measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Guide for Computer Program ARSTAN. In The International Tree-Ring Data Bank Program Library Version 2.0 User’s Mannual; The University of Arizona: Tucson, AZ, USA, 1996; Volume 2, pp. 75–87. [Google Scholar]
- Cook, E.R.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Osborn, T.J.; Biffa, K.R.; Jones, P.D. Adjusting variance for sample-size in tree-ring chronologies and other regional-mean timeseries. Dendrochronologia 1997, 15, 89–99. [Google Scholar]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization (Dendrochronology, Forestry, Dendroclimatology, Autogressive Process); The University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Shestakova, T.A.; Gutiérrez, E.; Kirdyanov, A.V.; Camarero, J.J.; Génova, M.; Knorre, A.A.; Linares, J.C.; de Dios, V.R.; Sánchez-Salguero, R.; Voltas, J. Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc. Natl. Acad. Sci. USA 2016, 113, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schurman, J.S.; Babst, F.; Björklund, J.; Rydval, M.; Bače, R.; Čada, V.; Janda, P.; Mikolas, M.; Saulnier, M.; Trotsiuk, V.; et al. The climatic drivers of primary Picea forest growth along the Carpathian arc are changing under rising temperatures. Glob. Chang. Biol. 2019, 25, 3136–3150. [Google Scholar] [CrossRef]
- Alday, J.G.; Shestakova, T.A.; de Dios, V.R.; Voltas, J. DendroSync: An R package to unravel synchrony patterns in tree-ring networks. Dendrochronologia 2018, 47, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Girardin, M.P.; Bouriaud, O.; Hogg, E.H.; Kurz, W.; Zimmermann, N.E.; Metsaranta, J.M.; de Jong, R.; Frank, D.C.; Esper, J.; Büntgen, U.; et al. No growth stimulation of Canada’s boreal forest under half-century of combined warming and CO2 fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, E8406–E8414. [Google Scholar] [CrossRef] [Green Version]
- Marchand, W.; Girardin, M.P.; Hartmann, H.; Gauthier, S.; Bergeron, Y. Taxonomy, together with ontogeny and growing conditions, drives needleleaf species’ sensitivity to climate in boreal North America. Glob. Chang. Biol. 2019, 25, 2793–2809. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: London, UK, 2017. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R package, version 3.1–127. 2017. Available online: https://CRAN.R-project.org/package=nlme (accessed on 23 March 2020).
- Ravi, S.; D’Odorico, P.; Over, T.M.; Zobeck, T.M. On the effect of air humidity on soil susceptibility to wind erosion: The case of air-dry soils. Geophys. Res. Lett. 2004, 31, L09501. [Google Scholar] [CrossRef]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, Drimys brasiliensis (Winteraceae). New Phytol. 2013, 199, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.L.; Liu, W.J.; Fan, Z.X.; Cao, K.F. Is fog an important water source for woody plants in an Asian tropical karst forest during the dry season? Ecohydrology 2016, 9, 964–972. [Google Scholar] [CrossRef]
- Panthi, S.; Bräuning, A.; Zhou, Z.; Fan, Z. Tree rings reveal recent intensified spring drought in the central Himalaya, Nepal. Glob. Planet. Chang. 2017, 157, 26–34. [Google Scholar] [CrossRef]
- Gaire, N.P.; Bhuju, D.R.; Koirala, M.; Shah, S.K.; Carrer, M.; Timilsena, R. Tree-ring based spring precipitation reconstruction in western Nepal Himalaya since AD 1840. Dendrochronologia 2017, 42, 21–30. [Google Scholar] [CrossRef]
- Hutameta, A.; Pumijumnong, N. Pine tree-ring response to climate and ENSO at Ban Wat Chan, Chiang Mai. Environ. Nat. Res. J. 2003, 1, 69–76. [Google Scholar]
- Peng, J.; Li, J.; Wang, T.; Huo, J.; Yang, L. Effect of altitude on climate–growth relationships of Chinese white pine (Pinus armandii) in the northern Funiu Mountain, central China. Clim. Chang. 2019, 154, 273–288. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Qian, H.; Liu, R. Inverse effects of recent warming on trees growing at the low and high altitudes of the Dabie Mountains, subtropical China. Dendrochronologia 2020, 59, 125649. [Google Scholar] [CrossRef]
- Koskela, J.; Hari, P.; Pipatwattanakul, D. Analysis of gas exchange of Merkus pine population by the optimality approach. Tree Physiol. 1999, 19, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Macias, M.; Andreu, L.; Bosch, O.; Camarero, J.J.; Gutiérrez, E. Increasing aridity is enhancing silver fir (Abies alba Mill.) water stress in its south-western distribution limit. Clim. Chang. 2006, 79, 289–313. [Google Scholar] [CrossRef]
- Ashiq, M.W.; Anand, M. Spatial and temporal variability in dendroclimatic growth response of red pine (Pinus resinosa Ait.) to climate in northern Ontario, Canada. For. Ecol. Manag. 2016, 372, 109–119. [Google Scholar] [CrossRef]
- Andreu, L.; Gutierrez, E.; Macias, M.; Ribas, M.; Bosch, O.; Camarero, J.J. Climate increases regional tree-growth variability in Iberian pine forests. Glob. Chang. Biol. 2007, 13, 804–815. [Google Scholar] [CrossRef]
- Sohar, K.; Altman, J.; Lehĕcková, E.; Doležal, J. Growth–climate relationships of Himalayan conifers along elevational and latitudinal gradients. Int. J. Climatol. 2017, 37, 2593–2605. [Google Scholar] [CrossRef]
- Schwab, N.; Kaczka, R.J.; Jenecka, K.; Böhner, J.; Chaudhary, R.P.; Scholten, T.; Schickhoff, U. Climate change-induced shift of tree growth sensitivity at a central Himalayan treeline ecotone. Forests 2018, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Gaire, N.P.; Fan, Z.-X.; Bräuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Su, J.; Gou, X.; Deng, Y.; Zhang, R.; Liu, W.; Zhang, F.; Lu, M.; Chen, Y.; Zheng, W. Tree growth response of Fokienia hodginsii to recent climate warming and drought in southwest China. Int. J. Biometeorol. 2017, 61, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Natalini, F.; Correia, A.C.; Vázquez-Piqué, J.; Alejano, R. Tree rings reflect growth adjustments and enhanced synchrony among sites in Iberian stone pine (Pinus pinea L.) under climate change. Ann. For. Sci. 2015, 72, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Black, B.A.; van der Sleen, P.; Di Lorenzo, E.; Griffin, D.; Sydeman, W.J.; Dunham, J.B.; Rykaczewski, R.R.; García-Reyes, M.; Safeeq, M.; Arismendi, I. Rising synchrony controls western North American ecosystems. Glob. Chang. Biol. 2018, 24, 2305–2314. [Google Scholar] [CrossRef]
- Zhou, F.; Fang, K.; Zhang, F.; Dong, Z.; Chen, D. Climate-driven synchronized growth of alpine trees in the southeast Tibetan Plateau. PLoS ONE 2016, 11, e0156126. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L. Consequences of widespread tree mortality triggered drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Charney, N.D.; Babst, F.; Poulter, B.; Record, S.; Trouet, V.M.; Frank, D.; Enquist, B.J.; Evans, M.E.K. Observed forest sensitivity to climate implies large changes in 21st century North American forest growth. Ecol. Lett. 2016, 19, 1119–1128. [Google Scholar] [CrossRef]
- Belyazid, S.; Giuliana, Z. Water limitation can negate the effect of higher temperatures on forest carbon sequestration. European. J. For. Res. 2019, 138, 287–297. [Google Scholar]
- Yu, D.; Liu, J.; Lewis, B.J.; Li, Z.; Wangming, Z.; Xiangmin, F.; Yawei, W.; Shengwei, J.; Limin, D. Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For. Ecol. Manag. 2013, 300, 96–105. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude (°N) | Longitude (°E) | Elevation (m) | Cores/Trees | Span | AGR (mm) | MSL | MS | AC1 | Rbar | EPS |
|---|---|---|---|---|---|---|---|---|---|---|---|
| BWJ | 19.08 | 98.33 | 1100 | 56/29 | 1646–2017 | 2.233 | 140 | 0.279 | 0.462 | 0.355 | 0.862 |
| KJ | 15.44 | 105.48 | 170 | 60/32 | 1854–2017 | 2.941 | 74 | 0.277 | 0.384 | 0.253 | 0.9 |
| NK | 14.68 | 103.76 | 165 | 64/31 | 1886–2017 | 2.71 | 63 | 0.312 | 0.385 | 0.359 | 0.95 |
| Site | Period | Estimate | Standard Error | df | t Value | p | AIC |
|---|---|---|---|---|---|---|---|
| BWJ | 1951–1984 | 0.00238 | 0.00151 | 1671 | 1.577 | 0.115 | 929 |
| 1985–2017 | 0.00579 | 0.00153 | 1438 | 3.782 | <0.001 | 759 | |
| KJ | 1951–1984 | 0.00193 | 0.00124 | 1627 | 1.558 | 0.119 | 747 |
| 1985–2017 | −0.00585 | 0.0.00163 | 1485 | −3.601 | <0.001 | 1041 | |
| NK | 1951–1984 | 0.00297 | 0.00146 | 1753 | 2.039 | 0.042 | 980 |
| 1985–2017 | −0.00516 | 0.00132 | 1652 | −3.891 | <0.001 | 1559 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakthai, S.; Fu, P.-L.; Fan, Z.-X.; Gaire, N.P.; Pumijumnong, N.; Eiadthong, W.; Tangmitcharoen, S. Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand. Forests 2020, 11, 361. https://doi.org/10.3390/f11030361
Rakthai S, Fu P-L, Fan Z-X, Gaire NP, Pumijumnong N, Eiadthong W, Tangmitcharoen S. Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand. Forests. 2020; 11(3):361. https://doi.org/10.3390/f11030361
Chicago/Turabian StyleRakthai, Sakkarin, Pei-Li Fu, Ze-Xin Fan, Narayan Prasad Gaire, Nathsuda Pumijumnong, Wichan Eiadthong, and Suwan Tangmitcharoen. 2020. "Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand" Forests 11, no. 3: 361. https://doi.org/10.3390/f11030361
APA StyleRakthai, S., Fu, P. -L., Fan, Z. -X., Gaire, N. P., Pumijumnong, N., Eiadthong, W., & Tangmitcharoen, S. (2020). Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand. Forests, 11(3), 361. https://doi.org/10.3390/f11030361