Differences in Near Isohydric and Anisohydric Behavior of Contrasting Poplar Hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under Drought-Rehydration Treatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Conditions
2.2. Experimental Design
2.3. Aboveground Plant Growth Performance
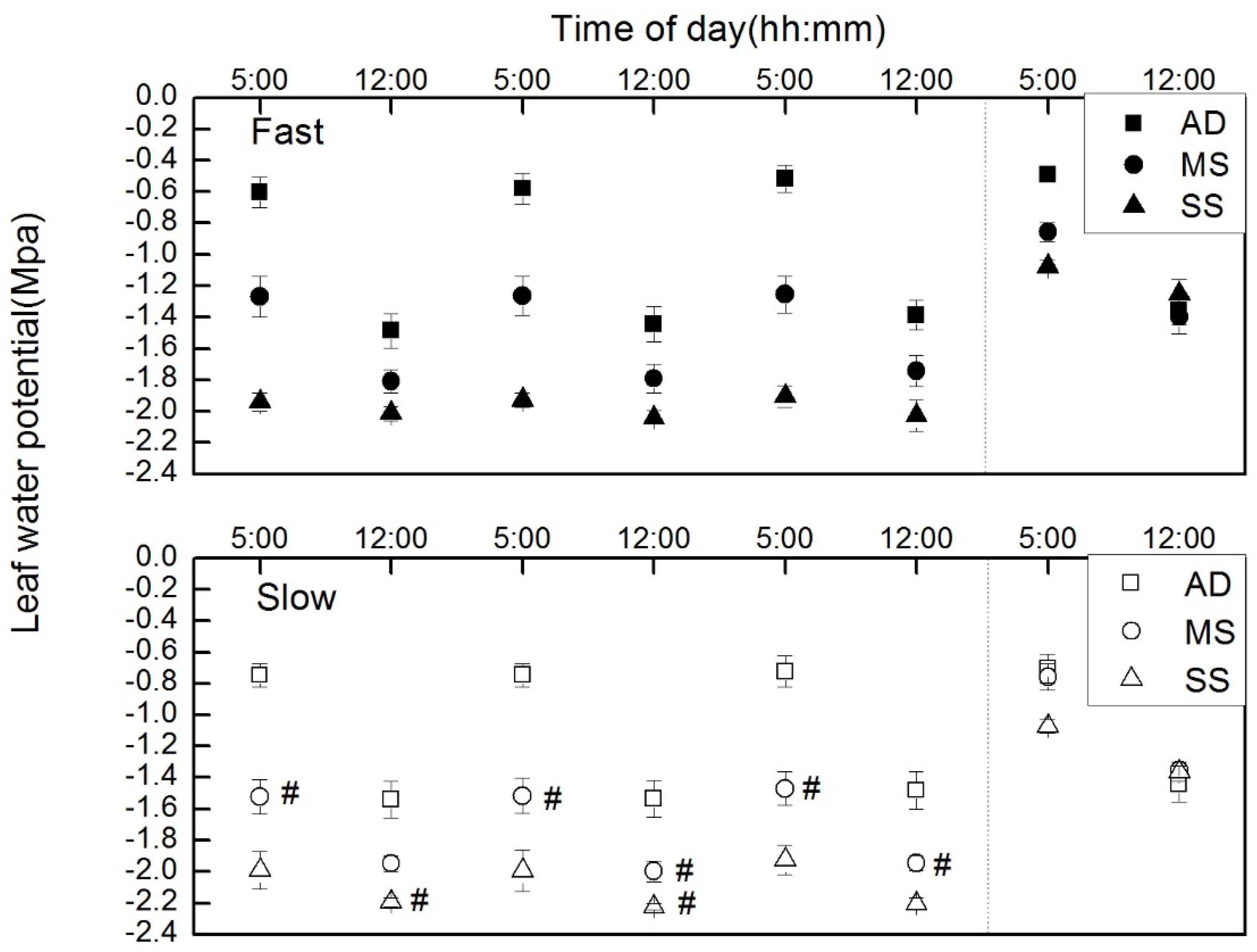
2.4. Leaf Water Potential
2.5. Native Embolism and Specific Conductivity
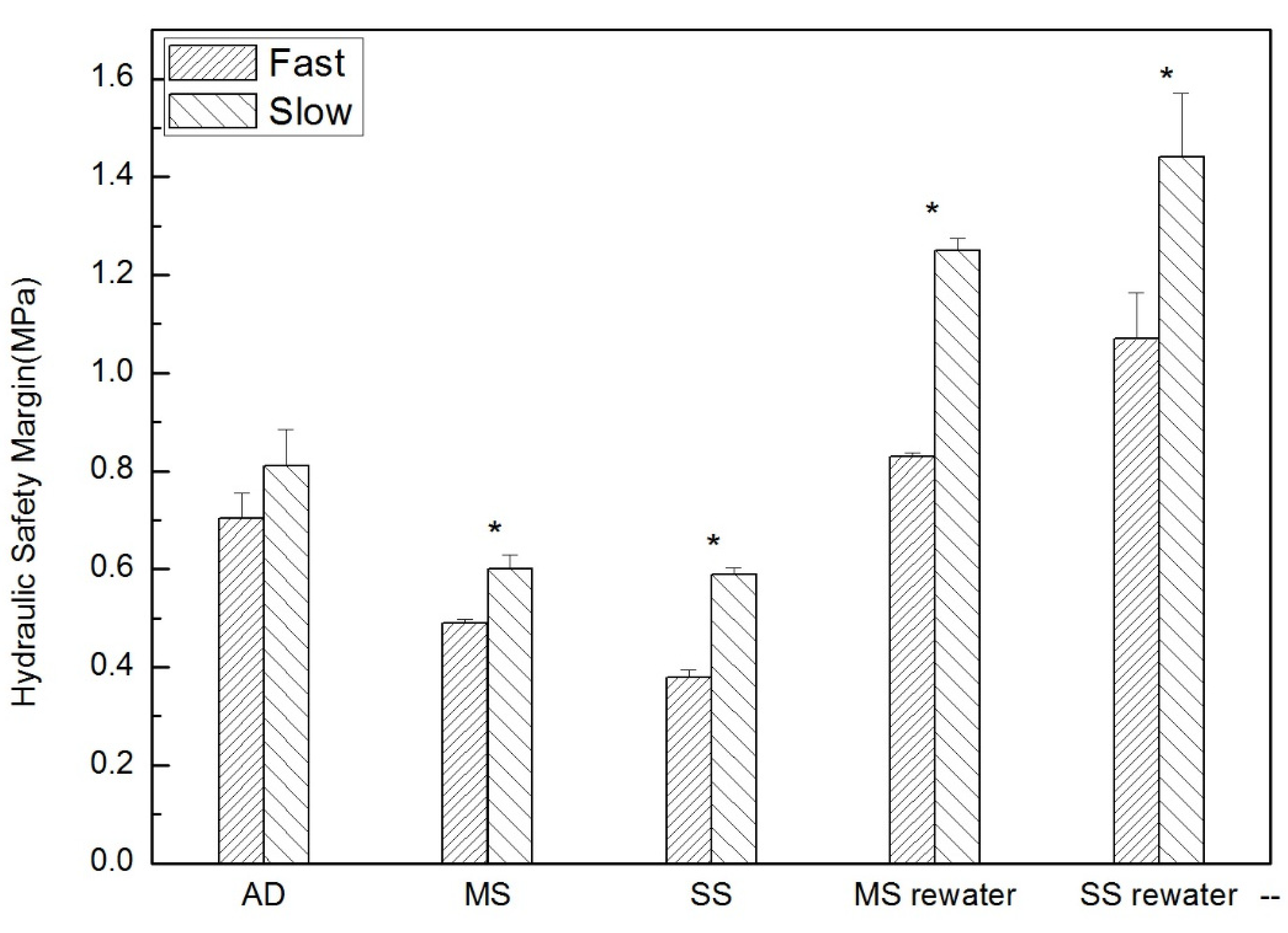
2.6. Vulnerability Curves and Estimation of Hydraulic Safety Margins
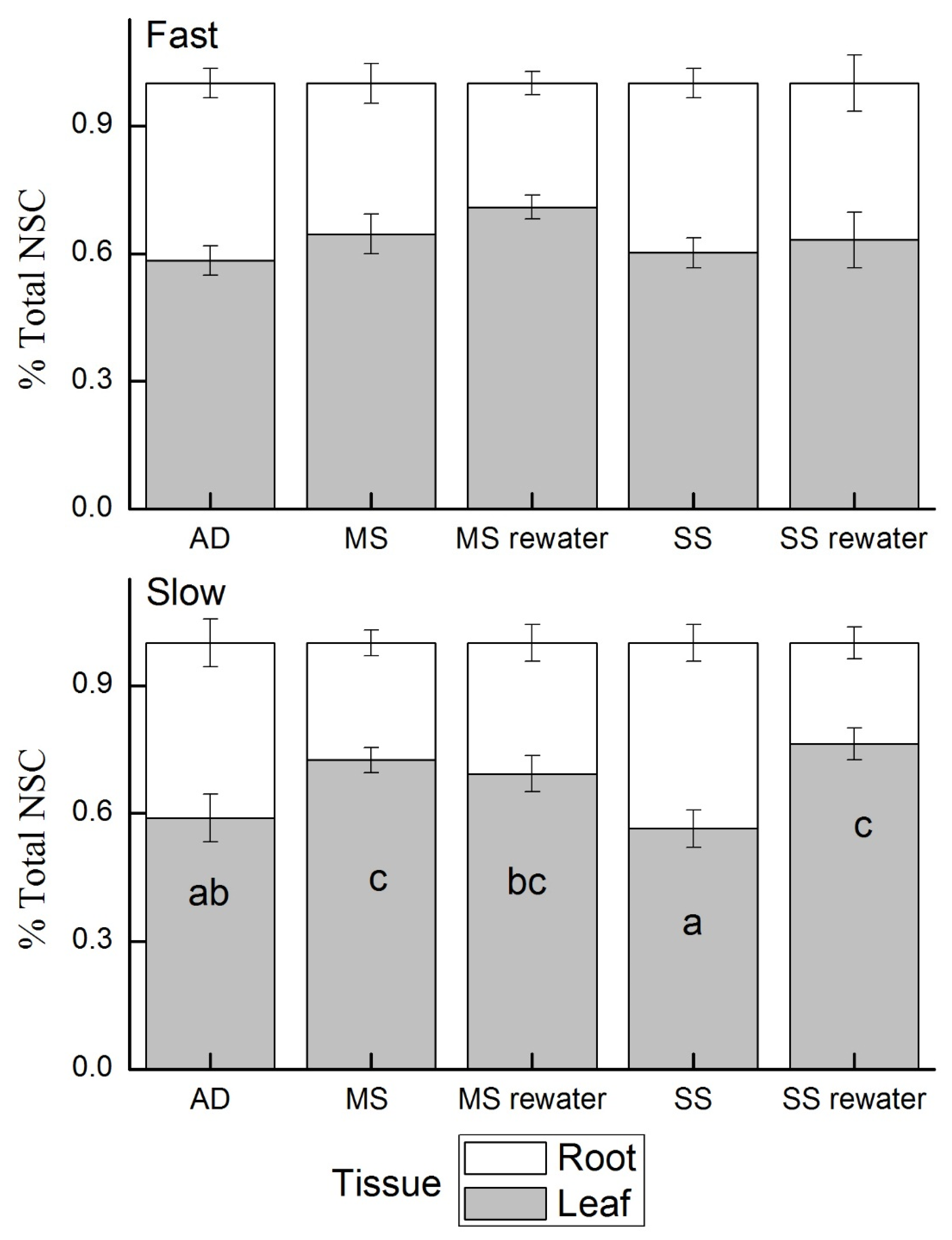
2.7. Nonstructural Carbohydrates (NSC)
2.8. Statistical Analysis
3. Results
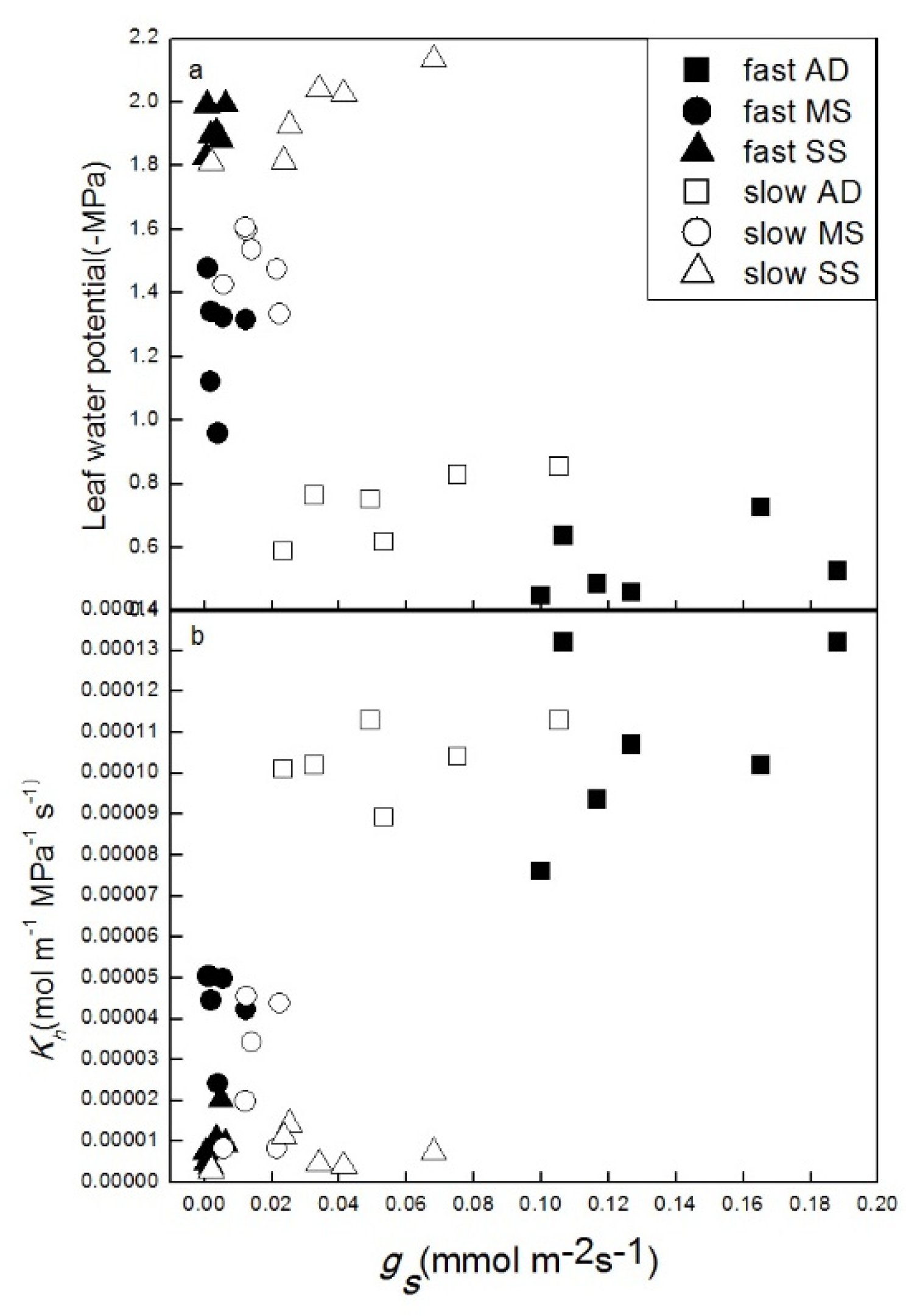
3.1. Influence of Growth Rate on Hydraulic Properties
3.2. NSC Concentration in Relation to Growth Rate and Different Organs
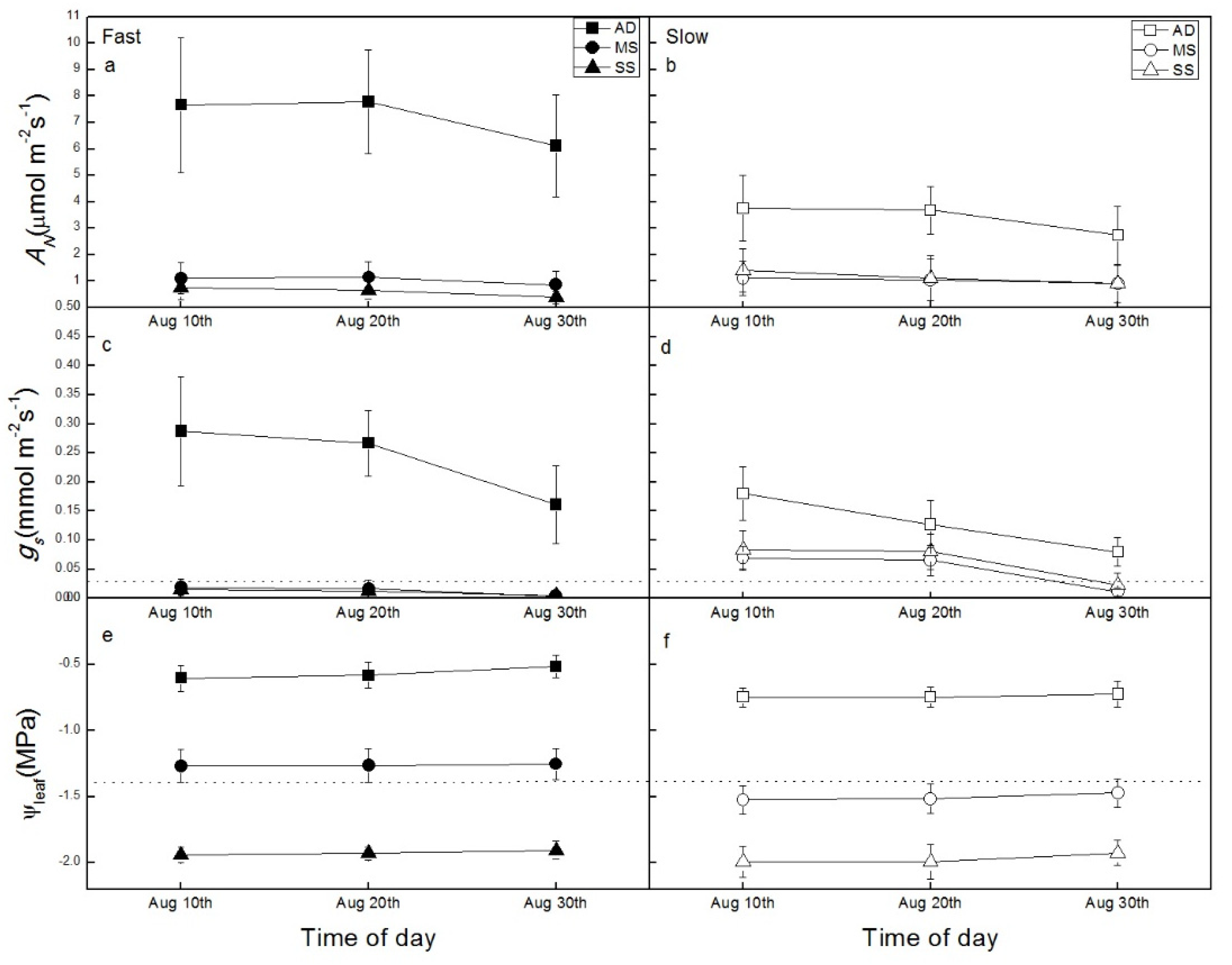
3.3. Relationship between Hydraulic Properties and Gas Exchange
4. Discussion
4.1. Hydraulic Risk
4.2. Carbon Assimilation, Use and Reserves
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, Z.; Lei, X.; Zhu, Q.; Chen, H.; Peng, C. A Drought-Induced Pervasive Increase in Tree Mortality across Canada’s Boreal Forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Joetzjer, E.; Delire, C.; Douville, H.; Ciais, P.; Decharme, B.; Fisher, R.; Christoffersen, B.; Calvet, J.C.; Costa, A.C.L.D.; Ferreira, L.V. Predicting the Response of the Amazon Rainforest to Persistent Drought Conditions under Current and Future Climates: A Major Challenge for Global Land Surface Models. Geosci. Model Dev. Discuss. 2014, 7, 2933–2950. [Google Scholar] [CrossRef]
- D’Orangeville, L.; Maxwell, J.; Kneeshaw, D.; Pederson, N.; Duchesne, L.; Logan, T.; Houle, D.; Arseneault, D.; Beier, C.M.; Bishop, D.A.; et al. Drought Timing and Local Climate Determine the Sensitivity of Eastern Temperate Forests to Drought. Glob. Chang. Biol. 2018, 24, 2339–2351. [Google Scholar] [CrossRef]
- Hrnjak, I.; Lukić, T.; Gavrilov, M.B.; Marković, S.B.; Unkašević, M.; Tošić, I. Aridity in Vojvodina, Serbia. Theor. Appl. Climatol. 2014, 115, 323–332. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap. Science 1983. [Google Scholar] [CrossRef]
- Mencuccini, M. Temporal Scales for the Coordination of Tree Carbon and Water Economies during Droughts. Tree Physiol. 2014, 34, 439–442. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The Interdependence of Mechanisms Underlying Climate-Driven Vegetation Mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Klein, T.; Hoch, G. Tree Carbon Allocation Dynamics Determined Using a Carbon Mass Balance Approach. New Phytol. 2015, 205, 147–159. [Google Scholar] [CrossRef]
- Hartmann, H.; Ziegler, W.; Kolle, O.; Trumbore, S. Thirst Beats Hunger—Declining Hydration during Drought Prevents Carbon Starvation in Norway Spruce Saplings. New Phytol. 2013, 200, 340–349. [Google Scholar] [CrossRef]
- Körner, C. Tansley Lecture: Carbon Limitation in Trees. J. Ecol. 2003, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Woodruff, D.R.; Meinzer, F.C. Carbon Dynamics in Trees: Feast or Famine? Tree Physiol. 2012, 32, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, N.G.; Ryan, M.G.; Zeppel, M.J.; Tissue, D.T. Feature: Feature: Improving Our Knowledge of Drought-Induced Forest Mortality through Experiments, Observations, and Modeling. New Phytol. 2013, 200, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Williams, M.; Do Vale, R.L.; Da Costa, A.L.; MEIR, P. Evidence from Amazonian Forests is Consistent with Isohydric Control of Leaf Water Potential. Plant Cell Environ. 2006, 29, 151–165. [Google Scholar] [CrossRef] [PubMed]
- West, A.G.; Hultine, K.R.; Sperry, J.S.; Bush, S.E.; Ehleringer, J.R. Transpiration and Hydraulic Strategies in a Pinon-Juniper Woodland. Ecol. Appl. 2008, 18, 911–927. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Anderegg, L.D.L. Hydraulic and Carbohydrate Changes in Experimental Drought-Induced Mortality of Saplings in Two Conifer Species. Tree Physiol. 2013, 33, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacke, U.G.; Sperry, J.S. Limits to Xylem Refilling under Negative Pressure in Laurus nobilis and Acer negundo. Plant Cell Environ. 2003, 26, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Ameglio, T.; Bodet, C.; Lacointe, A.; Cochard, H. Winter Embolism, Mechanisms of Xylem Hydraulic Conductivity Recovery and Springtime Growth Patterns in Walnut and Peach Trees. Tree Physiol. 2002, 22, 1211–1220. [Google Scholar] [CrossRef] [Green Version]
- Alves, G.; Ameglio, T.; Guilliot, A.; Fleurat-Lessard, P.; Lacointe, A.; Sakr, S.; Petel, G.; Julien, J.L. Winter Variation in Xylem Sap pH of Walnut Trees: Involvement of plasma membrane H+-ATPase of vessel-associated cells. Tree Physiol. 2004, 24, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Salleo, S.; Trifilò, P.; Lo Gullo, M.A. Phloem as a Possible Major Determinant of Rapid Cavitation Reversal in Stems of Laurus nobilis (Laurel). Funct. Plant Biol. 2006, 33, 1063. [Google Scholar] [CrossRef]
- Salleo, S.; Gullo, M.A.L.; Trfilo, P.; Nardini, A. New Evidence for a Role of Vessel-Associated Cells and Phloem in the Rapid Xylem Refilling of Cavitated Stems of Laurus nobilis L. Plant Cell Environ. 2004, 27, 1065–1076. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T. Variability among Species of Stomatal Control under Fluctuating Soil Water Status and Evaporative Demand: Modelling Isohydric and Anisohydric Behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A New Look at Water Transport Regulation in Plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelton, R.P.; West, A.G.; Dawson, T.E. Predicting Plant Vulnerability to Drought in Biodiverse Regions Using Functional Traits. Proc. Natl. Acad. Sci. USA 2015, 112, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant Responses to Rising Vapor Pressure Deficit. New Phytol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Rogiers, S.Y.; Greer, D.H.; Hatfield, J.M.; Hutton, R.J.; Clarke, S.J.; Hutchinson, P.A.; Somers, A. Stomatal Response of an Anisohydric Grapevine Cultivar to Evaporative Demand, Available Soil Moisture and Abscisic Acid. Tree Physiol. 2012, 32, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Conesa, M.R.; de la Rosa, J.M.; Domingo, R.; Bañon, S.; Pérez-Pastor, A. Changes Induced by Water Stress on Water Relations, Stomatal Behaviour and Morphology of Table Grapes (Cv. Crimson Seedless) Grown in Pots. Sci. Hortic. 2016, 202, 9–16. [Google Scholar] [CrossRef]
- Schultz, H.R. Differences in Hydraulic Architecture Account for Near-Isohydric and Anisohydric Behaviour of Two Field-Grown Vitis vinifera L. Cultivars during Drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Franks, P.J.; Drake, P.L.; Froend, R.H. Anisohydric but Isohydrodynamic: Seasonally Constant Plant Water Potential Gradient Explained by a Stomatal Control Mechanism Incorporating Variable Plant Hydraulic Conductance. Plant Cell Environ. 2007, 30, 19–30. [Google Scholar] [CrossRef]
- Garcia-Forner, N.; Adams, H.D.; Sevanto, S.; Collins, A.D.; Dickman, L.T.; Hudson, P.J.; Zeppel, M.J.B.; Jenkins, M.W.; Powers, H.; Martínez-Vilalta, J.; et al. Responses of Two Semiarid Conifer Tree Species to Reduced Precipitation and Warming Reveal New Perspectives for Stomatal Regulation. Plant Cell Environ. 2016, 39, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.L.; Sterck, F.J.; Martínez-Vilalta, J.; Villar, R. Water-use Strategies of Six Co-existing Mediterranean Woody Species during a Summer Drought. Oecologia 2011, 166, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.S.; Wang, J.; Zhou, Y.X.; Liu, Y.H.; Zheng, S.X. Studies on Drought-Resistance of Five Poplar Clones. J. Northwest A F Univ. (Nat. Sci. Ed.) 2005, 112–116. [Google Scholar] [CrossRef]
- Scholander, P.F. The Role of Solvent Pressure in Osmotic Systems. Proc. Natl. Acad. Sci. USA 1966, 55, 1407–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyree, M.T. Relationship to a Field Assessment of Drought Performance. Plant Physiol. 2003, 132, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Damour, G.; Bodet, C.; Tharwat, I.; Poirier, M.; Améglio, T. Evaluation of a New Centrifuge Technique for Rapid Generation of Xylem Vulnerability Curves. Physiol. Plant. 2005, 124, 410–418. [Google Scholar] [CrossRef]
- Cai, J.; Tyree, M.T. The Impact of Vessel Size on Vulnerability Curves: Data and Models for within-Species Variability in Saplings of Aspen, Populus tremuloides Michx. Plant Cell Environ. 2010, 33, 1059–1069. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752. [Google Scholar] [CrossRef] [Green Version]
- Fichot, R.; Brignolas, F.; Cochard, H.; Ceulemans, R. Vulnerability to Drought-Induced Cavitation in Poplars: Synthesis and Future Opportunities. Plant Cell Environ. 2015, 38, 1233–1251. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Oren, R.; Comstock, J.P. Water Deficits and Hydraulic Limits to Leaf Water Supply. Plant Cell Environ. 2002, 25, 251–263. [Google Scholar] [CrossRef]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; White, D.A.; Ottenschlaeger, M.L.; Pinkard, E.A. Drought Response Strategies Define the Relative Contributions of Hydraulic Dysfunction and Carbohydrate Depletion during Tree Mortality. New Phytol. 2013, 197, 862–872. [Google Scholar] [CrossRef]
- Ebell, L.F. Specific Total Starch Determinations in Conifer Tissues with Glucose Oxidase. Phytochemistry 1969, 8, 25–36. [Google Scholar] [CrossRef]
- Qian, L.W.; Zhang, X.S.; Yang, Z.J.; Han, Z.G. Comparison of Different Light Response Models for Photosynthesis. J. Wuhan Bot. Res. 2009, 2, 197–203. [Google Scholar] [CrossRef]
- Ogasa, M.; Miki, N.H.; Murakami, Y.; Yoshikawa, K. Recovery Performance in Xylem Hydraulic Conductivity is Correlated with Cavitation Resistance for Temperate Deciduous Tree Species. Tree Physiol. 2013, 33, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le Thiec, D.; Brechet, C.; Brignolas, F. Impact of Drought on Productivity and Water Use Efficiency in 29 Genotypes of Populus deltoides X Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Klein, T. The Variability of Stomatal Sensitivity to Leaf Water Potential across Tree Species Indicates a Continuum between Isohydric and Anisohydric Behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of Plant Survival and Mortality during Drought: Why do Some Plants Survive while Others Succumb to Drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Shackel, K.A.; Ahmadi, H.; Biasi, W.; Buchner, R.; Yeager, J. Plant water status as an index of irrigation need in deciduous fruit trees. Horttechnology 1997, 7, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Urli, M.; Porte, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem Embolism Threshold for Catastrophic Hydraulic Failure in Angiosperm Trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- Tyree, M.T.; Sperry, J.S. Vulnerability of Xylem to Cavitation and Embolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2003, 40, 19–38. [Google Scholar] [CrossRef]
- Pockman, W.T.; Sperry, J.S. Vulnerability to Xylem Cavitation and the Distribution of Sonoran Desert Vegetation. Am. J. Bot. 2000, 87, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Casella, E.; Mencuccini, M. Xylem Vulnerability to Cavitation Varies among Poplar and Willow Clones and Correlates with Yield. Tree Physiol. 2007, 27, 1761–1767. [Google Scholar] [CrossRef] [PubMed]
- Dalla-Salda, G.; Martinez-Meier, A.; Cochard, H.; Rozenberg, P. Variation of Wood Density and Hydraulic Properties of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) Clones Related to a Heat and Drought Wave in France. For. Ecol. Manag. 2009, 257, 182–189. [Google Scholar] [CrossRef]
- Sangsing, K.; Kasemsap, P.; Thanisawanyangkura, S.; Sangkhasila, K.; Gohet, E.; Thaler, P.; Cochard, H. Xylem Embolism and Stomatal Regulation in Two Rubber Clones (Hevea brasiliensis Muell. Arg.). Trees 2004, 18, 109–114. [Google Scholar] [CrossRef]
- Wikberg, J.; Gren, E. Interrelationships between Water Use and Growth Traits in Biomass-Producing Willows. Trees 2004, 18, 70–76. [Google Scholar] [CrossRef]
- Fichot, R.; Barigah, T.S.; Chamaillard, S.; Thiec, D.L.; Laurans, F.; Cochard, H.; Brignolas, F. Common trade-offs between Xylem Resistance to Cavitation and Other Physiological Traits do not Hold among Unrelated Populus deltoides × Populus nigra Hybrids. Plant Cell Environ. 2010, 33, 1553–1568. [Google Scholar] [CrossRef]
- Awad, H.; Barigah, T.; Badel, E.; Cochard, H.; Herbette, S. Poplar Vulnerability to Xylem Cavitation Acclimates to Drier Soil Conditions. Physiol. Plant. 2010, 139, 280–288. [Google Scholar] [CrossRef]
- Hajek, P.; Leuschner, C.; Hertel, D.; Delzon, S.; Schuldt, B. Trade-offs between Xylem Hydraulic Properties, Wood Anatomy and Yield in Populus. Tree Physiol. 2014, 34, 744–756. [Google Scholar] [CrossRef]
- Barigah, T.S.; Charrier, O.; Douris, M.; Bonhomme, M.; Herbette, S.; Améglio, T.; Fichot, R.; Brignolas, F.; Cochard, H. Water Stress-Induced Xylem Hydraulic Failure is a Causal Factor of Tree Mortality in Beech and Poplar. Ann. Bot.-Lond. 2013, 112, 1431–1437. [Google Scholar] [CrossRef]
- Moshelion, M.; Halperin, O.; Wallach, R.; Oren, R.; Way, D.A. Role of Aquaporins in Determining Transpiration and Photosynthesis in Water-stressed Plants: Crop Water-use Efficiency, Growth and Yield. Plant Cell Environ. 2015, 38, 1785–1793. [Google Scholar] [CrossRef]
- Hao, S.; Zhao, T.; Xia, X.; Yin, W. Genome-wide Comparison of Two poplar Genotypes with Different Growth Rates. Plant Mol. Biol. 2011, 76, 575–591. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural Carbon in Woody Plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, E.; Helliker, B. A Re-Evaluation of Carbon Storage in Trees Lends Greater Support for Carbon Limitation to Growth. New Phytol. 2012, 195, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H. Plant Responses to Water Stress. Ann. Bot.-Lond. 2002, 89, 801–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevanto, S.; McDowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A Test of the Hydraulic Failure and Carbon Starvation Hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Galiano, L.; Timofeeva, G.; Saurer, M.; Siegwolf, R.; Martínez-Vilalta, J.; Hommel, R.; Gessler, A. The Fate of Recently Fixed Carbon after Drought Release: Towards Unravelling C Storage Regulation in Tilia platyphyllos and Pinus sylvestris. Plant Cell Environ. 2017, 40, 1711–1724. [Google Scholar] [CrossRef]
- Huang, J.; Hammerbacher, A.; Weinhold, A.; Reichelt, M.; Gleixner, G.; Behrendt, T.; van Dam, N.M.; Sala, A.; Gershenzon, J.; Trumbore, S.; et al. Eyes on the Future-evidence for Trade-offs between Growth, Storage and Defense in Norway Spruce. New Phytol. 2019, 222, 144–158. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Burslem, D.F.R.P.; Caduff, A.; Tay, J.; Hector, A. Contrasting Nonstructural Carbohydrate Dynamics of Tropical Tree Seedlings under Water Deficit and Variability. New Phytol. 2015, 205, 1083–1094. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Rate | AD Plants | MS Plants | SS Plants | MS Rewater | SS Rewater |
|---|---|---|---|---|---|---|
| Height increment (%) | Fast | 21.76(6.3) Aa | 15.66(3.5) Aa | 11.36(1.7) Ab | 12.524(1.3) Aab | 10.156(1.7) Aab |
| Slow | 18.21(5.1) Aa | 9.16(2) Bc | 8.333(1.6) Bc | 13.384(1.5) Abc | 9.885(1.1) Abc | |
| P50 (-MPa) | Fast | 2.15 ± 0.08 Aa | 2.28 ± 0.04 Aab | 2.45 ± 0.19 Ac | 2.23 ± 0.07 Aab | 2.32 ± 0.09 Abc |
| Slow | 2.35 ± 0.13 Aa | 2.6 ± 0.25 Bb | 2.82 ± 0.17 Bc | 2.6 ± 0.21 Ab | 2.8 ± 0.25 Bc | |
| P12 (-MPa) | Fast | 1.12 ± 0.12 Aa | 1.70 ± 0.11 Ac | 1.88 ± 0.13 Ac | 1.33 ± 0.15 Ab | 1.88 ± 0.04 Ac |
| Slow | 1.33 ± 0.11 Ba | 2.05 ± 0.09 Bb | 2.26 ± 0.05 Bc | 1.87 ± 0.15 Bb | 2.13 ± 0.09 Bbd | |
| P88 (-MPa) | Fast | 2.53 ± 0.05 Aa | 2.71 ± 0.09 Ab | 2.87 ± 0.12 Ac | 2.65 ± 0.12 Aab | 2.62 ± 0.09 Aab |
| Slow | 2.75 ± 0.13 Ba | 3.03 ± 0.12 Bb | 3.32 ± 0.10 Bc | 3.33 ± 0.11 Bc | 3.45 ± 0.08 Bc | |
| Specific conductivity (mol m−1 MPa−1 s−1) | Fast | 1.02 × 10−4 ± 2.03 × 10−5 Aa | 4.22 × 10−5 ± 1.07 × 10−5 Ab | 9.43 × 10−6 ± 5.89 × 10−6 Ac | 2.90 × 10−5 ± 1.75 × 10−5 Abc | 2.03 × 10−5 ± 1.48 × 10−5 Abc |
| Slow | 1.02 × 10−4 ± 8.65 × 10−6 Aa | 3.43 × 10−5 ± 1.73 × 10−5 Ab | 7.44 × 10−6 ± 4.48 × 10−6 Bc | 2.14 × 10−5 ± 1.37 × 10−5 Abc | 1.05 × 10−5 ± 1.96 × 10−6 Abc | |
| Embolism level (%) | Fast | 14.28 ± 3.07 Aa | 44.58 ± 5.62 Ab | 85.77 ± 5.28 Ab | 40.15 ± 5.51 Ab | 59.36 ± 6.86 Ab |
| Slow | 16.02 ± 2.53 Aa | 54.98 ± 4.92 Ab | 85.73 ± 3.69 Ac | 50.71 ± 4.85 Ab | 71.62 ± 4.59 Bd | |
| hydraulic safety margin (Mpa) | Fast | 0.704 ± 0.01 Aa | 0.49 ± 0.008 Ab | 0.38 ± 0.01 Ac | 0.83 ± 0.007 Ad | 1.07 ± 0.09 Ae |
| Slow | 0.81 ± 0.03 Ba | 0.6 ± 0.03 Bb | 0.59 ± 0.01 Bc | 1.25 ± 0.02 Bd | 1.44 ± 0.13 Bd |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Liu, L.; Zhao, H.; Jiang, Z.; Cai, J. Differences in Near Isohydric and Anisohydric Behavior of Contrasting Poplar Hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under Drought-Rehydration Treatments. Forests 2020, 11, 402. https://doi.org/10.3390/f11040402
Zhang L, Liu L, Zhao H, Jiang Z, Cai J. Differences in Near Isohydric and Anisohydric Behavior of Contrasting Poplar Hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under Drought-Rehydration Treatments. Forests. 2020; 11(4):402. https://doi.org/10.3390/f11040402
Chicago/Turabian StyleZhang, Li, Li Liu, Han Zhao, Zaimin Jiang, and Jing Cai. 2020. "Differences in Near Isohydric and Anisohydric Behavior of Contrasting Poplar Hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under Drought-Rehydration Treatments" Forests 11, no. 4: 402. https://doi.org/10.3390/f11040402
APA StyleZhang, L., Liu, L., Zhao, H., Jiang, Z., & Cai, J. (2020). Differences in Near Isohydric and Anisohydric Behavior of Contrasting Poplar Hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under Drought-Rehydration Treatments. Forests, 11(4), 402. https://doi.org/10.3390/f11040402