Roads Impact Tree and Shrub Productivity in Adjacent Boreal Peatlands


Abstract
:1. Introduction
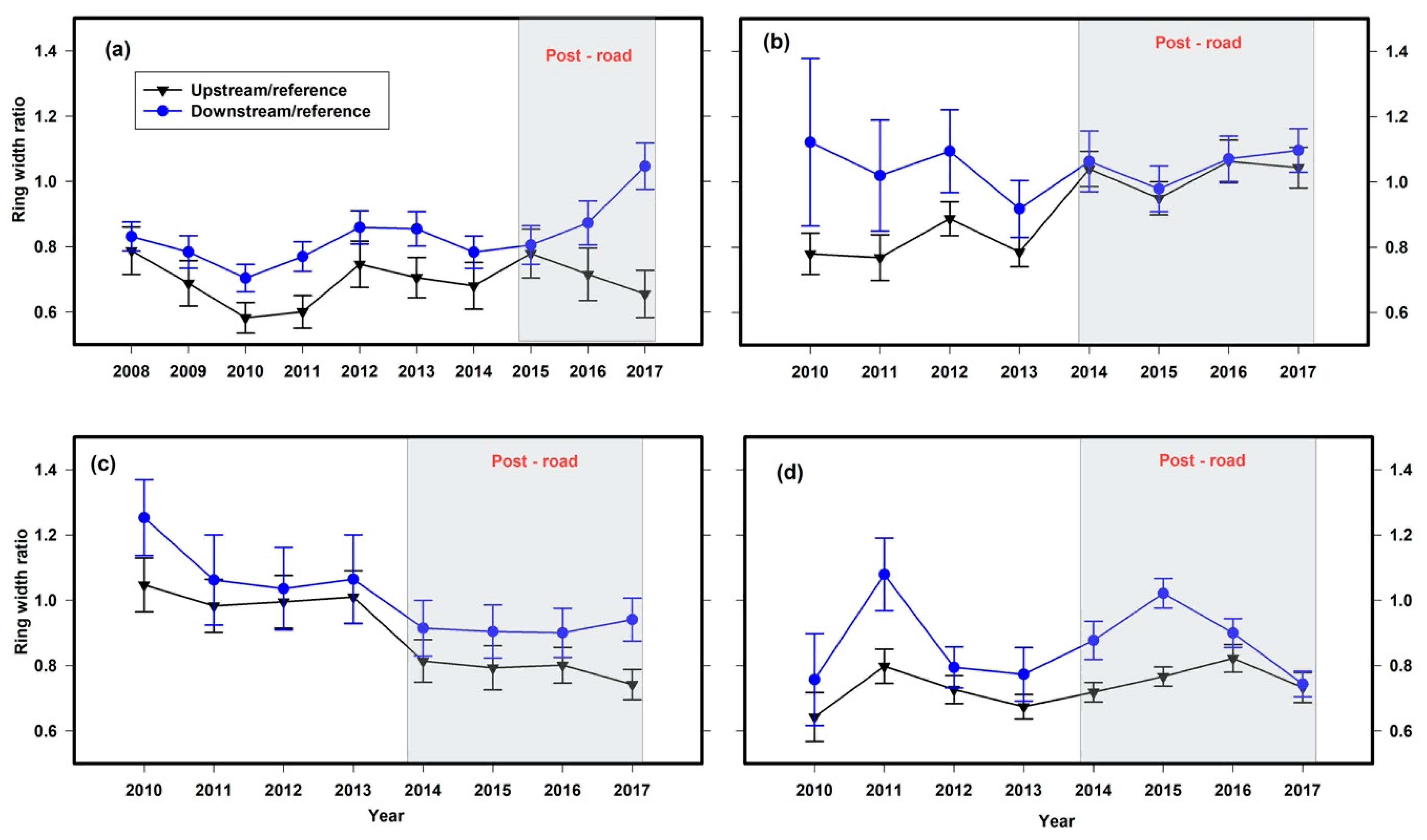
- On the upstream side of the road, the annual radial growth of trees and tall shrubs would be significantly less than the downstream side of the road.
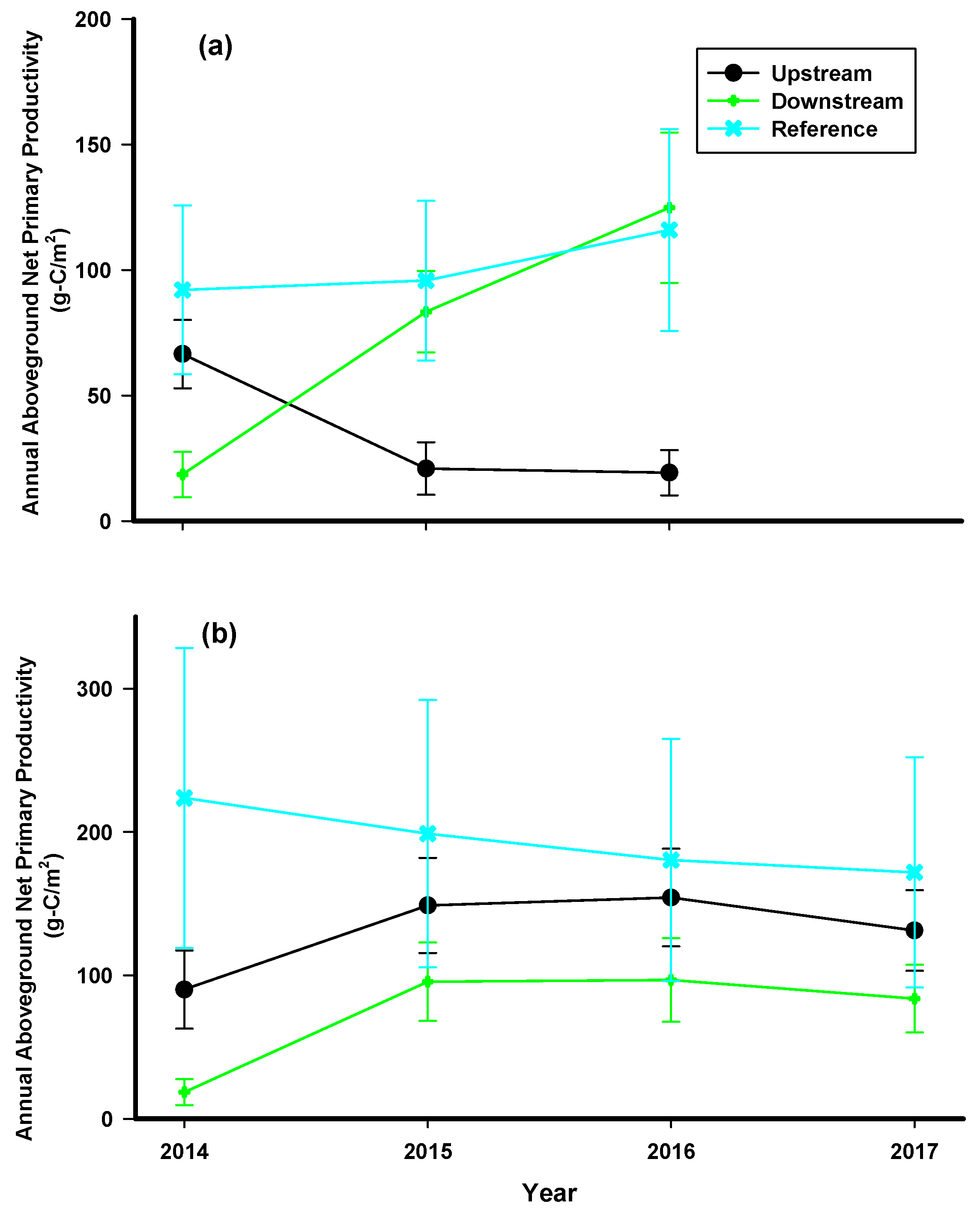
- The NPPag of trees and tall shrubs would be significantly reduced by vegetation clearing during road construction.
- The greatest impact of the road would be observed closest to the road where hydrologic impacts and clearing of vegetation have been observed to be most severe.
2. Materials and Methods
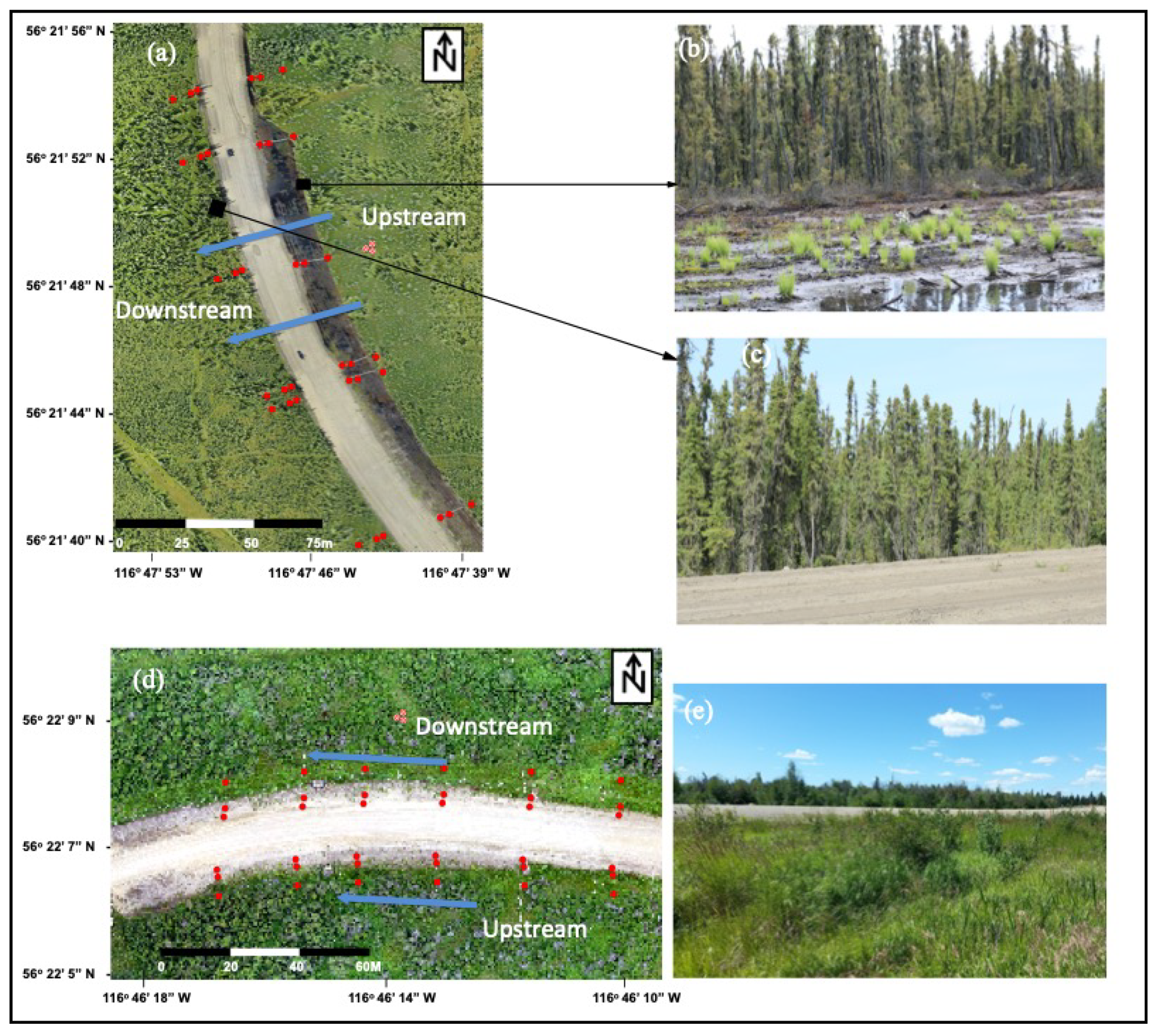
2.1. Study Sites
2.2. Study Design and Data Collection
2.3. Ring Width Processing and Biomass Estimation
2.4. Annual NPPag Estimation
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitt, D.; Halsey, L.; Bauer, I.; Campbell, C. Spatial and temporal trends in carbon storage of peatlands of continental western Canada through the Holocene. Can. J. Earth Sci. 2000, 37, 683–693. [Google Scholar] [CrossRef]
- Page, S.E.; Reiley, J.O.; Banks, C.J. Global and regional importance of the tropical peatland carbon pool. Glob. Chang. Biol. 2011, 17, 798–818. [Google Scholar] [CrossRef] [Green Version]
- Saraswati, S.; Petrone, R.M.; Mustafiz, R.; McDermid, G.J.; Xu, B.; Strack, M. Hydrological effects of resource-access road crossings on boreal forested peatlands. J. Hydrol. 2020, 584, 124748. [Google Scholar] [CrossRef]
- Alm, J.; Schulman, L.; Walden, J.; Nykanen, H.; Martikainen, P.J.; Silvola, J. Carbon balance of a boreal bog during a year with an exceptionally dry summer. Ecology 1999, 80, 161–174. [Google Scholar] [CrossRef]
- Petrone, R.M.; Solondz, D.S.; Macrae, M.L.; Gignac, D.; Devito, K.J. Microtopographical and canopy cover controls on moss carbon dioxide exchange in a western Boreal Plain peatland. Ecohydrology 2011, 4, 115–129. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Pastor, J.; Harth, C.; Bridgham, S.D.; Updegraff, K.; Chapin, C.T. Response of Bog and Fen Plant Communities to Warming and Water-Table Manipulations. Ecology 2000, 81, 3464–3478. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; ISBN 9781118676820. [Google Scholar]
- Laiho, R. Decomposition in peatlands: Reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biol. Biochem. 2006, 38, 2011–2024. [Google Scholar] [CrossRef]
- Clymo, R.S.; Pearce, D.M.E.; Conrad, R. Methane and Carbon Dioxide Production in, Transport through, and Efflux from a Peatland. Philos. Trans. R. Soc. 1995, 351, 249–259. [Google Scholar]
- Gorham, E. Northern peatlands: Role in the carbon cycle and probably responses to climate warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef]
- Freeman, C.; Ostle, N.; Kang, H. An enzymic “latch” on a global carbon store. Nature 2001, 409, 149. [Google Scholar] [CrossRef]
- Keim, R.F.; Amos, J.B. Dendrochronological analysis of baldcypress (Taxodium distichum) responses to climate and contrasting flood regimes. Can. J. For. Res. 2012, 42, 423–436. [Google Scholar] [CrossRef]
- Tardif, J.; Bergeron, Y. Comparative dendroclimatological analysis of two black ash and two white cedar populations from contrasting sites in the Lake Duparquet region, northwestern Quebec. Can. J. For. Res. 1997, 27, 108–116. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Rothwell, R.L. Rooting of peatland black spruce and tamarack in relation to depth of water table. Can. J. Bot. 1987, 65, 817–821. [Google Scholar] [CrossRef]
- Dymond, S.F.; D’Amato, A.W.; Kolka, R.K.; Bolstad, P.V.; Sebestyen, S.D.; Gill, K.; Curzon, M.T. Climatic controls on peatland black spruce growth in relation to water table variation and precipitation. Ecohydrology 2019, 12, e2137. [Google Scholar] [CrossRef] [Green Version]
- Dang, Q.L.; Lieffers, V.J. Climate and annual ring growth of black spruce in some Alberta peatlands. Can. J. Bot. 1989, 67, 1885–1889. [Google Scholar] [CrossRef]
- Munir, T.M.; Perkins, M.; Kaing, E.; Strack, M. Carbon dioxide flux and net primary production of a boreal treed bog: Responses to warming and water-table-lowering simulations of climate change. Biogeosciences 2015, 12, 1091–1111. [Google Scholar] [CrossRef] [Green Version]
- Blodau, C.; Moore, T.R. Experimental response of peatland carbon dynamics to a water table fluctuation. Aquat. Sci.-Res. Across Boundaries 2003, 65, 47–62. [Google Scholar] [CrossRef]
- Charman, D. Peatlands and Environmental Change.; John Wiley & Sons Ltd: Chichester, UK, 2002; ISBN 0470844108. [Google Scholar]
- Iversen, C.M.; Childs, J.; Norby, R.J.; Ontl, T.A.; Kolka, R.K.; Brice, D.J.; McFarlane, K.J.; Hanson, P.J. Fine-root growth in a forested bog is seasonally dynamic, but shallowly distributed in nutrient-poor peat. Plant Soil 2018, 424, 123–143. [Google Scholar] [CrossRef]
- Hsueh, Y.H.; Chambers, J.L.; Krauss, K.W.; Allen, S.T.; Keim, R.F. Hydrologic exchanges and baldcypress water use on deltaic hummocks, Louisiana, USA. Ecohydrology 2016, 9, 1452–1463. [Google Scholar] [CrossRef]
- Strack, M.; Waller, M.F.; Waddington, J.M. Sedge Succession and Peatland Methane Dynamics: A Potential Feedback to Climate Change. Ecosystems 2006, 9, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Benscoter, B.W.; Turetsky, M.R. The effect of long-term drying associated with experimental drainage and road construction on vegetation composition and productivity in boreal fens. Wetl. Ecol. Manag. 2015, 23, 845–854. [Google Scholar] [CrossRef]
- Plach, J.M.; Wood, M.E.; Macrae, M.L.; Osko, T.J.; Petrone, R.M. Effect of a semi-permanent road on N, P, and CO2 dynamics in a poor fen on the Western Boreal Plain, Canada. Ecohydrology 2017, 10, e1874. [Google Scholar] [CrossRef]
- Wood, M.E.; Macrae, M.L.; Strack, M.; Price, J.S.; Osko, T.J.; Petrone, R.M. Spatial variation in nutrient dynamics among five different peatland types in the Alberta oil sands region. Ecohydrology 2016, 9, 688–699. [Google Scholar] [CrossRef]
- Caners, R.T.; Lieffers, V.J. Divergent Pathways of Successional Recovery for In Situ Oil Sands Exploration Drilling Pads on Wooded Moderate-Rich Fens in Alberta, Canada. Restor. Ecol. 2014, 22, 657–667. [Google Scholar] [CrossRef]
- Vitt, D.H.; Wieder, R.K.; Xu, B.; Kaskie, M.; Koropchak, S. Peatland establishment on mineral soils: Effects of water level, amendments, and species after two growing seasons. Ecol. Eng. 2011, 37, 354–363. [Google Scholar] [CrossRef]
- Strack, M.; Hayne, S.; Lovitt, J.; McDermid, G.J.; Rahman, M.M.; Saraswati, S.; Xu, B. Petroleum exploration increases methane emissions from northern peatlands. Nat. Commun. 2019, 10, 2804. [Google Scholar] [CrossRef] [Green Version]
- Graf, M.D. Literature Review on the Restoration of Alberta’s Boreal Wetlands: Affected by Oil, Gas and in Situ Oil Sands Development; Ducks Unlimited Canada: Edmonton, AB, Canada, 2009. [Google Scholar]
- Bocking, E.; Cooper, D.J.; Price, J. Using tree ring analysis to determine impacts of a road on a boreal peatland. For. Ecol. Manage. 2017, 404, 24–30. [Google Scholar] [CrossRef]
- Rooney, R.C.; Bayley, S.E.; Schindler, D.W. Oil sands mining and reclamation cause massive loss of peatland and stored carbon. Proc. Natl. Acad. Sci. USA 2012, 109, 4933–4937. [Google Scholar] [CrossRef] [Green Version]
- Pasher, J.; Seed, E.; Duffe, J. Development of boreal ecosystem anthropogenic disturbance layers for Canada based on 2008 to 2010 Landsat imagery. Can. J. Remote Sens. 2013, 39, 42–58. [Google Scholar] [CrossRef]
- Partington, M.; Gillies, C.; Gingras, B.; Smith, C.; Morissette, J. Resource Roads and Wetlands: A Guide for Planning, Construction and Maintenance; FPInnovations: Quebec City, QC, Canada, 2016; ISBN 978-0-86488-573-9. [Google Scholar]
- Willier, C. Changes in Peatland Plant Communitity Composition and Stand Structure Due to Road Induced Flooding and Desiccation. Mater’s Thesis, University of Alberta, Edmonton, AB, Canada, 2017. [Google Scholar]
- Environment and Climate Change Canada Daily Data Report for May 2014-Climate-Environment and Climate Change Canada. Available online: http://climate.weather.gc.ca/climate_data/daily_data_e.html?StationID=52258&timeframe=2&StartYear=1840&EndYear=2018&Day=11&Year=2014&Month=5# (accessed on 12 September 2018).
- Saraswati, S.; Parsons, C.T.; Strack, M. Access roads impact enzyme activities in boreal forested peatlands. Sci. Total Environ. 2019, 651, 1405–1415. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2019. Available online: https://www.r-project.org/ (accessed on 16 December 2019).
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; M´erian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program Library in R. R package version 1.7.1 2020. Available online: https://github.com/AndyBunn/dplR (accessed on 10 March 2020).
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba. Can. J. For. Res. 2002, 32, 1441–1450. [Google Scholar] [CrossRef]
- He, A.; McDermid, G.J.; Rahman, M.M.; Strack, M.; Saraswati, S.; Xu, B. Developing allometric equations for estimating shrub biomass in a boreal fen. Forests 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- IPCC. 2006 IPCC Guidelines for National Greenhouse Gas Inventories; Eggleston, S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; IGES: Hayama, Japan, 2006; ISBN 4-88788-032-4. [Google Scholar]
- Somogyi, Z.; Cienciala, E.; Mäkipää, R.; Muukkonen, P.; Lehtonen, A.; Weiss, P. Indirect methods of large-scale forest biomass estimation. Eur. J. For. Res. 2007, 126, 197–207. [Google Scholar] [CrossRef]
- Ontl, T.; Iversen, C. SPRUCE S1 Bog Areal Coverage of Hummock and Hollow Microtopography Assessed Along Three Transects in the S1 Bog; U.S. Department of Energy: Oak Ridge, TN, USA, 2017.
- Murphy, M.; Laiho, R.; Moore, T.R. Effects of water table drawdown on root production and aboveground biomass in a boreal Bog. Ecosystems 2009, 12, 1268–1282. [Google Scholar] [CrossRef]
- Campbell, D.; Bergeron, J. Natural Revegetation of Winter Roads on Peatlands in the Hudson Bay Lowland, Canada. Arctic, Antarct. Alp. Res. 2012, 44, 155–163. [Google Scholar] [CrossRef]
- Strack, M.; Softa, D.; Bird, M.; Xu, B. Impact of winter roads on boreal peatland carbon exchange. Glob. Chang. Biol. 2018, 24, e201–e212. [Google Scholar] [CrossRef]
- Tei, S.; Morozumi, T.; Nagai, S.; Takano, S.; Sugimoto, A.; Shingubara, R.; Fan, R.; Fedorov, A.; Gavrilyeva, T.; Tananaev, N.; et al. An extreme flood caused by a heavy snowfall over the Indigirka River basin in Northeastern Siberia. Hydrol. Process. 2020, 34, 522–537. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Kotani, A.; Maximov, T.C. Effects of extreme drought and wet events for tree mortality: Insights from tree-ring width and carbon isotope ratio in a Siberian larch forest. Ecohydrology 2019, 12, e2143. [Google Scholar] [CrossRef]
- Choi, W.-J.; Chang, S.X.; Bhatti, J.S. Drainage affects tree growth and C and N dynamics in a minerotrophic peatland. Ecology 2007, 88, 443–453. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | a | b | Logarithmic Correction Factor |
|---|---|---|---|
| Black spruce 1 | 1.743 | 2.401 | 1.094 |
| Birch 2 | 49.52 | 2.027 | -- |
| Willow 2 | 55.85 | 2.325 | |
| Alder 2 | 44.06 | 2.395 |
| Effects | Black Spruce (R2 = 0.05, F(10, 1464) = 1905.61, p < 0.0001) | |||
|---|---|---|---|---|
| Estimates 2 | Standard Error | t-Values | p Values | |
| Intercept | 0.894 | 0.044 | 21.684 | 0.000 |
| AR-BR | −0.096 | 0.049 | −1.949 | 0.087 |
| D (AR)-U (AR) | −0.178 | 0.055 | −3.264 | 0.001 |
| D (BR)-U (BR) | −0.113 | 0.036 | −3.144 | 0.002 |
| Alder (R2 = 0.09, F(8, 619) = 2062.94, p < 0.0001) | ||||
| Intercept | 1.013 | 0.039 | 26.375 | 0.000 |
| AR-BR | −0.001 | 0.077 | −0.001 | 0.947 |
| D (AR)-U (AR) | 0.054 | 0.051 | 1.059 | 0.389 |
| D (BR)-U (BR) | −0.209 | 0.056 | −3.193 | 0.002 |
| Birch (R2 = 0.10, F(8, 521) = 2001.58, p < 0.0001) | ||||
| Intercept | 0.916 | 0.058 | 17.451 | 0.000 |
| AR-BR | 0.180 | 0.077 | 2.961 | 0.017 |
| D (AR)-U (AR) | −0.137 | 0.061 | −2.114 | 0.030 |
| D (BR)-U (BR) | −0.093 | 0.069 | −1.239 | 0.173 |
| Willow (R2 = 0.09, F(8, 641) = 927.91, p < 0.0001) | ||||
| Intercept | 0.896 | 0.043 | 21.496 | 0.000 |
| AR-BR | −0.092 | 0.062 | −1.950 | 0.091 |
| D (AR)-U (AR) | −0.178 | 0.052 | −3.033 | 0.001 |
| D (BR)-U (BR) | −0.113 | 0.041 | −3.414 | 0.001 |
| Effects | Black Spruce (R2 = 0.20, F(2, 296) = 63.20, p < 0.0001) | |||
|---|---|---|---|---|
| Estimates 2 | Standard Error | t-Values | p Values | |
| Intercept | 0.185 | 0.053 | 3.490 | 0.001 |
| WT | −0.004 | 0.002 | −2.283 | 0.027 |
| BD | 0.024 | 0.009 | 2.721 | 0.007 |
| WT × BD | 0.001 | 0.0003 | 2.590 | 0.010 |
| Alder (R2 = 0.41, F(2, 226) = 372.52, p < 0.0001) | ||||
| Intercept | 1.408 | 0.138 | 10.209 | 0.000 |
| WT | 0.021 | 0.013 | 1.662 | 0.1023 |
| BD | 0.187 | 0.045 | 4.154 | 0.0001 |
| WT × BD | −0.008 | 0.005 | −1.751 | 0.0817 |
| Birch (R2 = 0.37, F (2, 161) = 112.74, p < 0.0001) | ||||
| Intercept | 1.007 | 0.199 | 5.064 | 0.000 |
| WT | −0.034 | 0.013 | −2.588 | 0.011 |
| BD | 0.173 | 0.030 | 5.798 | 0.000 |
| WT × BD | 0.004 | 0.003 | 1.399 | 0.164 |
| Willow (R2 = 0.20, F (1, 239) = 1551.31, p < 0.0001) | ||||
| Intercept | 0.967 | 0.087 | 11.078 | 0.000 |
| WT | 0.009 | 0.007 | 1.284 | 0.201 |
| BD | 0.179 | 0.051 | 3.516 | 0.001 |
| WT × BD | −0.002 | 0.004 | −0.575 | 0.566 |
| Side of the Road | Distance to Plots (m) | NPPag Mean ± SE (g-C/m2) 1 | |
|---|---|---|---|
| Bog | Fen | ||
| Downstream | 2 | 164.7 ± 30.1 a | 29.4 ± 3.5 a |
| Downstream | 6 | 51.9 ± 5.8 b | 56.6 ± 14.1 b |
| Downstream | 20 | 58.2 ± 6.5 b | 187.4 ± 27.4 c |
| Downstream | mean ± SE | 91.6 ± 12.6 d | 91.8 ± 14.7 d |
| Upstream | 2 | 0.0 ± 0.0 c | 110.2 ± 19.2 |
| Upstream | 6 | 0.0 ± 0.0 c | 154.3 ± 20.8 c |
| Upstream | 20 | 58.4 ± 11.8 b | 176.6 ± 33.8 c |
| Upstream | mean ± SE | 19.6 ± 11.3 e | 147.9 ± 14.7 e |
| Reference areas | 101.3 ± 19.4 d | 182.7 ± 41.1 f | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saraswati, S.; Bhusal, Y.; Trant, A.J.; Strack, M. Roads Impact Tree and Shrub Productivity in Adjacent Boreal Peatlands. Forests 2020, 11, 594. https://doi.org/10.3390/f11050594
Saraswati S, Bhusal Y, Trant AJ, Strack M. Roads Impact Tree and Shrub Productivity in Adjacent Boreal Peatlands. Forests. 2020; 11(5):594. https://doi.org/10.3390/f11050594
Chicago/Turabian StyleSaraswati, Saraswati, Yubraj Bhusal, Andrew J. Trant, and Maria Strack. 2020. "Roads Impact Tree and Shrub Productivity in Adjacent Boreal Peatlands" Forests 11, no. 5: 594. https://doi.org/10.3390/f11050594
APA StyleSaraswati, S., Bhusal, Y., Trant, A. J., & Strack, M. (2020). Roads Impact Tree and Shrub Productivity in Adjacent Boreal Peatlands. Forests, 11(5), 594. https://doi.org/10.3390/f11050594