Overstory Effects on the Understory of Aleppo Pine Plantations—Implications for Ecosystem Restoration



Abstract
:1. Introduction
2. Materials and Methods
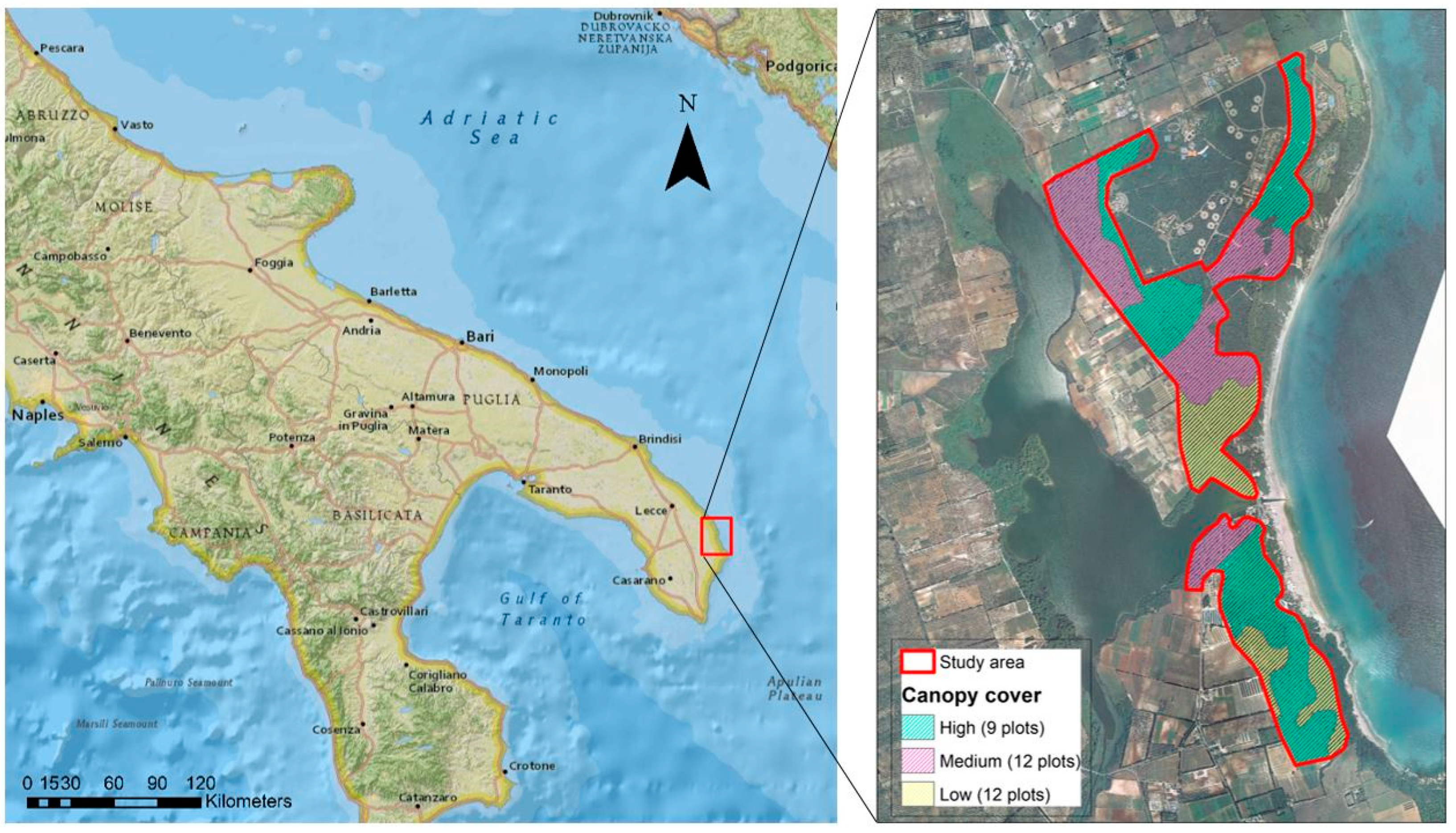
2.1. Study Area
2.2. Sampling
2.3. Data Analysis
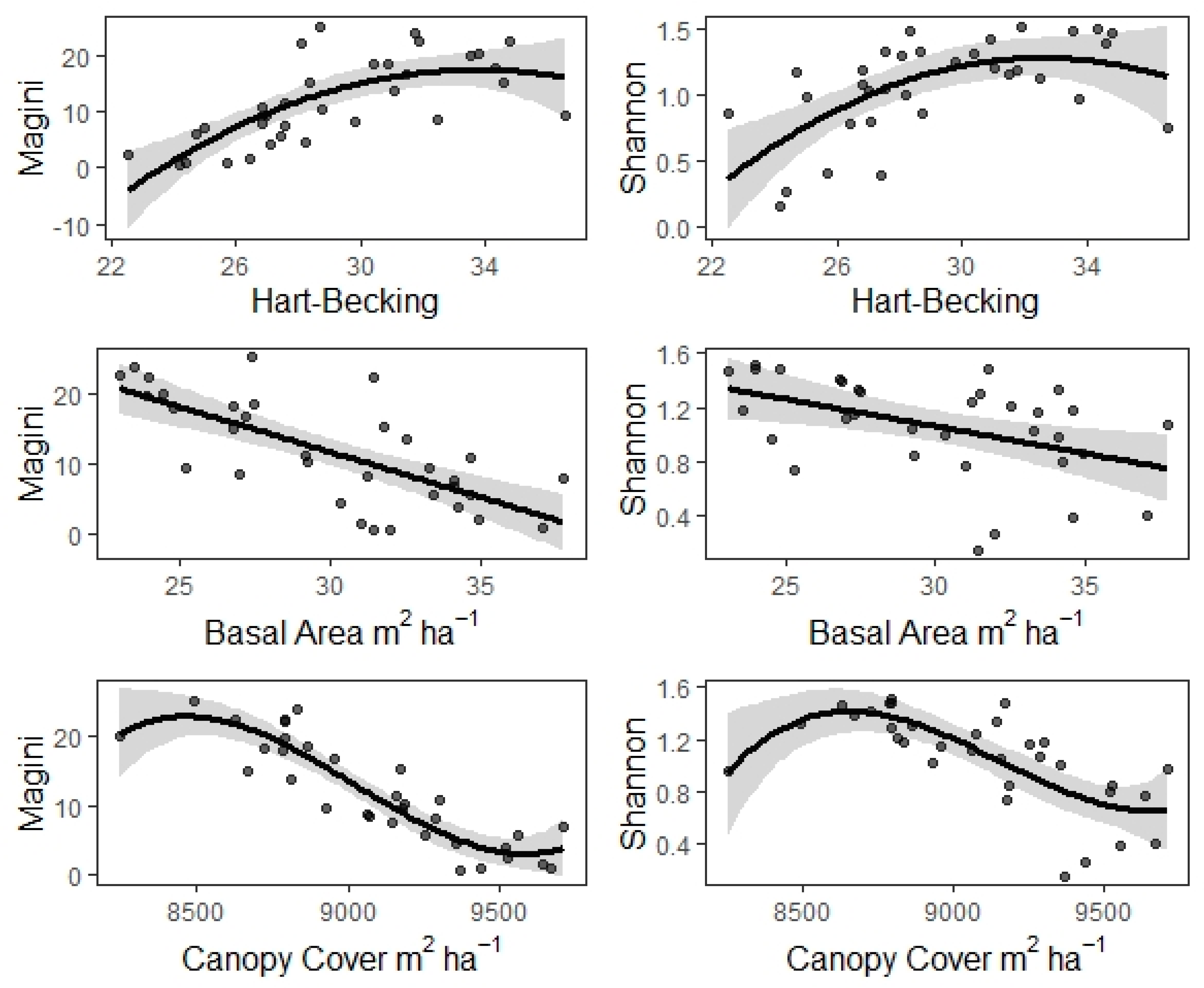
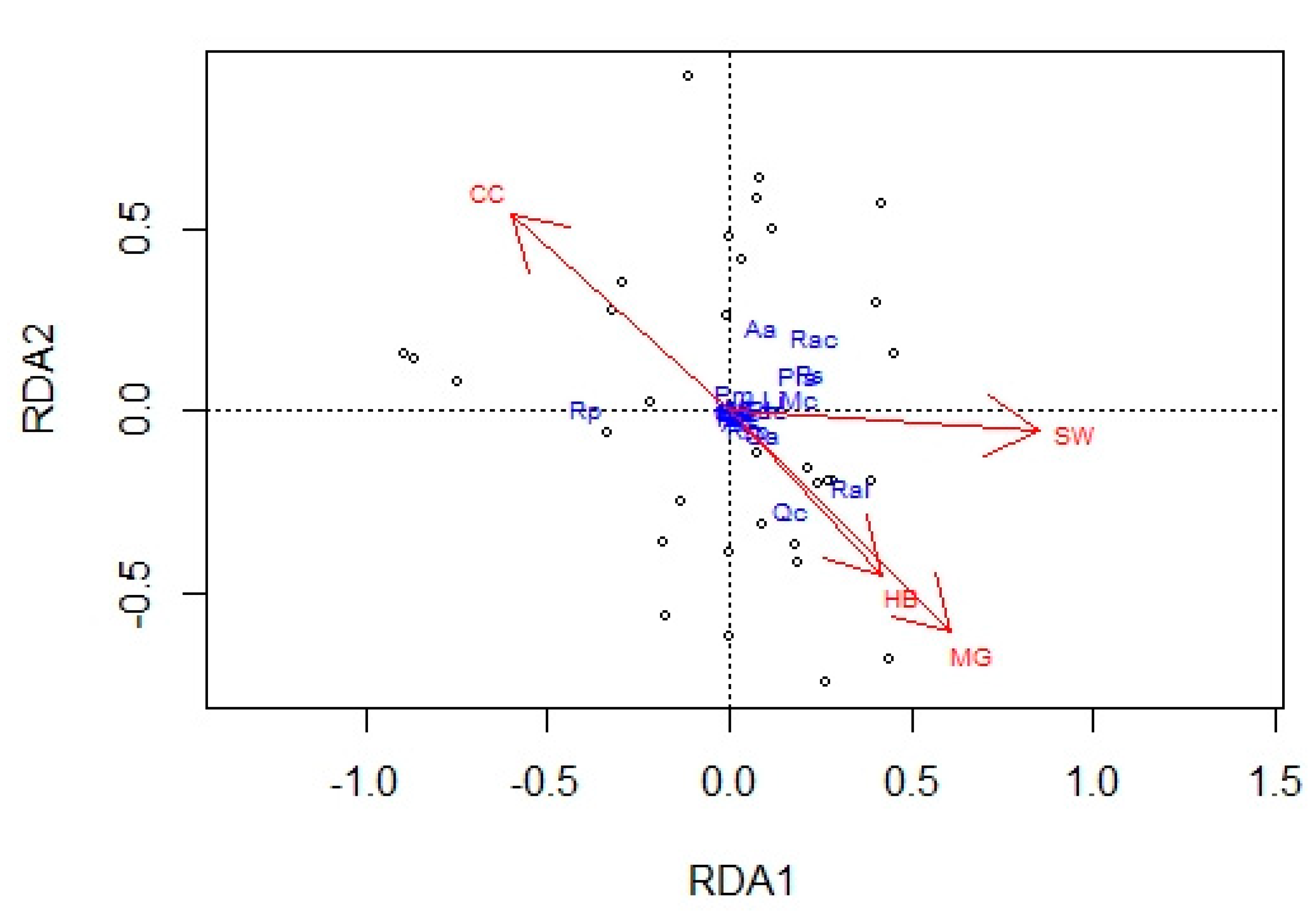
3. Results
4. Discussion
4.1. Overstory Effects on Understory Development
4.2. Overstory Effects on Understory Diversity and Composition
4.3. Management Applications of This Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evans, J. The Multiple Roles of Planted Forests. In Planted Forests: Uses, Impacts, and Sustainability; Evans, J., Ed.; FAO: Rome, Italy, 2009; pp. 61–90. [Google Scholar]
- Nunes, A.; Oliveira, G.; Cabral, M.S.; Branquinho, C.; Correia, O. Beneficial effect of pine thinning in mixed plantations through changes in the understory functional composition. Ecol. Eng. 2014, 70, 387–396. [Google Scholar] [CrossRef]
- Osem, Y.; Zangy, E.; Bney-Moshe, E.; Moshe, Y.; Karni, N.; Nisan, Y. The potential of transforming simple structured pine plantations into mixed Mediterranean forests through natural regeneration along a rainfall gradient. For. Ecol. Manag. 2009, 259, 14–23. [Google Scholar] [CrossRef]
- Pausas, J.G.; Blade, C.; Valdecantos, A.; Seva, J.P.; Fuentes, D.; Alloza, J.A.; Vilagrosa, A.; Bautista, S.; Cortina, J.; Vallejo, R. Pines and oaks in the restoration of Mediterranean landscapes of Spain: New perspectives for an old practice—A review. Plant Ecol. 2004, 171, 209–220. [Google Scholar] [CrossRef]
- Vieira, J.N. Floresta Portuguesa-imagens de tempos idos; Público/Fundação Luso-Americana para o Desenvolvimento/Liga para a Protecção da Natureza: Lisboa, Portugal, 2007; Volume 1, p. 236. [Google Scholar]
- Chauvin, C.; Vallauri, D. Indicateurs de restauration écologique de marnes dégradées dans les Alpes du Sud, 120 ans après reboisement. Revue Écologique 2002, 24, 241–250. [Google Scholar]
- Lahouati, R. Expérience des Plantations en Climat Aride. Cas de la Ceinture Verte en Algérie; Direction Générale des forêts, Ministère de l’Agriculture: Alger, Algeria, 2000. [Google Scholar]
- Ortuño, F. El Plan para la repoblación forestal de España del año 1939. Análisis y comentarios. Ecología 1990, 1, 373–392. [Google Scholar]
- Barbéro, M.; Loisel, R.; Quézel, P.; Richardson, D.M.; Romane, F. Pines as Invaders in the Southern Hemisphere. In Pines of the Mediterranean Basin; Richardson, D.M., Ed.; Cambridge Univesity Press: Cambridge, UK, 2000; pp. 153–170. [Google Scholar]
- Cortina, J.; Maestre, F.T.; Vallejo, R.; Baeza, M.J.; Valdecantos, A.; Pérez-Devesa, M. Ecosystem structure, function, and restoration success: Are they related? J. Nat. Conserv. 2006, 14, 152–160. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For. Ecol. Manag. 2004, 198, 303–317. [Google Scholar] [CrossRef]
- Bellot, J.; Maestre, F.T.; Chirino, E.; Hernández, N.; De Urbina, J.O. Afforestation with Pinus halepensis reduces native shrub performance in a Mediterranean semiarid area. Acta Oecologica 2004, 25, 7–15. [Google Scholar] [CrossRef]
- D’Alessandro, C.M.; Borghetti, M.; Saracino, A. Ecophysiological behaviour of hardwood species in renaturalization processes of coniferous plantations. Ital. J. Forest Mountain Environ. 2005, 60, 429–445. [Google Scholar]
- Puddu, S.; Bianchi, L.; Maltoni, A.; Paci, M.; Tani, A. Preliminary investigations on vegetation dynamics in artificial stands of Pinus radiata in Central Sardinia. Ital. J. Forest Mountain Environ. 2013, 57, 339–352. [Google Scholar]
- Lookingbill, T.R.; Zavala, M.A. Spatial pattern of Quercus ilex and Quercus pubescens recruitment in Pinus halepensis dominated woodlands. J. Veg. Sci. 2000, 11, 607–612. [Google Scholar] [CrossRef]
- Vallejo, V.R.; Arianoutsou, M.; Moreira, F. Fire Ecology and Post-Fire Restoration Approaches in Southern European Forest Types. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 93–119. [Google Scholar]
- Fernandes, P.M.; Luz, A.; Loureiro, C. Changes in wildfire severity from maritime pine woodland to contiguous forest types in the mountains of northwestern Portugal. For. Ecol. Manag. 2010, 260, 883–892. [Google Scholar] [CrossRef]
- Pinto, A.; Fernandes, P.M. Microclimate and modeled fire behavior differ between adjacent forest types in northern Portugal. Forests 2014, 5, 2490–2504. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.R.; Palahi, M.; Trasobares, A.; Pukkala, T. A fire probability model for forest stands in Catalonia (north-east Spain). Ann. Forest Sci. 2006, 63, 169–176. [Google Scholar] [CrossRef]
- Santana, J.; Porto, M.; Reino, L.; Beja, P. Long-term understory recovery after mechanical fuel reduction in Mediterranean cork oak forests. For. Ecol. Manag. 2011, 261, 447–459. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, diversity, and invasion: Implications for conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Masino, S.A.; Faison, E.K. Intact forests in the United States: Proforestation mitigates climate change and serves the greatest good. Front. For. Glob. Chang. 2019, 2. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.L.; Wheeler, C.E.; Mitchard, E.T.A.; Koch, A. Restoring natural forests is the best way to remove atmospheric carbon. Nature 2019, 568, 25–28. [Google Scholar] [CrossRef]
- Casals, P.; Valor, T.; Besalú, A.; Molina-Terrén, D. Understory fuel load and structure eight to nine years after prescribed burning in Mediterranean pine forests. For. Ecol. Manag. 2016, 362, 156–168. [Google Scholar] [CrossRef]
- Fonseca, T.F.; Duarte, J.C. A silvicultural stand density model to control understory in maritime pine stands. iForest Biogeosciences For. 2017, 10, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Coll, L.; González-Olabarria, J.R.; Mola-Yudego, B.; Pukkala, T.; Messier, C. Predicting understory maximum shrubs cover using altitude and overstory basal area in different Mediterranean forests. Eur. J. Forest Res. 2011, 130, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Castedo-Dorado, F.; Gómez-Vázquez, I.; Fernandes, P.M.; Crecente-Campo, F. Shrub fuel characteristics estimated from overstory variables in NW Spain pine stands. For. Ecol. Manag. 2012, 275, 130–141. [Google Scholar] [CrossRef]
- Nunes, L.; Álvarez-González, J.; Alberdi, I.; Silva, V.; Rocha, M.; Rego, F.C. Analysis of the occurrence of wildfires in the Iberian Peninsula based on harmonised data from national forest inventories. Ann. Forest Sci. 2019, 76. [Google Scholar] [CrossRef] [Green Version]
- Pasta, S.; Mantia, T.L.; Rühl, J. The impact of Pinus halepensis afforestation on Mediterranean spontaneous vegetation: Do soil treatment and canopy cover matter? J. For. Res. 2012, 23, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, M.N.; Spotswood, E.N.; Cañadas, E.M.; Navarro, F.B. Stand management to reduce fire risk promotes understorey plant diversity and biomass in a semi-arid Pinus halepensis plantation. Appl. Veg. Sci. 2015, 18, 467–480. [Google Scholar] [CrossRef]
- Tartarino, P.; Galante, W.; Greco, R. Using the Hart-Becking spacing index in a study of the naturalisation of Pinus halepensis Miller plantation stands in the South-Eastern Salento peninsula [Southern Italy]. In Proceedings of the International Workshop MEDPINE 3: Conservation, Regeneration and Restoration of Mediterranean Pines and Their Ecosystems, Bari, Italy, 26–30 September 2005; pp. 175–188. [Google Scholar]
- Di Rita, F.; Magri, D. Holocene drought, deforestation and evergreen vegetation development in the central Mediterranean: A 5500 year record from Lago Alimini Piccolo, Apulia, southeast Italy. Holocene 2009, 19, 295–306. [Google Scholar] [CrossRef]
- Lorenzoni, G.; Ghirelli, L. Lineamenti della vegetazione del Salento (Puglia meridionale-Italia). Thalass. Salentina 1988, 18, 11–19. [Google Scholar]
- Nathan, R.; Safriel, U.N.; Noy-Meir, I.; Schiller, G. Seed release without fire inPinus halepensis, a Mediterranean serotinous wind-dispersed tree. J. Ecol. 1999, 87, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Magini, E. Ricerche sui fattori della rinnovazione naturale dell‘abete bianco sull’Appennino. L Ital. For. E Mont. 1967, 22, 261–270. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference. A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 20. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Legendre, P.; Oksanen, J.; Ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5-5. 2019. Available online: https://r.meteo.uni.wroc.pl/web/packages/vegan/index.html (accessed on 10 June 2020).
- R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019.
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Springer: New York, NY, USA, 2007; p. 680. [Google Scholar]
- Alrababah, M.A.; Tadros, M.J.; Samarah, N.H.; Ghosheh, H. Allelopathic effects of Pinus halepensis and Quercus coccifera on the germination of Mediterranean crop seeds. New For. 2009, 38, 261–272. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J.; Bautista, S. Mechanisms underlying the interaction between Pinus halepensis and the native late-successional shrub Pistacia lentiscus in a semi-arid plantation. Ecography 2004, 27, 776–786. [Google Scholar] [CrossRef]
- Knowles, R.L.; Horvath, G.C.; Carter, M.A.; Hawke, M.F. Developing a canopy closure model to predict overstorey/understorey relationships in Pinus radiata silvopastoral systems. In Agroforestry for Sustainable Land-Use Fundamental Research and Modelling with Emphasis on Temperate and Mediterranean Applications, Proceedings of the workshop held in Montpellier, Montpellier, France, 23–29 June 1997; Auclair, D., Dupraz, C., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1999; pp. 109–119. [Google Scholar]
- Bergqvist, G. Wood volume yield and stand structure in Norway spruce understorey depending on birch shelterwood density. For. Ecol. Manag. 1999, 122, 221–229. [Google Scholar] [CrossRef]
- Harrington, T.B. Overstory and understory relationships in longleaf pine plantations 14 years after thinning and woody control. Can. J. Forest Res. 2011, 41, 2301–2314. [Google Scholar] [CrossRef]
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Navarro, F.B.; Jiménez, M.N.; Gallego, E.; Ripoll, M.A. Short-term effects of overstory reduction and slash mulching on ground vegetation in a Mediterranean Aleppo pine woodland. Eur. J. Forest Res. 2010, 129, 689–696. [Google Scholar] [CrossRef]
- De las Heras, J.; Gonzalez-Ochoa, A.; Lopez-Serrano, F.; Simarro, M.E. Effects of silviculture treatments on vegetation after fire in Pinus halepensis Mill. woodlands (SE Spain). Ann. Forest Sci. 2004, 61, 661–667. [Google Scholar] [CrossRef]
- De Stefano, A.; Blazier, M.A.; Comer, C.E.; Dean, T.J.; Wigley, T.B. Understory vegetation richness and diversity of Eucalyptus benthamii and Pinus elliottii plantations in the Midsouth US. For. Sci. 2019. [Google Scholar] [CrossRef]
- Duan, W.; Ren, H.; Fu, S.; Wang, J.; Zhang, J.; Yang, L.; Huang, C. Community comparison and determinant analysis of understory vegetation in six plantations in South China. Restor. Ecol. 2010, 18, 206–214. [Google Scholar] [CrossRef]
- Otto, R.; García-Del-Rey, E.; Méndez, J.; Fernández-Palacios, J.M. Effects of thinning on seed rain, regeneration and understory vegetation in a Pinus canariensis plantation (Tenerife, Canary Islands). For. Ecol. Manag. 2012, 280, 71–81. [Google Scholar] [CrossRef]
- Pausas, J.G. Species richness patterns in the understorey of Pyrenean Pinus sylvestris forest. J. Veg. Sci. 1994, 5, 517–524. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Jack, S.B.; Engstrom, R.T. Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. Forest Res. 2006, 36, 2724–2736. [Google Scholar] [CrossRef] [Green Version]
- Kirkman, L.K.; Goebel, P.C.; Palik, B.J.; West, L.T. Predicting plant species diversity in a longleaf pine landscape. Ecoscience 2004, 11, 80–93. [Google Scholar] [CrossRef]
- Amici, V.; Santi, E.; Filibeck, G.; Diekmann, M.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Influence of secondary forest succession on plant diversity patterns in a Mediterranean landscape. J. Biogeogr. 2013, 40, 2335–2347. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. Forest Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Bonari, G.; Acosta, A.T.R.; Angiolini, C. Mediterranean coastal pine forest stands: Understorey distinctiveness or not? For. Ecol. Manag. 2017, 391, 19–28. [Google Scholar] [CrossRef]
- Wilson, F. Numerical expression of stocking in terms of height. J. For. 1946, 44, 758–761. [Google Scholar]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Perino, A.; Pereira, H.M.; Navarro, L.M.; Fernández, N.; Bullock, J.M.; Ceaușu, S.; Cortés-Avizanda, A.; Van Klink, R.; Kuemmerle, T.; Lomba, A.; et al. Rewilding complex ecosystems. Science 2019, 364, eaav5570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Aparicio, L.; Zavala, M.A.; Bonet, F.J.; Zamora, R. Are pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecol. Appl. 2009, 19, 2124–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Abbreviation | Min. | Max. | Mean ± SD |
|---|---|---|---|---|
| Overstory | ||||
| Tree density (n ha−1) * | Tree Density | 297 | 594 | 432.8 ± 74.2 |
| Average diameter at breast height (cm) | DBH | 26.6 | 33.9 | 29.8 ± 1.8 |
| Average tree height (m) | Tree Height | 12.2 | 17.7 | 15.1 ± 1.3 |
| Dominant height (m) | Hdom | 14.1 | 18.9 | 16.8 ± 1.2 |
| Average base height green canopy (m) | Canopy Green Height | 5.7 | 10.1 | 8.2 ± 0.9 |
| Average base height full canopy (m) | Canopy Base Height | 2.3 | 6.5 | 3.9 ± 1.4 |
| Average length of full canopy (m) | Canopy Length | 5.4 | 8.6 | 7.0 ± 0.8 |
| Basal area (m2 ha−1) | Basal Area | 23.0 | 37.8 | 30.0 ± 4.2 |
| Canopy cover (%) | Canopy Cover | 82.5 | 97.1 | 90.9 ± 36.0 |
| Average taper ratio | Taper Ratio | 42.0 | 61.5 | 50.7 ± 4.3 |
| Average percent. green canopy | Perc. Green Canopy | 19.3 | 55.7 | 44.3 ± 5.9 |
| Average tree distance (m) * | Tree Distance | 4.1 | 5.8 | 4.9 ± 0.4 |
| Hart-Becking index (%) | Hart-Becking | 22.5 | 36.5 | 29.2 ± 3.5 |
| Understory | ||||
| Number of species | Richness | 1 | 12 | 7.2 ± 2.9 |
| Plant density (plants m−2) | Density | 1.8 | 45.3 | 22.6 ± 11.0 |
| Average plant height (m) | Height | 0.1 | 1.3 | 0.5 ± 0.2 |
| Magini index | Magini | 0.4 | 28.2 | 11.8 ± 8.0 |
| Shannon-Wiener index | Shannon | 0.07 | 1.66 | 1.06 ± 0.39 |
| Species | Symbol | Abs. freq. | Rel. freq. | Height (m) | Density (n m−2) | Magini (m m−2) |
|---|---|---|---|---|---|---|
| Rubia peregrina L. * | Rp | 33 | 1.00 | 0.36 | 7.68 | 2.76 |
| Smilax aspera L. * | Sa | 30 | 0.91 | 0.91 | 1.39 | 1.26 |
| Phillyrea latifolia L. | Pla | 28 | 0.85 | 0.55 | 1.94 | 1.07 |
| Pistacia lentiscus L. | Ple | 28 | 0.85 | 0.63 | 0.53 | 0.33 |
| Olea europaea L. var. sylvestris | Oe | 28 | 0.85 | 0.46 | 0.61 | 0.28 |
| Rhamnus alaternus L. | Ral | 27 | 0.82 | 0.75 | 2.60 | 1.95 |
| Asparagus acutifolius L. * | Aa | 27 | 0.82 | 0.58 | 0.83 | 0.48 |
| Quercus coccifera L. | Qc | 25 | 0.76 | 0.47 | 5.69 | 2.67 |
| Myrtus communis L. | Mc | 21 | 0.64 | 0.62 | 1.35 | 0.84 |
| Rosa sempervirens L. | Rs | 20 | 0.61 | 0.49 | 1.73 | 0.85 |
| Ruscus aculeatus L. | Rac | 19 | 0.58 | 0.35 | 1.54 | 0.54 |
| Lonicera implexa Aiton * | Li | 17 | 0.52 | 0.68 | 0.72 | 0.49 |
| Rubus spp. | Rsp | 15 | 0.45 | 0.77 | 0.65 | 0.50 |
| Arbutus unedo L. | Au | 11 | 0.33 | 0.22 | 0.30 | 0.07 |
| Osyris alba L. | Oa | 9 | 0.27 | 0.57 | 0.59 | 0.34 |
| Clematis flammula L. * | Cf | 6 | 0.18 | 0.82 | 0.22 | 0.18 |
| Crataegus monogyna Jacq. | Cm | 3 | 0.09 | 0.28 | 0.15 | 0.04 |
| Viburnum tinus L. | Vt | 3 | 0.09 | 0.53 | 0.79 | 0.42 |
| Coronilla emerus L. | Ce | 2 | 0.06 | 0.55 | 0.15 | 0.08 |
| Quercus ilex L. | Qi | 2 | 0.06 | 1.75 | 0.22 | 0.39 |
| Prunus spinosa L. | Ps | 2 | 0.06 | 0.54 | 0.44 | 0.24 |
| Prasium majus L. | Pm | 2 | 0.06 | 0.61 | 0.74 | 0.45 |
| Laurus nobilis L. | Ln | 1 | 0.03 | 0.32 | 0.74 | 0.24 |
| Pittosporum tobira L. (Thunb.) Ait. | Pt | 1 | 0.03 | 0.52 | 0.15 | 0.08 |
| Models | Variables | Coeff. | SE | t-value | AICc | R2 fix. | R2 tot. |
|---|---|---|---|---|---|---|---|
| Density | |||||||
| Dh | Hart–Becking | 1.5979 | 0.4210 | 3.7950 *** | 696.5 | 0.25 | 0.73 |
| Db | Basal Area | −1.4194 | 0.3415 | −4.1561 *** | 694.9 | 0.28 | 0.73 |
| Dc | Can. Cover Can. Base Heig. | −0.0188 1.8134 | 0.0034 0.9065 | −5.4708 *** 2.0003 ns | 694.4 | 0.42 | 0.73 |
| Height | |||||||
| Hh | Hart–Becking | 0.0407 | 0.0082 | 4.9633 *** | −62.0 | 0.36 | 0.75 |
| Hb | Basal Area | −0.0316 | 0.0072 | −4.3687 *** | −58.4 | 0.31 | 0.75 |
| Hc | Can. Cover | −0.0005 | 0.0001 | −8.7896 *** | −73.4 | 0.59 | 0.75 |
| Magini | |||||||
| Mh | Hart–Becking | 1.4262 | 0.2840 | 5.0225 *** | 610.4 | 0.37 | 0.80 |
| Mb | Basal Area | −1.2835 | 0.2217 | −5.7884 *** | 606.5 | 0.44 | 0.80 |
| Mc | Can. Cover Can. Base Heig. | −0.0179 0.9810 | 0.0017 0.4592 | −10.2877 *** 2.1362 *** | 592.0 | 0.66 | 0.80 |
| Richness | |||||||
| Rh | Hart–Becking | 0.3826 | 0.1191 | 3.2121 *** | 439.2 | 0.20 | 0.75 |
| Rb | Basal Area | −0.3004 | 0.1016 | −2.9570 *** | 440.8 | 0.18 | 0.75 |
| Rc | Can. Cover | −0.0048 | 0.0010 | −4.6366 *** | 441.1 | 0.33 | 0.75 |
| Shannon | |||||||
| Sh | Hart–Becking | 0.0544 | 0.0156 | 3.4818 *** | 47.9 | 0.22 | 0.75 |
| Sb | Basal Area | −0.0395 | 0.0137 | −2.8932 *** | 51.1 | 0.17 | 0.75 |
| Sc | Can. Cover | −0.0006 | 0.0001 | −4.7772 *** | 50.3 | 0.34 | 0.75 |
| Function | Variables | F | SE | AICc | BIC | R2 | adj. R2 |
|---|---|---|---|---|---|---|---|
| Magini | |||||||
| P_Mh (2) | Hart–Becking | 17.2687 *** | 5.296 | 209.95 | 214.51 | 0.53 | 0.50 |
| P_Mb (1) | Basal Area | 33.7739 *** | 5.287 | 208.31 | 211.97 | 0.52 | 0.51 |
| P_Mc (3) | Can. Cover | 43.9490 *** | 3.355 | 181.49 | 186.75 | 0.82 | 0.80 |
| Shannon | |||||||
| P_Sh (2) | Hart–Becking | 9.4909 *** | 0.292 | 18.84 | 23.40 | 0.39 | 0.35 |
| P_Sb (1) | Basal Area | 8.4057 *** | 0.326 | 24.50 | 28.17 | 0.21 | 0.19 |
| P_Sc (3) | Can. Cover | 13.5110 *** | 0.245 | 8.96 | 14.22 | 0.58 | 0.54 |
| Studies | Stand Age | Soil | Climate | Precip. mm | Country: Region(s) | Study Type |
|---|---|---|---|---|---|---|
| Coll et al. [27] | Nonspecified | Non-specified | Nonspecified | 500–750 | Spain: Catalonia | Data from NFI |
| Jiménez et al. [31] | 10–15 years | Petric calcisol | Semiarid | 302 | Spain: Granada | Thinning experiment |
| De las Heras et al. [54] | 5 years | Sandy, sandy loam | Arid, semiarid | 226, 845 | Spain: Albacete, Murcia | Thinning experiment |
| Pasta et al. [30] | 15–46 years | From limestone, marls | Infra-Mediterranean upper semiarid | 300 | Italy: Lampedusa | Field survey |
| Nunes et al. [2] | Nonspecified | From limestone | Dry subhumid | 650 | Portugal: Setúbal | Thinning experiment |
| Navarro et al. [53] | 11 years | Petric calcisol | Semiarid | 302 | Spain: Granada | Thinning experiment |
| Overstory Variables | Understory Variables | ||||
|---|---|---|---|---|---|
| Cover | Density | Biomass | Richness | Shannon | |
| Stocking | ns [27] negative [31] | negative [31] | negative [30,31,54] | negative [31] | |
| Density | ns [54] | negative [2] | negative [53] | negative [54] ns [2] | ns [2,54] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tartarino, P.; Greco, R.; Silva, J.S. Overstory Effects on the Understory of Aleppo Pine Plantations—Implications for Ecosystem Restoration. Forests 2020, 11, 664. https://doi.org/10.3390/f11060664
Tartarino P, Greco R, Silva JS. Overstory Effects on the Understory of Aleppo Pine Plantations—Implications for Ecosystem Restoration. Forests. 2020; 11(6):664. https://doi.org/10.3390/f11060664
Chicago/Turabian StyleTartarino, Patrizia, Roberto Greco, and Joaquim S. Silva. 2020. "Overstory Effects on the Understory of Aleppo Pine Plantations—Implications for Ecosystem Restoration" Forests 11, no. 6: 664. https://doi.org/10.3390/f11060664
APA StyleTartarino, P., Greco, R., & Silva, J. S. (2020). Overstory Effects on the Understory of Aleppo Pine Plantations—Implications for Ecosystem Restoration. Forests, 11(6), 664. https://doi.org/10.3390/f11060664