Landscape Patterns of Rare Vascular Plants in the Lower Athabasca Region of Alberta, Canada



Abstract
:1. Introduction
2. Materials and Methods
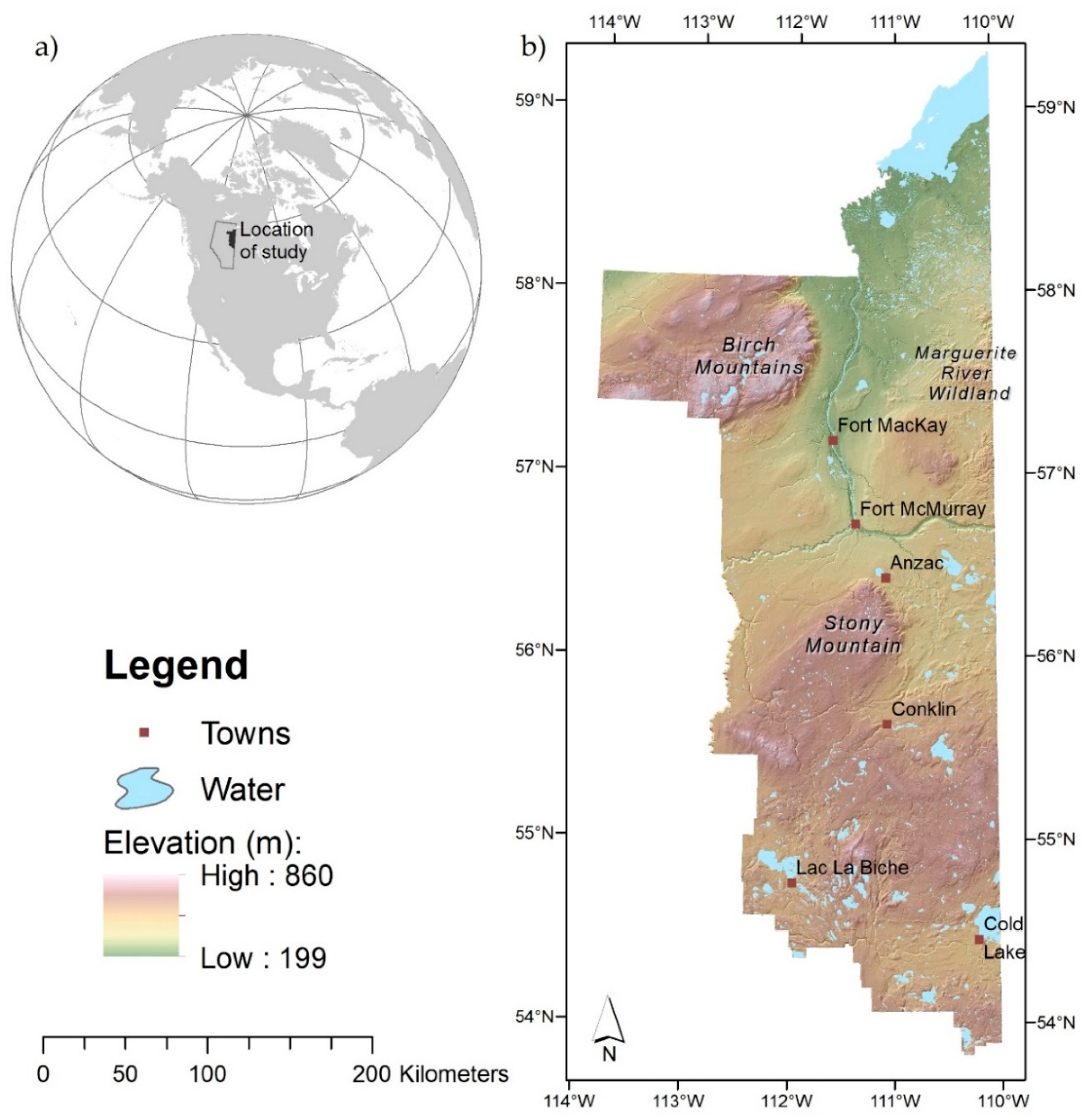
2.1. Background to Initial Model Building, Field Methods, and Data Sources
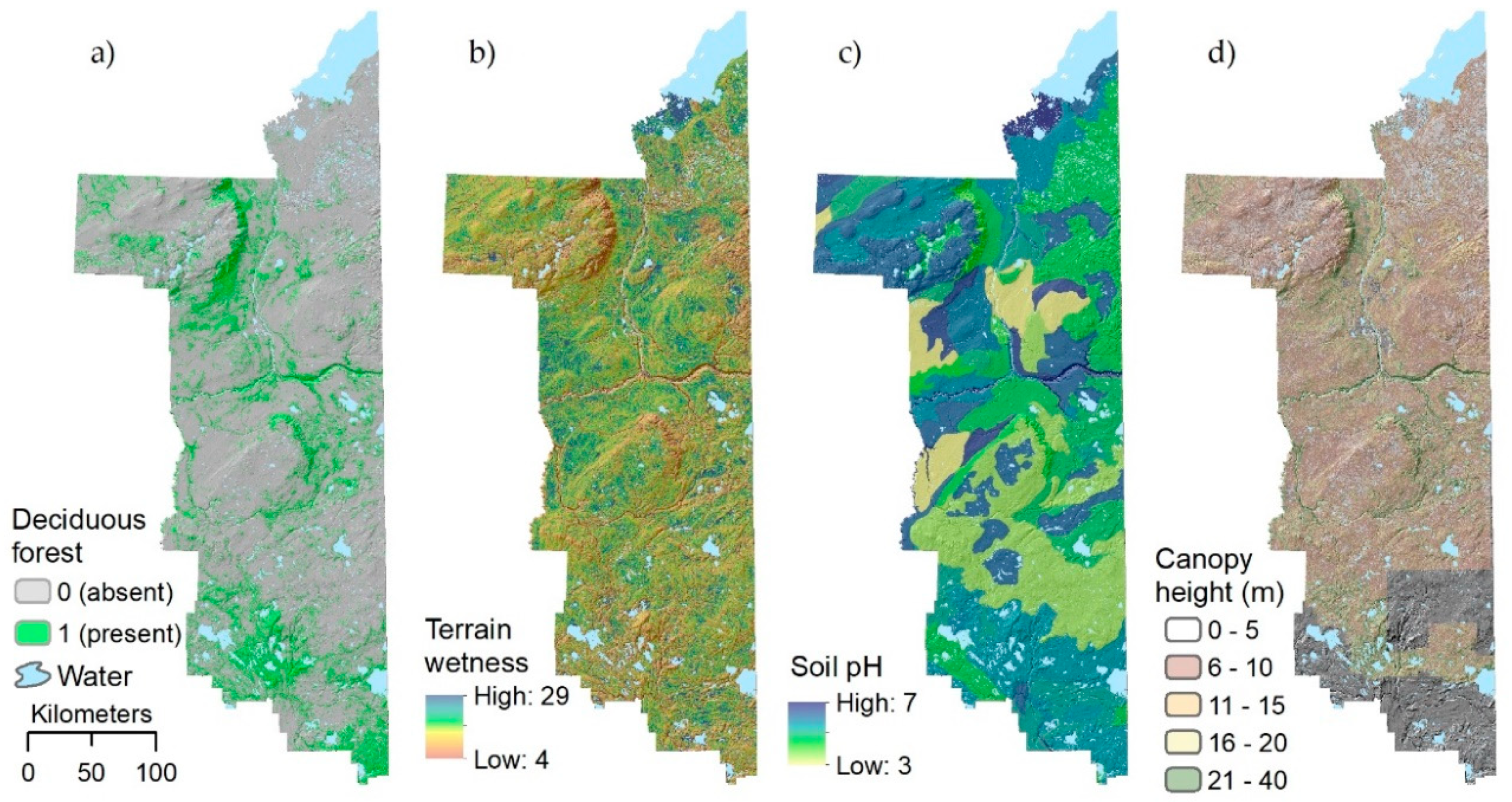
2.2. Final Landscape Map of Rare Plant Habitat
3. Results
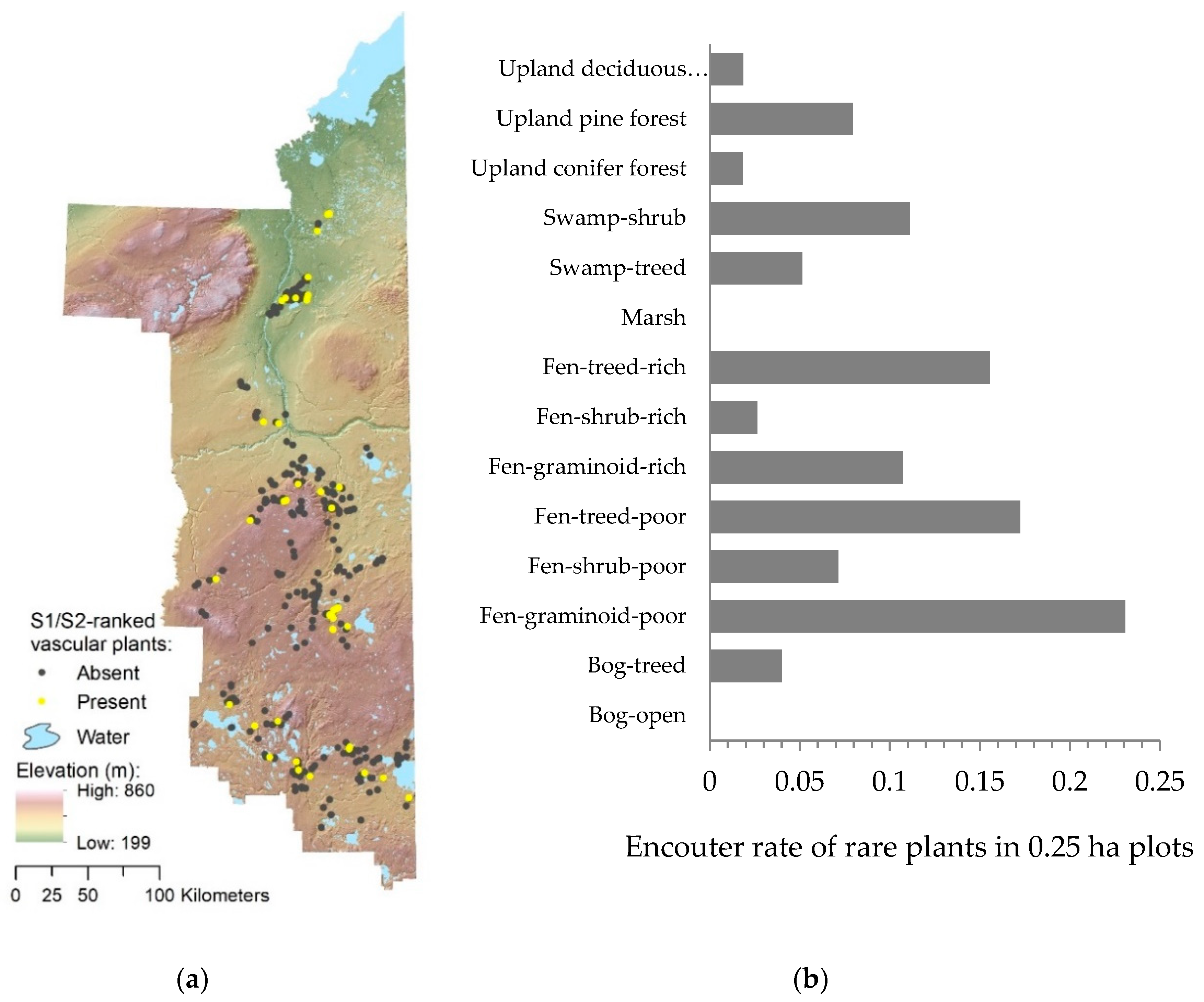
3.1. General Patterns in Plant Rarity
3.2. Landscape Predictors of Plant Rarity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sub-National Conservation Status Rank (S-Rank) | Number of Observations |
|---|---|---|
| Carex adusta | S1 | 7 |
| Carex hystericina | S1 | 1 |
| Lechea intermedia var. depauperata | S1 | 1 |
| Malaxis paludosa | S1 | 9 |
| Spiranthes lacera | S1 | 3 |
| Utricularia cornuta | S1 | 1 |
| Botrychium simplex | S2 | 1 |
| Carex heleonastes | S2 | 8 |
| Carex lacustris | S2 | 2 |
| Carex umbellata | S2 | 2 |
| Diphasiastrum sitchense | S2 | 5 |
| Hypericum majus | S2 | 1 |
| Juncus brevicaudatus | S2 | 5 |
| Juncus stygius | S2 | 6 |
| Lactuca biennis | S2 | 2 |
References
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will Remote Sensing Shape the Next Generation of Species Distribution Models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Edwards, T.C.; Cutler, D.R.; Zimmermann, N.E.; Geiser, L.; Alegria, J. Model-Based Stratifications for Enhancing the Detection of Rare Ecological Events. Spec. Featur. Ecol. 2005, 86, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Broennimann, O.; Engler, R.; Vust, M.; Yoccoz, N.G.; Lehmann, A.; Zimmermann, N.E. Using Niche-Based Models to Improve the Sampling of Rare Species. Conserv. Biol. 2006, 20, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Bourg, N.A.; Mcshea, W.J.; Gill, D.E. Putting a Cart before the Search: Successful Habitat Prediction for a Rare Forest Herb. Ecology 2005, 86, 2793–2804. [Google Scholar] [CrossRef]
- Williams, J.N.; Seo, C.; Thorne, J.; Nelson, J.K.; Erwin, S.; O’Brien, J.M.; Schwartz, M.W. Using Species Distribution Models to Predict New Occurrences for Rare Plants. Divers. Distrib. 2009, 15, 565–576. [Google Scholar] [CrossRef]
- De Queiroz, T.F.; Baughman, C.; Baughman, O.; Gara, M.; Williams, N. Species Distribution Modeling for Conservation of Rare, Edaphic Endemic Plants in White River Valley, Nevada. Nat. Areas J. 2012, 32, 149. [Google Scholar] [CrossRef]
- Peterman, W.E.; Crawford, J.A.; Kuhns, A.R. Using Species Distribution and Occupancy Modeling to Guide Survey Efforts and Assess Species Status. J. Nat. Conserv. 2013, 21, 114–121. [Google Scholar] [CrossRef]
- Rabinowitz, D. Seven Forms of Rarity. In The Biological Aspects of Rare Plant Conservation; Synge, H., Ed.; John Wiley & Sons Ltd.: Somerset, NJ, USA, 1981; pp. 205–217. [Google Scholar]
- Wang, O.; Zachmann, L.J.; Sesnie, S.E.; Olsson, A.D.; Dickson, B.G. An Iterative and Ttargeted Sampling Design informed by Habitat Suitability Models for Detecting Focal Plant Species over Extensive Areas. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Lefsky, M.; Cohen, W.; Parker, G.; Harding, D. Lidar Remote Sensing for Ecosystem Studies. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Beland, M.; Parker, G.; Sparrow, B.; Harding, D.; Chasmer, L.; Phinn, S.; Antonarakis, A.; Strahler, A. On Promoting the Use of Lidar Systems in Forest Ecosystem Research. For. Ecol. Manag. 2019, 450, 117484. [Google Scholar] [CrossRef]
- Sellars, J.D.; Jolls, C.L. Habitat Modeling for Amaranthus Pumilus: An Application of Light Detection and Ranging (LIDAR) Data. J. Coast. Res. 2007, 23, 1193–1202. [Google Scholar] [CrossRef]
- Questad, E.J.; Kellner, J.R.; Kinney, K.; Cordell, S.; Asner, G.P.; Thaxton, J.; Diep, J.; Uowolo, A.; Brooks, S.; Inman-Narahari, N.; et al. Mapping Habitat Suitability for At-Risk Plant Species and its Implications for Restoration and Reintroduction. Ecol. Appl. 2014, 24, 385–395. [Google Scholar] [CrossRef] [PubMed]
- West, A.M.; Evangelista, P.H.; Jarnevich, C.S.; Kumar, S.; Swallow, A.; Luizza, M.W.; Chignell, S.M. Using Multi-Date Satellite Imagery to Monitor Invasive Grass Species Distribution in Post-Wildfire Landscapes: An Iterative, Adaptable Approach that Employs Open-Source Data and Software. Int. J. Appl. Earth Obs. Geoinf. 2017, 59, 135–146. [Google Scholar] [CrossRef]
- Burgman, M.A.; Keith, D.; Hopper, S.D.; Widyatmoko, D.; Drill, C. Threat Syndromes and Conservation of the Australian Flora. Biol. Conserv. 2007, 134, 73–82. [Google Scholar] [CrossRef]
- Rooney, R.C.; Bayley, S.E.; Schindler, D.W. Oil Sands Mining and Reclamation Cause Massive Loss of Peatland and Stored Carbon. Proc. Natl. Acad. Sci. USA 2012, 109, 4933–4937. [Google Scholar] [CrossRef] [Green Version]
- Price, J.S.; McLaren, R.G.; Rudolph, D.L. Landscape Restoration After Oil Sands Mining: Conceptual Design and Hydrological Modelling for Fen Reconstruction. Int. J. Min. Reclam. Environ. 2010, 24, 109–123. [Google Scholar] [CrossRef]
- Renault, S.; Lait, C.; Zwiazek, J.; MacKinnon, M. Effect of High Salinity Tailings Waters Produced from Gypsum Treatment of Oil Sands Tailings on Plants of the Boreal Forest. Environ. Pollut. 1998, 102, 177–184. [Google Scholar] [CrossRef]
- Purdy, B.G.; Macdonald, S.E.; Lieffers, V.J. Naturally Saline Boreal Communities as Models for Reclamation of Saline Oil Sand Tailings. Restor. Ecol. 2005, 13, 667–677. [Google Scholar] [CrossRef]
- Dabros, A.; James Hammond, H.E.; Pinzon, J.; Pinno, B.; Langor, D. Edge Influence of Low-Impact Seismic Lines for Oil Exploration on Upland Forest Vegetation in Northern Alberta (Canada). For. Ecol. Manag. 2017, 400, 278–288. [Google Scholar] [CrossRef]
- Rooney, R.C.; Bayley, S.E. Setting reclamation targets and evaluating progress: Submersed Aquatic Vegetation in Natural and Post-Oil sands Mining Wetlands in Alberta, Canada. Ecol. Eng. 2011, 37, 569–579. [Google Scholar] [CrossRef]
- Guo, X.; Coops, N.C.; Tompalski, P.; Nielsen, S.E.; Bater, C.W.; Stadt, J.J. Regional Mapping of Vegetation Structure for Biodiversity Monitoring Using Airborne Lidar Data. Ecol. Inform. 2017, 38, 50–61. [Google Scholar] [CrossRef]
- Coops, N.C.; Tompaski, P.; Nijland, W.; Rickbeil, G.J.M.; Nielsen, S.E.; Bater, C.W.; Stadt, J.J. A Forest Structure Habitat Index Based on Airborne Laser Scanning Data. Ecol. Indic. 2016, 67, 346–357. [Google Scholar] [CrossRef]
- Ducks Unlimited Canada. Enhanced Wetland Classification Inferred Products User Guide Version 1.0; Ducks Unlimited Canada: Edmonton, AB, Canada, 2011. [Google Scholar]
- Master, L.L.; Faber-Langendoen, D.; Bittman, R.; Hammerson, G.A.; Heidel, B.; Ramsay, L.; Snow, K.; Teucher, A.; Tomaino, A. NatureServe Conservation Status Assessments: Factors for Evaluating Species and Ecosystem Risk; NatureServe: Arlington, VA, USA, 2012. [Google Scholar]
- Zhang, J.; Nielsen, S.E.; Grainger, T.N.; Kohler, M.; Chipchar, T.; Farr, D.R. Sampling Plant Diversity and Rarity at Landscape Scales: Importance of Sampling Time in Species Detectability. PLoS ONE 2014, 9, e95334. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Dennett, J.; Denny, C.; Kohler, M.; Farr, D. Terrestrial Vascular Plant Monitoring Project for the Lower Athabasca (2012–2016); Applied Conservation Ecology Laboratory: Edmonton, AB, Canada, 2017. [Google Scholar]
- Alberta Biodiversity Monitoring Institute. ABMI 10-Year Science and Program Review v. 8.1; Alberta Biodiversity Monitoring Institute: Edmonton, AB, Canada, 2019. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2000. [Google Scholar]
- StataCorp. Stata Statistical Software: Release 14; StataCorp: College Station, TX, USA, 2015. [Google Scholar]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Natural Regions Committee. Natural Regions and Subregions of Alberta; Pub. No. T/852; Downing, D.J., Pettapiece, W.W., Eds.; Government of Alberta: Edmonton, AB, Canada, 2006; ISBN 0778545725.
- Natural Resources Canada. CanTopo Digital Elevation Model; Natural Resources Canada: Ottawa, ON, Canada, 2014. [Google Scholar]
- Moore, I.; Gessler, P.; Nielsen, G.; Petersen, G. Terrain Attributes: Estimation Methods and Scale Effects. In Modeling Change in Environmental Systems; Jakeman, A., Beck, M., McAleer, M., Eds.; Wiley: London, UK, 1993; pp. 189–214. [Google Scholar]
- Gessler, P.; Moore, I.; McKenzie, N.; Ryan, P. Soil-Landscape Modelling and Spatial Prediction of Soil Attributes. Int. J. Geogr. Inf. Syst. 1995, 9, 421–432. [Google Scholar] [CrossRef]
- White, B.; Ogilvie, J.; Campbell, D.M.H.; Hiltz, D.; Gautheir, B.; Chisholm, H.K.H.; Wen, H.K.; Murphy, P.N.C.; Arp, P.A. Using the Cartographic Depth-to-Water Index to Locate Small Streams and Associated Wet Areas across Landscapes. Can. Water Resour. J. 2012, 37, 333–347. [Google Scholar] [CrossRef]
- Soil Landscapes of Canada Working Group. Available online: https://open.canada.ca/data/en/dataset/5ad5e20c-f2bb-497d-a2a2-440eec6e10cd (accessed on 1 January 2014).
- McGaughey, R. FUSION/LVD: Software for LIDAR Data Analysis and Visualization; US Forest Service: Washington, DC, USA, 2016.
- Parker, G.G.; Russ, M.E. The Canopy Surface and Stand Development: Assessing Forest Canopy Structure and Complexity with Near-Surface Altimetry. For. Ecol. Manage. 2004, 189, 307–315. [Google Scholar] [CrossRef]
- Mura, M.; McRoberts, R.E.; Chirici, G.; Marchetti, M. Estimating and Mapping Forest Structural Diversity Using Airborne Laser Scanning Data. Remote Sens. Environ. 2015, 170, 133–142. [Google Scholar] [CrossRef]
- Møller, A.; Jennions, M. How Much Variance can be explained by Ecologists and Evolutionary Biologists? Oecologia 2002, 132, 492–500. [Google Scholar] [CrossRef]
- Swets, J. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Manel, S.; Williams, H.; Ormerod, S. Evaluating Presence-Absence Models in Diversity on the Roof of the World: Spatial Patterns and Environmental Determinants. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Nijland, W.; Coops, N.C.; Macdonald, S.E.; Nielsen, S.E.; Bater, C.W.; White, B.; Ogilvie, J.; Stadt, J.J. Remote Sensing Proxies of Productivity and Moisture Predict Forest Stand Type and Recovery Rate Following Experimental Harvest. For. Ecol. Manage. 2015, 357, 239–247. [Google Scholar] [CrossRef]
- Da Silveira, H.L.F.; Galvão, L.S.; Sanches, I.D.; de Sá, I.B.; Taura, T.A. Use of MSI/Sentinel-2 and Airborne Lidar Data for Mapping Vegetation and Studying the Relationhips with Soil Attributes in the Brazilian Semi-Arid Region. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 179–190. [Google Scholar] [CrossRef]
- Wendelberger, K.S.; Maschinski, J. Assessing Microsite and Regeneration Niche Preferences through Experimental Reintroduction of the Rare Plant Tephrosia Angustissima Var. Corallicola. Plant Ecol. 2016, 217, 155–167. [Google Scholar] [CrossRef]
- Crisfield, V.E.; Dennett, J.M.; Denny, C.K.; Mao, L.; Nielsen, S.E. Species Richness is a Surrogate for Rare Plant Occurrence, but not Conservation Value, in Boreal Plant Communities. Biodivers. Conserv. 2020, 29, 99–114. [Google Scholar] [CrossRef]




| Class Name | n (% of Area) | Type of Community | Soil Moisture | Hydro-Dynamics | Nutrient Regime |
|---|---|---|---|---|---|
| Emergent Marsh | 8 (0.6%) | Mineral Wetland | Very Hydric | Very Dynamic | Very Rich |
| Meadow Marsh | 1 (0.5%) | Mineral Wetland | Hydric | Very Dynamic | Very Rich |
| Graminoid Rich Fen | 28 (0.5%) | Peat Wetland | Hydric | Moving | Rich |
| Graminoid Poor Fen | 13 (0.9%) | Peat Wetland | Hydric | Slow Moving | Poor |
| Shrubby Rich Fen | 39 (3.8%) | Peat Wetland | Sub Hydric | Moving | Rich |
| Shrubby Poor Fen | 28 (0.8%) | Peat Wetland | Sub Hydric | Slow Moving | Poor |
| Treed Rich Fen | 44 (7.7%) | Peat Wetland | Sub Hydric | Moving | Rich |
| Treed Poor Fen | 58 (9.1%) | Peat Wetland | Hygric | Slow Moving | Poor |
| Open Bog | 1 (0.02%) | Peat Wetland | Sub Hygric | Stagnant | Very Poor |
| Shrubby Bog | 7 (0.7%) | Peat Wetland | Sub Hygric | Stagnant | Very Poor |
| Treed Bog | 25 (6.2%) | Peat Wetland | Sub Hygric | Stagnant | Very Poor |
| Shrub Swamp | 18 (2.5%) | Mineral Wetland | Hydric | Dynamic | Rich |
| Hardwood Swamp | 3 (0.9%) | Mineral Wetland | Hygric | Dynamic | Rich |
| Mixedwood Swamp | 3 (0.7%) | Mineral Wetland | Hygric | Dynamic | Rich |
| Tamarack Swamp | 5 (0.6%) | Mineral Wetland | Hygric | Slow Moving | Medium |
| Conifer Swamp | 30 (5.0%) | Mineral Wetland | Sub Hygric | Stagnant | Medium |
| Upland Conifer | 54 (8.7%) | Upland | Mesic to Xeric | Upland | Upland |
| Upland Deciduous | 109 (18.4%) | Upland | Mesic to Xeric | Upland | Upland |
| Upland Mixedwood | 3 (0.4%) | Upland | Mesic to Xeric | Upland | Upland |
| Upland Pine | 113 (13.0%) | Upland | Xeric | Upland | Upland |
| Burn | 4 (5.6%) | Other | Other | Other | Other |
| Model | AIC | K | ROC | R2 |
|---|---|---|---|---|
| Single factor models: | ||||
| S-Soils | 270.48 | 3 | 0.600 | 0.016 |
| T-Terrain wetness | 266.79 | 3 | 0.594 | 0.029 |
| L-Land cover | 254.54 | 11 | 0.769 | 0.134 |
| V-Vegetation structure | 250.60 | 4 | 0.765 | 0.097 |
| Null (constant) | 270.65 | 1 | 0.500 | 0.000 |
| Two-factor models: | ||||
| S + T | 267.55 | 5 | 0.638 | 0.041 |
| S + L | 251.07 | 13 | 0.804 | 0.162 |
| S + V | 249.6 | 6 | 0.778 | 0.116 |
| T + L | 252.33 | 13 | 0.788 | 0.158 |
| T + V | 248.63 | 6 | 0.777 | 0.119 |
| L + V | 247.09 | 14 | 0.829 | 0.185 |
| Three-factor models: | ||||
| S + T + L | 249.89 | 15 | 0.812 | 0.182 |
| S + T + V | 248.69 | 8 | 0.792 | 0.134 |
| S + L + V | 243.70 | 16 | 0.837 | 0.212 |
| T + L + V | 244.56 | 16 | 0.838 | 0.209 |
| Four-factor model: | ||||
| Global (S + T + L + V) | 241.45 | 17 | 0.841 | 0.228 |
| w/LiDAR Variables (n = 469) | w/o LiDAR Variables (n = 602) | |||||
|---|---|---|---|---|---|---|
| Variable | β | SE | p | β | SE | p |
| S-Soil pH | 5.706 | 2.788 | 0.041 | 6.183 | 2.686 | 0.021 |
| S-Soil pH2 | −0.581 | 0.298 | 0.051 | −0.641 | 0.287 | 0.026 |
| T-CTI (wetness) | 41.53 | 21.53 | 0.054 | 39.00 | 20.64 | 0.059 |
| T-CTI2 (wetness) | −8.679 | 4.49 | 0.053 | −8.194 | 4.31 | 0.057 |
| L-Treed bog | 3.161 | 1.453 | 0.03 | 2.556 | 1.381 | 0.064 |
| L-Graminoid poor fen | 5.324 | 1.458 | <0.001 | 5.616 | 1.353 | <0.001 |
| L-Shrub poor fen | 3.851 | 1.369 | 0.005 | 4.168 | 1.255 | 0.001 |
| L-Treed poor fen | 3.928 | 1.212 | 0.001 | 3.94 | 1.142 | 0.001 |
| L-Graminoid rich fen | 3.357 | 1.551 | 0.03 | 3.827 | 1.528 | 0.012 |
| L-Shrub rich fen | 2.426 | 1.401 | 0.083 | 2.681 | 1.329 | 0.044 |
| L-Treed rich fen | 4.263 | 1.197 | <0.001 | 4.098 | 1.126 | <0.001 |
| L-Conifer swamp | 3.174 | 1.474 | 0.031 | 3.031 | 1.466 | 0.039 |
| L-Upland burn | 4.893 | 1.708 | 0.004 | 4.544 | 1.678 | 0.007 |
| L-Upland pine | 2.520 | 1.005 | 0.012 | 2.808 | 1.024 | 0.006 |
| V-Canopy relief ratio (CRR) | 5.872 | 1.799 | 0.001 | |||
| V-Canopy height (p95) | −0.116 | 0.053 | 0.028 | |||
| Constant (intercept) | −69.29 | 26.19 | 0.008 | −65.77 | 25 | 0.009 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, S.E.; Dennett, J.M.; Bater, C.W. Landscape Patterns of Rare Vascular Plants in the Lower Athabasca Region of Alberta, Canada. Forests 2020, 11, 699. https://doi.org/10.3390/f11060699
Nielsen SE, Dennett JM, Bater CW. Landscape Patterns of Rare Vascular Plants in the Lower Athabasca Region of Alberta, Canada. Forests. 2020; 11(6):699. https://doi.org/10.3390/f11060699
Chicago/Turabian StyleNielsen, Scott E., Jacqueline M. Dennett, and Christopher W. Bater. 2020. "Landscape Patterns of Rare Vascular Plants in the Lower Athabasca Region of Alberta, Canada" Forests 11, no. 6: 699. https://doi.org/10.3390/f11060699
APA StyleNielsen, S. E., Dennett, J. M., & Bater, C. W. (2020). Landscape Patterns of Rare Vascular Plants in the Lower Athabasca Region of Alberta, Canada. Forests, 11(6), 699. https://doi.org/10.3390/f11060699