Predicting Occurrence, Abundance, and Fruiting of a Cultural Keystone Species to Inform Landscape Values and Priority Sites for Habitat Enhancements



Abstract
:1. Introduction
2. Materials and Methods
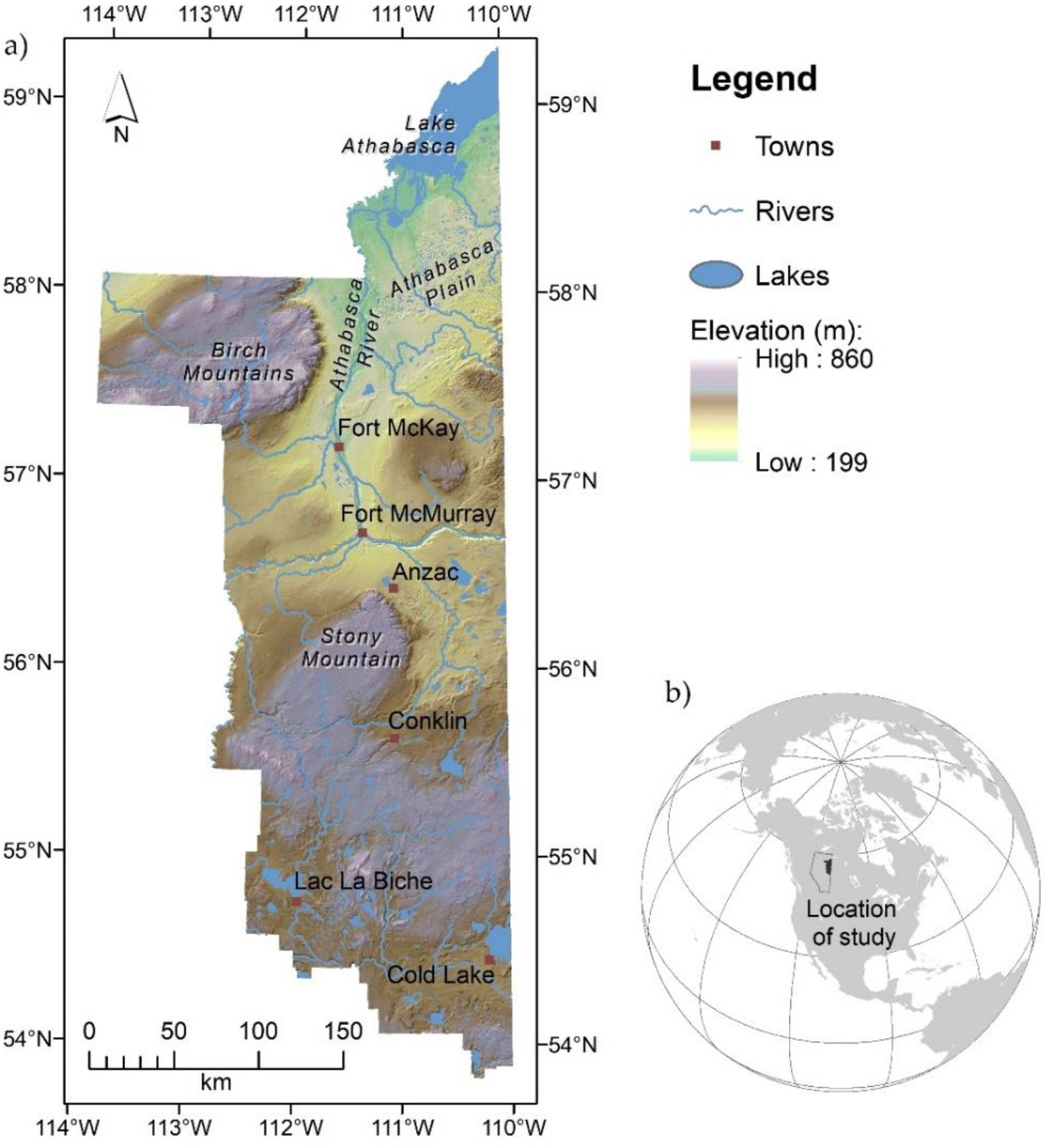
2.1. Study Species and Study Area
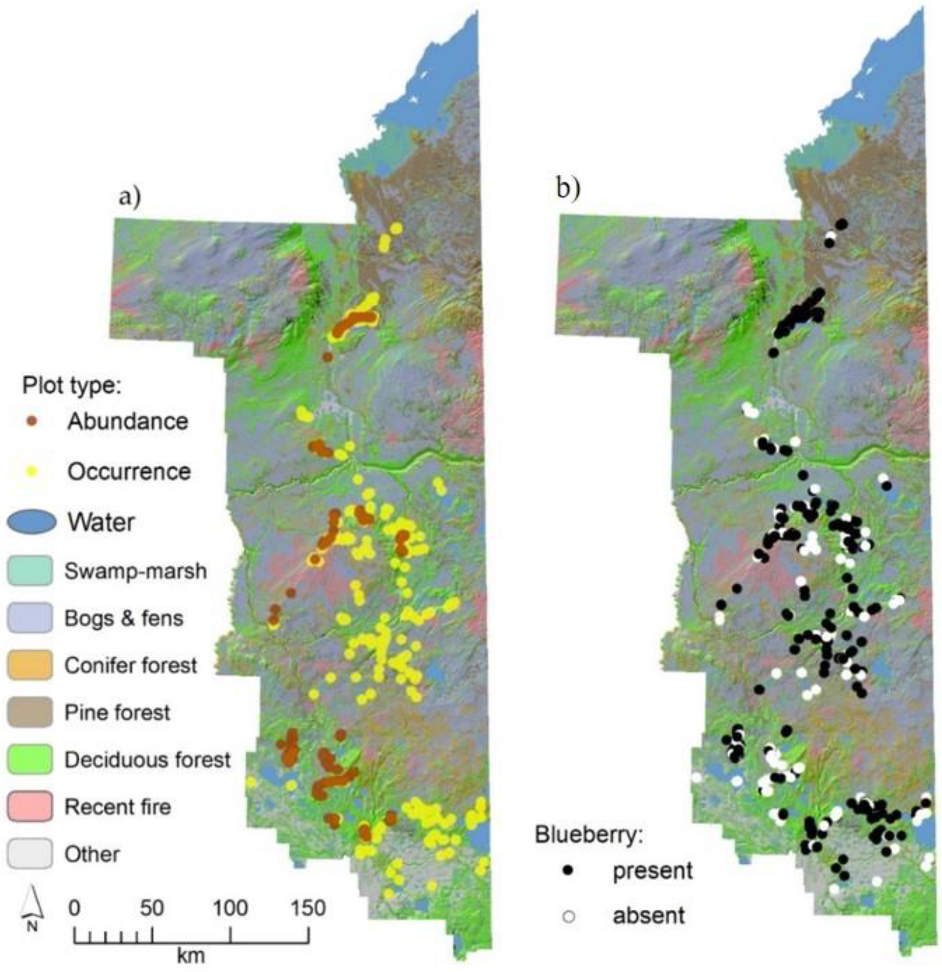
2.2. Study Design and Field Measures for Mapping Blueberry Distribution (Occurrence)
2.3. Study Design and Field Measures of Blueberry Shrub Abundance and Fruit Production
2.4. Statistical Modeling of Blueberry Occurrence, Abundance, and Fruit Production
2.5. Landscape Predictions of Blueberry Occurrence, Abundance, and Fruit Production
2.6. Habitat Enhancements: Effects of Simulated Forest Canopy Removal on Fruit Production
3. Results
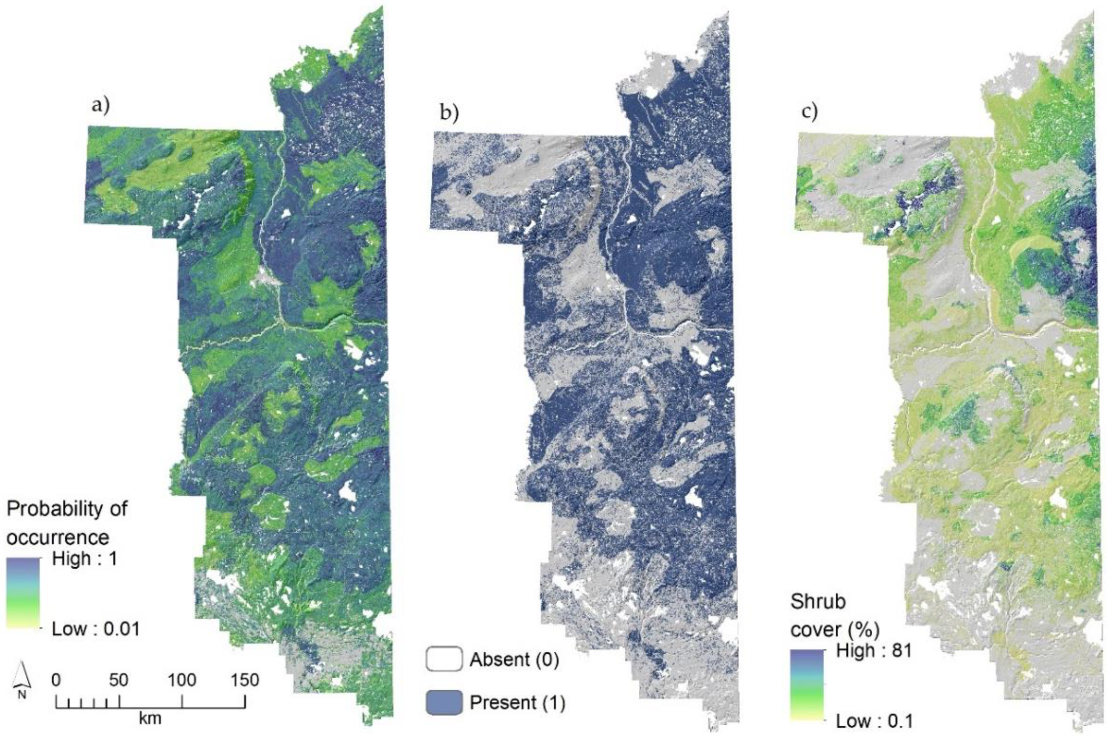
3.1. Landscape Patterns in Blueberry Occurrence
3.2. Landscape Patterns in Blueberry Shrub Abundance
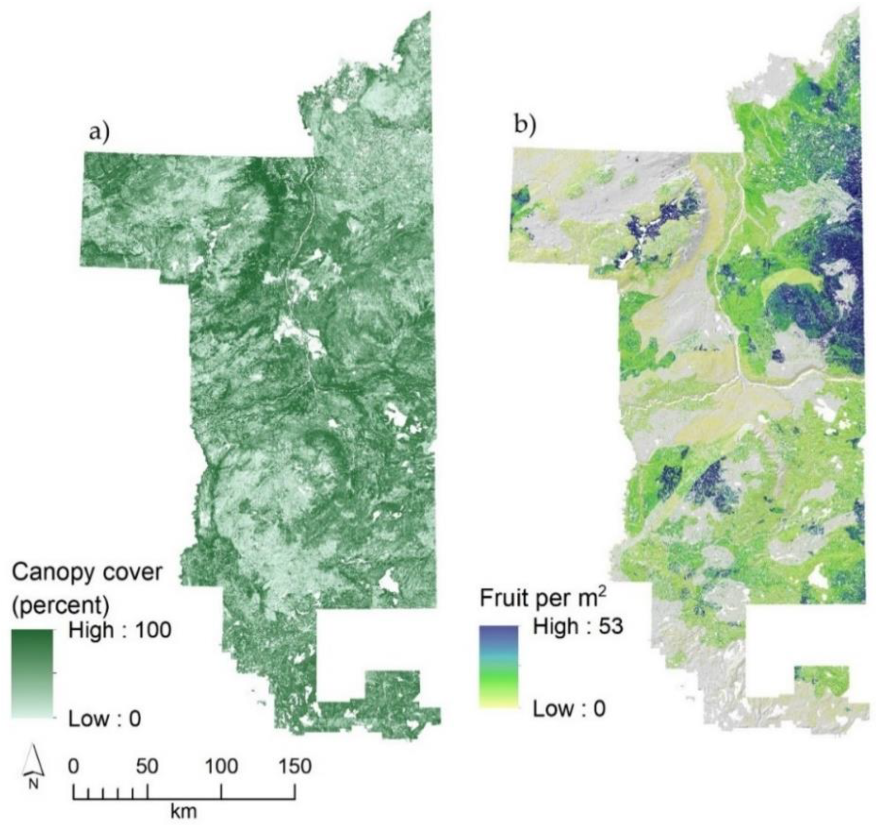
3.3. Landscape Patterns in Blueberry Fruit Production
3.4. Identifying Priority Sites for Habitat Enhancements
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Data Source |
|---|---|---|
| Land cover | (used as binary categories) | |
| Marsh | Land cover of marsh | DU-Enhanced Wetland Classification |
| Fen-G-R | Land cover of fen-graminoid-rich | DU-Enhanced Wetland Classification |
| Fen-G-P | Land cover of fen-graminoid-poor | DU-Enhanced Wetland Classification |
| Fen-S-R | Land cover of fen-shrub-rich | DU-Enhanced Wetland Classification |
| Fen-S-P | Land cover of fen-shrub-poor | DU-Enhanced Wetland Classification |
| Fen-T-P | Land cover of fen-treed-poor | DU-Enhanced Wetland Classification |
| Bog | Land cover of bog | DU-Enhanced Wetland Classification |
| Bog-treed | Land cover of bog-treed | DU-Enhanced Wetland Classification |
| Swamp-S | Land cover of swamp-shrub | DU-Enhanced Wetland Classification |
| Swamp-D | Land cover of swamp-deciduous | DU-Enhanced Wetland Classification |
| Swamp-T | Land cover of swamp-tamarack | DU-Enhanced Wetland Classification |
| Swamp-C | Land cover of swamp-conifer | DU-Enhanced Wetland Classification |
| Decid | Land cover of upland-deciduous | DU-Enhanced Wetland Classification |
| Conifer | Land cover of upland-conifer, not pine | DU-Enhanced Wetland Classification |
| Pine | Land cover of upland-pine | DU-Enhanced Wetland Classification |
| Burn | Land cover of recently burned | DU-Enhanced Wetland Classification |
| Soils | ||
| pH * | Soil pH | Soil Landscapes of Canada v3.2 |
| Clay * | Soil texture, percent clay | Soil Landscapes of Canada v3.2 |
| Sand * | Soil texture, percent sand | Soil Landscapes of Canada v3.2 |
| Depth * | Soil depth (cm) | Soil Landscapes of Canada v3.2 |
| Terrain | ||
| Slope | Percent slope | DEM, derived product (this study) |
| Wetness * | Terrain wetness (CTI method) | DEM, derived product (this study) |
| Heatload | Heatload index | DEM, derived product (this study) |
| Climate | ||
| MAT * | Mean annual temperature (°C) | ClimateAB |
| MAP * | Mean annual precipitation (mm) | ClimateAB |
| FFP | Frost free period (days) | ClimateAB |
| Forest canopy cover | ||
| Canopy | Percent forest canopy cover | Airborne Laser Scanning (this study) |
| Occurrence | Abundance | |||||
|---|---|---|---|---|---|---|
| Variable | β | SE | p | β | SE | p |
| Land cover | ||||||
| Marsh | −0.412 | 0.751 | 0.583 | |||
| Fen-G-R | −1.682 | 0.569 | 0.003 | 0.197 | 0.958 | 0.837 |
| Fen-G-P | −1.930 | 0.827 | 0.020 | 0.197 | 0.958 | 0.837 |
| Fen-S-R | −0.621 | 0.395 | 0.116 | 0.197 | 0.958 | 0.837 |
| Fen-S-P | −2.959 | 0.779 | 0.001 | 0.197 | 0.958 | 0.837 |
| Fen-T-R | −0.079 | 0.344 | 0.818 | 0.627 | 0.320 | 0.050 |
| Fen-T-P | −0.497 | 0.311 | 0.110 | −1.222 | 0.707 | 0.084 |
| Bog | −0.927 | 0.939 | 0.323 | −0.824 | 0.367 | 0.025 |
| Bog-T | −0.883 | 0.457 | 0.053 | −0.824 | 0.367 | 0.025 |
| Swamp-S | −1.040 | 0.611 | 0.089 | −0.824 | 0.367 | 0.025 |
| Swamp-D | −1.497 | 0.934 | 0.109 | −0.824 | 0.367 | 0.025 |
| Swamp-T | 14.48 | 682.1 | 0.983 | −0.824 | 0.367 | 0.025 |
| Swamp-C | 0.858 | 0.467 | 0.066 | −0.824 | 0.367 | 0.025 |
| Conifer | 0.674 | 0.267 | 0.012 | −1.329 | 0.298 | 0.001 |
| Pine | 0.866 | 0.273 | 0.002 | −0.344 | 0.272 | 0.205 |
| Burn | 1.581 | 1.071 | 0.140 | 1.962 | 0.655 | 0.003 |
| Soil | ||||||
| pH | −4.830 | 2.025 | 0.017 | |||
| pH2 | 0.460 | 0.207 | 0.027 | |||
| Clay | 0.019 | 0.008 | 0.023 | −0.194 | 0.054 | 0.001 |
| Clay2 | 0.003 | 0.001 | 0.001 | |||
| Sand | 0.019 | 0.004 | 0.001 | |||
| Depth | −0.011 | 0.004 | 0.002 | −0.076 | 0.020 | 0.001 |
| Depth2 | 0.0014 | 0.0003 | 0.001 | |||
| Terrain | ||||||
| Wetness | −0.198 | 0.054 | 0.001 | |||
| Slope | −0.389 | 0.085 | 0.001 | |||
| Heatload | −0.005 | 0.002 | 0.014 | |||
| Climate | ||||||
| MAT | −2.296 | 0.706 | 0.001 | |||
| MAP | 0.176 | 0.101 | 0.083 | |||
| MAP2 | −0.0002 | 0.0001 | 0.087 | |||
| Constant | 14.657 | 4.553 | 0.001 | −38.765 | 25.120 | 0.123 |

References
- Lefsky, M.; Cohen, W.; Parker, G.; Harding, D. Lidar remote sensing for ecosystem studies. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Franklin, J. Predictive vegetation mapping: Geographic modelling of biospatial patterns in relation to environmental gradients. Prog. Phys. Geogr. 1995, 19, 474–499. [Google Scholar] [CrossRef]
- Blanco, J.A.; Ameztegui, A.; Rodriguez, F. Modelling Forest Ecosystems: A crossroad between scales, techniques, and applications. Ecol. Model. 2020, 425, 109030. [Google Scholar] [CrossRef]
- Vauhkonen, J. Predicting the provisioning potential of forest ecosystem services using airborne laser scanning data and forest resource maps. For. Ecosyst. 2018, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Dennett, J.; Bater, C.W.; Tompalski, P.; Coops, N.C.; Farr, D.; Kohler, M.; White, B.; Stadt, J.J.; Nielsen, S.E. Using airborne laser scanning to predict plant species richness and assess conservation threats in the oil sands region of Alberta’s boreal forest. For. Ecol. Manag. 2018, 409, 29–37. [Google Scholar] [CrossRef]
- Thers, H.; Kirstine, A.; Læssøe, T.; Ejrnæs, R.; Klith, P.; Svenning, J.-C. Lidar-derived variables as a proxy for fungal species richness and composition in temperate Northern Europe. Remote Sens. Environ. 2017, 200, 102–113. [Google Scholar] [CrossRef]
- Guo, X.; Coops, N.C.; Tompalski, P.; Nielsen, S.E.; Bater, C.W.; Stadt, J.J. Regional mapping of vegetation structure for biodiversity monitoring using airborne lidar data. Ecol. Inform. 2017, 38, 50–61. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Mcdermid, G.; Stenhouse, G.B.; Boyce, M.S. Dynamic wildlife habitat models: Seasonal foods and mortality risk predict occupancy-abundance and habitat selection in grizzly bears. Biol. Conserv. 2010, 143, 1623–1634. [Google Scholar] [CrossRef]
- Pollock, S.Z.; Nielsen, S.E.; St. Clair, C.C. A railway increases the abundance and accelerates the phenology of bear-attracting plants in a forested, mountain park. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- Questad, E.J.; Kellner, J.R.; Kinney, K.; Cordell, S.; Asner, G.P.; Thaxton, J.; Diep, J.; Uowolo, A.; Brooks, S.; Inman-Narahari, N.; et al. Mapping habitat suitability for at-risk plant species and its implications for restoration and reintroduction. Ecol. Appl. 2014, 24, 385–395. [Google Scholar] [CrossRef]
- Baumflek, M.; DeGloria, S.; Kassam, K.A. Habitat modeling for health sovereignty: Increasing indigenous access to medicinal plants in northern Maine, USA. Appl. Geogr. 2015, 56, 83–94. [Google Scholar] [CrossRef]
- Gaikwad, J.; Wilson, P.D.; Ranganathan, S. Ecological niche modeling of customary medicinal plant species used by Australian Aborigines to identify species-rich and culturally valuable areas for conservation. Ecol. Model. 2011, 222, 3437–3443. [Google Scholar] [CrossRef]
- Kangas, K.; Markkanen, P. Factors affecting participation in wild berry picking by rural and urban dwellers. Silva Fenn. 2001, 35, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Kuhnlein, H.V.; Turner, N.J. Traditional Plant Foods of Canadian Indigenous Peoples; Gordon and Breach Science: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Barber, Q.E.; Bater, C.W.; Braid, A.C.R.; Coops, N.C.; Tompalski, P.; Nielsen, S.E. Airborne laser scanning for modelling understory shrub abundance and productivity. For. Ecol. Manag. 2016, 377, 46–54. [Google Scholar] [CrossRef]
- Gopalakrishnan, R.; Thomas, V.A.; Wynne, R.H.; Coulston, J.W.; Fox, T.R. Shrub detection using disparate airborne laser scanning acquisitions over varied forest cover types. Int. J. Remote Sens. 2018, 39, 1220–1242. [Google Scholar] [CrossRef]
- Reich, R.M.; Lojewski, N.; Lundquist, J.E.; Bravo, V.A. Predicting abundance and productivity of blueberry plants under insect defoliation in Alaska. J. Sustain. For. 2018, 37, 525–536. [Google Scholar] [CrossRef]
- Franklin, C.M.A.; Nielsen, S.E.; Macdonald, S.E. Understory vascular plant responses to retention harvesting with and without prescribed fire. Can. J. For. Res. 2019, 49, 1087–1100. [Google Scholar] [CrossRef]
- Koivula, M.; Vanha-Majamaa, I. Experimental evidence on biodiversity impacts of variable retention forestry, prescribed burning, and deadwood manipulation in Fennoscandia. Ecol. Process. 2020, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Granath, G.; Kouki, J.; Johnson, S.; Heikkala, O.; Rodríguez, A.; Strengbom, J. Trade-offs in berry production and biodiversity under prescribed burning and retention regimes in boreal forests. J. Appl. Ecol. 2018, 55, 1658–1667. [Google Scholar] [CrossRef] [Green Version]
- Duchesne, L.C.; Wetzel, S. Effect of Fire Intensity and Depth of Burn on Lowbush Blueberry, Vaccinium angustifolium, and Velvet Leaf Blueberry, Vaccinium myrtilloides, Production in Eastern Ontario. Can. Field-Nat. 2004, 118, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Braid, A.C.R.; Manzer, D.; Nielsen, S.E. Wildlife habitat enhancements for grizzly bears: Survival rates of planted fruiting shrubs in forest harvests. For. Ecol. Manag. 2016, 369, 144–154. [Google Scholar] [CrossRef]
- Roberts, D.R.; Nielsen, S.E.; Stenhouse, G.B. Idiosyncratic responses of grizzly bear habitat to climate change based on projected food resource changes. Ecol. Appl. 2014, 24, 1144–1154. [Google Scholar] [CrossRef]
- Garibaldi, A. Moving from model to application: Cultural keystone species and reclamation in Fort McKay, Alberta. J. Ethnobiol. 2009, 29, 323–338. [Google Scholar] [CrossRef]
- Golzadeh, N.; Barst, B.D.; Basu, N.; Baker, J.M.; Auger, J.C.; McKinney, M.A. Evaluating the concentrations of total mercury, methylmercury, selenium, and selenium:mercury molar ratios in traditional foods of the Bigstone Cree in Alberta, Canada. Chemosphere 2020, 250, 126285. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M. Do berries listen? Berries as indicators, ancestors, and agents in Canada’s Oil Sands Region. Ethnos 2020. [Google Scholar] [CrossRef]
- Jarron, L.R.; Coops, N.C.; Mackenzie, W.H.; Tompalski, P.; Dykstra, P. Detection of sub-canopy forest structure using airborne LiDAR. Remote Sens. Environ. 2020, 244, 111770. [Google Scholar] [CrossRef]
- Berkes, F.; Davidson-Hunt, I.J. Biodiversity, traditional management systems, and cultural landscapes: Examples from the boreal forest of Canada. Int. Soc. Sci. J. 2006, 58, 35–47. [Google Scholar] [CrossRef]
- Seeram, N.P. Berry fruits: Compositional elements, biochemical activities, and the impact of their intake on human health, performance, and disease. J. Agric. Food Chem. 2008, 56, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Munro, R.H.M.; Bainbridge, E.L.; Stenhouse, G.B.; Boyce, M.S. Grizzly bears and forestry II. Distribution of grizzly bear foods in clearcuts of west-central Alberta, Canada. For. Ecol. Manag. 2004, 199, 67–82. [Google Scholar] [CrossRef]
- Garibaldi, A.; Straker, J. Cultural keystone species in oil sands mine reclamation, Fort McKay, Alberta, Canada. In Proceedings of the British Columbia Mine Reclamation Symposium. University of British Columbia Library, Cranbrook, BC, Canada, 14–17 September 2009. [Google Scholar]
- Garibaldi, A.; Turner, N. Cultural keystone species: Implications for ecological conservation and restoration. Ecol. Soc. 2004, 9. [Google Scholar] [CrossRef]
- Marles, R.; Clavelle, C.; Monteleone, L.; Tays, N.; Burns, D. Aboriginal Plant Use in Canada’s Northwest Boreal Forest; Natural Resources Canada: Edmonton, AB, Canada, 2012. [Google Scholar]
- Kloet, V. The Genus Vaccinium in North America; Agriculture Canada Research Branch: Ottawa, ON, Canada, 1988. [Google Scholar]
- Smith, D.W. Ecological studies of Vaccinium species in Alberta. Can. J. Plant Sci. 1962, 82–90. [Google Scholar] [CrossRef]
- Alberta Environment and Sustainable Resource Development. Regional Forest Landscape Assessment—Lower Athabasca Region; Alberta Environment and Sustainable Resource Development: Edmonton, AB, Canada, 2012. [Google Scholar]
- Raine, M.; Mackenzie, I.; Gilchrist, I. CNRL Horizon Project Environmental Impact Assessment; Wetlands and Forest Resources Baseline: Calgary, AB, Canada, 2002; Volume 6. [Google Scholar]
- de Groot, W.J.; Flannigan, M.D.; Cantin, A.S. Climate change impacts on boreal fire regimes. For. Ecol. Manag. 2013, 294, 35–44. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Dennett, J.M.; Bater, C.W. Landscape patterns of rare vascular plants in the Lower Athabasca. Forests 2020, 11, 699. [Google Scholar] [CrossRef]
- Zhang, J.; Nielsen, S.E.; Grainger, T.N.; Kohler, M.; Chipchar, T.; Farr, D.R. Sampling plant diversity and rarity at landscape scales: Importance of sampling time in species detectability. PLoS ONE 2014, 9, e95334. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberon, J.; Pearson, R.G.; Anderson, R.P.; Martinez-Meyer, E.; Nakamura, M.; Araujo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Nielsen, S.E. Fruiting Shrubs of the Lower Athabasca: Distribution, Ecology and a Digital Atlas; ACE-Lab: Edmonton, AB, Canada, 2016; p. 81. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Johnson, C.J.; Heard, D.C.; Boyce, M.S.; Nielsen, S.E.; Johnson, C.J.; Heard, D.C.; Boyce, M.S. Can models of presence-absence be used to scale abundance? Two case studies considering extremes in life history. Ecography 2005, 28, 197–208. [Google Scholar] [CrossRef]
- Baum, C.F. Modeling proportions. STATA J. 2008, 8, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Papke, L.E.; Wooldridge, J.M. Econometric methods for fractional response variables with an application to 401(k) plan participation rates. J. Appl. Econom. 1996, 11, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Soil Landscapes of Canada Working Group. Available online: https://open.canada.ca/data/en/dataset/5ad5e20c-f2bb-497d-a2a2-440eec6e10cd (accessed on 1 September 2010).
- Evans, J. CTI.aml Compound Topographic Index AML Script 2004. Available online: http://arcscripts.esri.com/details.asp?dbid=11863 (accessed on 1 September 2010).
- Jenness, J. Topographic Position Index (tpi_jen.avx) Extension for ArcView 3.x, v. 1.2. 2006. Available online: http://www.jennessent.com/arcview/tpi.htm (accessed on 1 September 2010).
- McCune, B. Improved estimates of incident radiation and heat load using non-parametric regression against topographic variables. J. Veg. Sci. 2007, 18, 751–754. [Google Scholar] [CrossRef]
- Mbogga, M.S.; Hansen, C.; Wang, T.; Hamann, A. A Comprehensive Set of Interpolated Climate Data for Alberta; 2010. Available online: https://open.alberta.ca/publications/9780778591849 (accessed on 1 September 2010).
- Environment and Sustainable Resource Environment. General Specifications for the Acquisition of Lidar Data; Informatics Branch, Government of Alberta: Edmonton, AB, Canada; p. 52.
- McGaughey, R. FUSION/LVD: Software for LIDAR Data Analysis and Visualization 2016. Available online: http://forsys.cfr.washington.edu/FUSION/fusion_overview.html (accessed on 1 July 2010).
- StataCorp Stata Statistical Software: Release 14 2015. Available online: www.stata.com (accessed on 1 July 2010).
- Bursac, Z.; Gauss, C.H.; Williams, D.K.; Hosmer, D.W. Purposeful selection of variables in logistic regression. Source Code Biol. Med. 2008, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Lemeshow, S.; Hosmer, D.W. A review of goodness of fit statistics for use in the development of logistic regression models. Am. J. Epidemiol. 1982, 115, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Manel, S.; Williams, H.; Ormerod, S. Evaluating presence-absence models in diversity on the roof of the world: Spatial patterns and environmental determinants. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raes, N.; ter Steege, H. A null-model for significance testing of presence-only species distribution models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Moola, F.M. Yield and Morphological Responses of Wild Blueberry (Vaccinium spp.) to Forest Harvesting and Conifer Release Treatments. Master’s Thesis, Lakehead University, Thunder Bay, ON, Canada, 1997. [Google Scholar]
- Kolb, A.; Barsch, F.; Diekmann, M. Determinants of local abundance and range size in forest vascular plants. Glob. Ecol. Biogeogr. 2006, 15, 237–247. [Google Scholar] [CrossRef]
- Flora of North America Editorial Committee. Flora of North America North of Mexico; Flora of North America Editorial Committee: New York, NY, USA, 1993. [Google Scholar]
- Okland, R.H. Persistence of vascular plants in a Norwegian boreal coniferous forest. Ecography 1995, 18, 3–14. [Google Scholar] [CrossRef]
- Strengbom, J.; Axelsson, E.P.; Lundmark, T.; Nordin, A. Trade-offs in the multi-use potential of managed boreal forests. J. Appl. Ecol. 2018, 55, 958–966. [Google Scholar] [CrossRef]
- Penney, B.G.; McRae, K.B.; Rayment, A.F. Long-term effects of burn-pruning on lowbush blueberry (Vaccinium angustifolium Ait.) production. Can. J. Plant Sci. 1997, 77, 421–425. [Google Scholar] [CrossRef]
- Michell, H.J. Gathering berries in northern contexts: A Woodlands Cree metaphor for community-based research. Pimatisiwin 2009, 7, 65–73. [Google Scholar]






| Variable | β | SE | p |
|---|---|---|---|
| Shrub abundance to 1 | 3.721 | 0.768 | 0.001 |
| Canopy, (√percent) | −0.230 | 0.079 | 0.004 |
| Clay, percent | −0.069 | 0.019 | 0.001 |
| Heatload, index | 0.010 | 0.003 | 0.001 |
| Constant | −21.079 | 5.543 | 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, S.E.; Dennett, J.M.; Bater, C.W. Predicting Occurrence, Abundance, and Fruiting of a Cultural Keystone Species to Inform Landscape Values and Priority Sites for Habitat Enhancements. Forests 2020, 11, 783. https://doi.org/10.3390/f11070783
Nielsen SE, Dennett JM, Bater CW. Predicting Occurrence, Abundance, and Fruiting of a Cultural Keystone Species to Inform Landscape Values and Priority Sites for Habitat Enhancements. Forests. 2020; 11(7):783. https://doi.org/10.3390/f11070783
Chicago/Turabian StyleNielsen, Scott E., Jacqueline M. Dennett, and Christopher W. Bater. 2020. "Predicting Occurrence, Abundance, and Fruiting of a Cultural Keystone Species to Inform Landscape Values and Priority Sites for Habitat Enhancements" Forests 11, no. 7: 783. https://doi.org/10.3390/f11070783
APA StyleNielsen, S. E., Dennett, J. M., & Bater, C. W. (2020). Predicting Occurrence, Abundance, and Fruiting of a Cultural Keystone Species to Inform Landscape Values and Priority Sites for Habitat Enhancements. Forests, 11(7), 783. https://doi.org/10.3390/f11070783