Allometric Equations for Predicting Agave lechuguilla Torr. Aboveground Biomass in Mexico

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
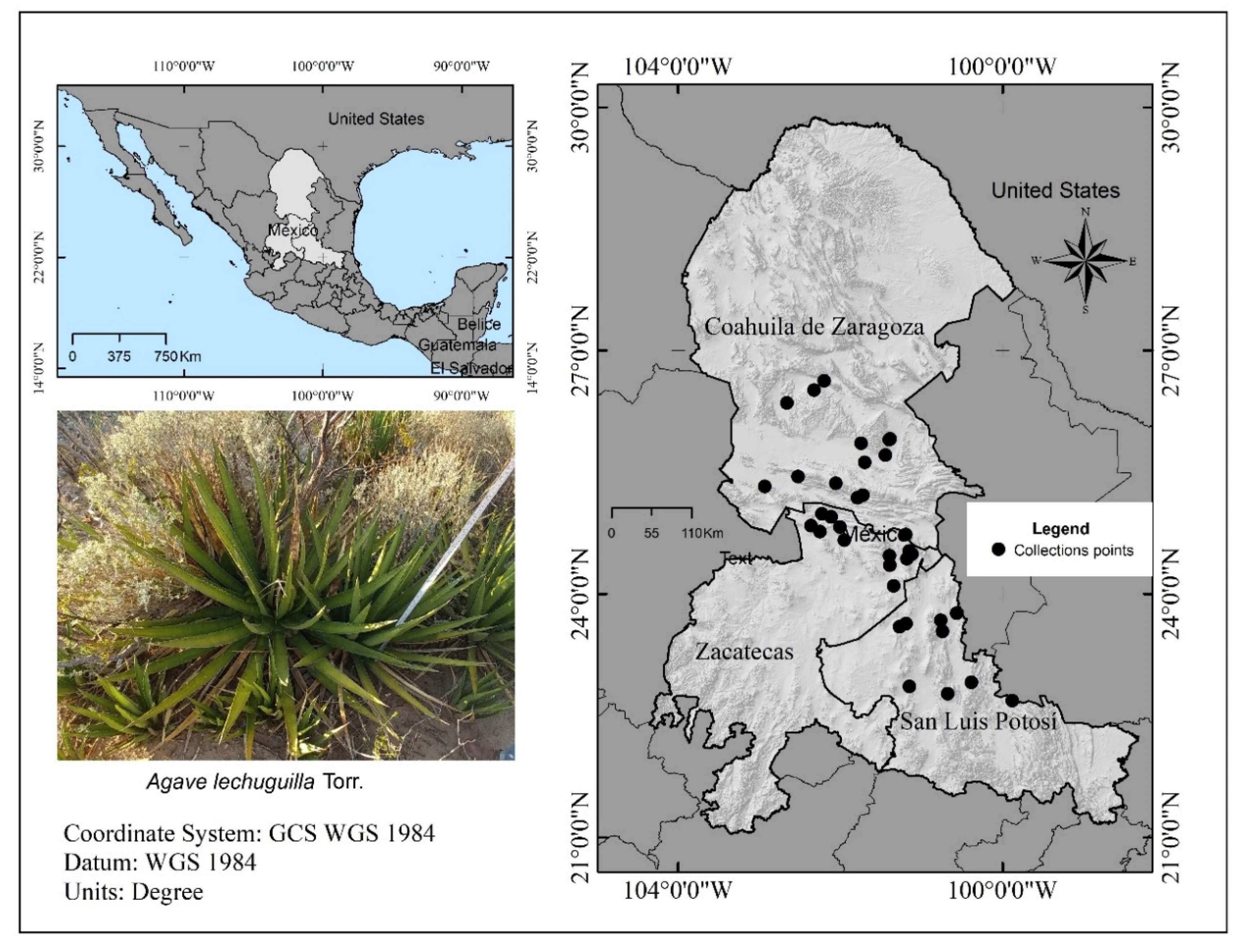
2.1. Study Area
2.2. Sampling of Aboveground Biomass
2.3. Statistical Analysis
2.4. Adaptation of the Regression Model
2.5. Robust Regression Techniques
3. Results and Discussion
3.1. Descriptive Statistics Within Its Algorithm
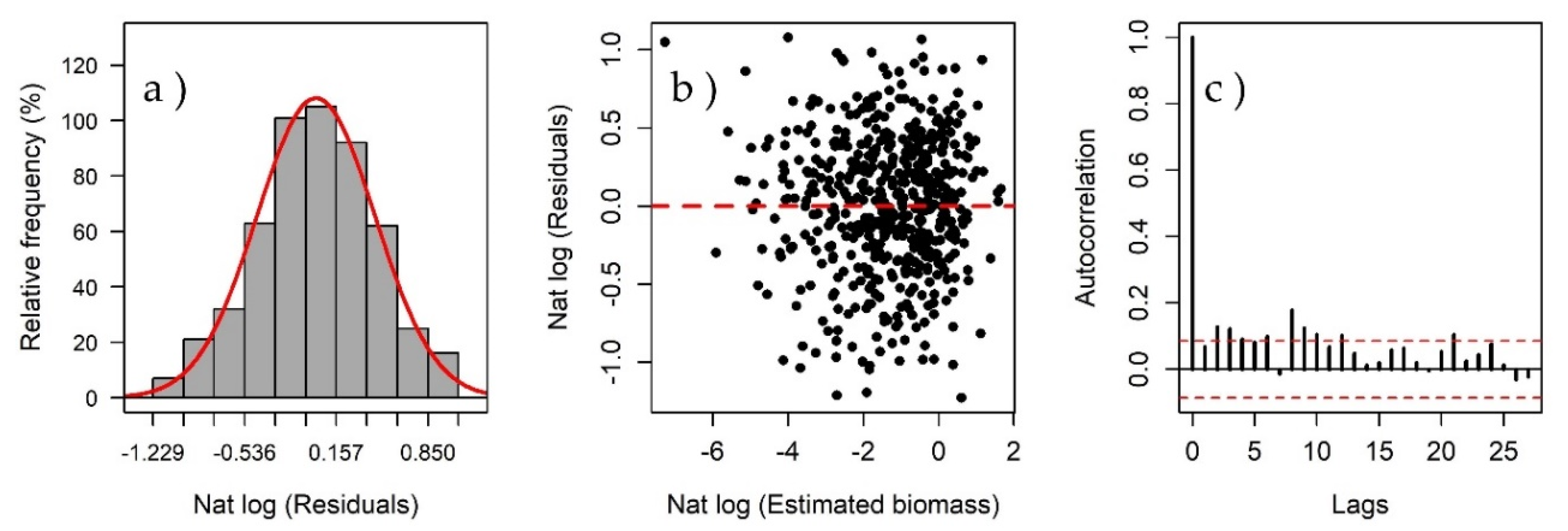
3.2. Model Fit and Detection of Atypical Observations
3.3. Selection of the Best Model
3.4. Model Validation
3.5. Robust Estimation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Climate Change: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, NY, USA, 2014; 1435p. [Google Scholar]
- National Oceanic and Atmospheric Administration (NOAA). 2019. Available online: https://www.noaa.gov/news/global-carbon-dioxide-growth-in-2018-reached-4th-highest-on-record (accessed on 2 May 2020).
- Comisión Europea (CE). El Papel de la Naturaleza en el Cambio Climático. 2009. Available online: https://ec.europa.eu/environment/pubs/pdf/factsheets/Nature%20and%20Climate%20Change/Nature%20and%20Climate%20Change_ES.pdf (accessed on 2 May 2020).
- Singh, G.; Singh, B. Biomass equations and assessment of carbon stock of Calligonum polygonoides L., a shrub of Indian arid zone. Curr. Sci. 2017, 112, 2456. [Google Scholar] [CrossRef]
- Forestry National Commission (CONAFOR). Bosques, Cambio Climático y REDD+ en México. Guía Básica; Área de Mercados y Proyectos Forestales de Carbon: Jalisco, México, 2012; 56p. [Google Scholar]
- Zamora, M.M. Cambio climático. Rev. Mex. Cienc. For. 2015, 6, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, R.; Ahlawat, S.; Gupta, A. Production of wood biomass by high density Acacia nilotica plantation in semi-arid region of central India. Rang. Man. Agrofor. 2014, 35, 128–132. [Google Scholar]
- Briones, O.; Búrquez, A.; Martínez-Yrízar, A.; Pavón, N.; Perroni, Y. Biomasa y productividad en las zonas. Áridas Mexicanas. Madera Bosques 2018, 24, e2401898. [Google Scholar] [CrossRef]
- Global Terrestrial Observing System (GTOS). Terrestrial Essential Climate Variables. For Climate Change Assessment, Mitigation and Adaptation; FAO: Rome, Italy, 2008; 41p, Available online: http://www.fao.org/3/a-i0197e.pdf (accessed on 7 June 2020).
- Brown, S. Tropical forests and the global carbon cycle: Estimating state and change in biomass density. In Forest Ecosystems, Forest Management and the Global Carbon Cycle; Springer: Berlin/Heidelberg, Germany, 1996; Volume 40, pp. 135–144. [Google Scholar] [CrossRef]
- Návar, J.; Rodriguez-Flores, F.J.; Rios-Saucedo, J. Biomass estimation equations for mesquite trees in the Americas. PeerJ 2019, 7, e6782. [Google Scholar] [CrossRef] [Green Version]
- Montaño, N.M.; Ayala, F.; Bullock, S.H.; Briones, O.; García, O.F.; García, S.R.; Maya, Y.; Perroni, Y.; Siebe, C.; Tapia, T.Y.; et al. Almacenes y flujos de carbono en ecosistemas áridos y semiáridos de México: Síntesis y perspectivas. Terra Latinoamericana 2016, 34, 39–59. [Google Scholar]
- Nobel, P.S.; Quero, E. Environmental productivity indices for a Chihuahuan desert CAM plant. Agave Lechuguilla Ecol. 1986, 67, 1–11. [Google Scholar] [CrossRef]
- Reyes, A.J.; Aguirre, R.R.; Peña, V.C. Biología y aprovechamiento de Agave lechuguilla Torrey. Bol. Soc. Bot. México 2000, 1, 75–88. [Google Scholar] [CrossRef]
- Picard, N.; Saint-André, L.; Henry, M. Manual de Construcción de Ecuaciones Alométricas para Estimar el Volumen y la Biomasa de los Árboles: Del Trabajo de Campo a la Predicción; FAO: Rome, Italy, 2012. [Google Scholar]
- Singh, T.S.P.; Verma, A.; Kumar, P.; Meherul, A.N.; Krishna, B.R. Biomass and carbon projection models in Hardwickia binata Roxb. vis a vis estimation of its carbon sequestration potential under arid environment. Arch. Agron. Soil Sci. 2019. [Google Scholar] [CrossRef]
- Huff, S.; Ritchie, M.; Temesgen, H. Allometric equations for estimating aboveground biomass for common shrubs in northeastern California. For. Ecol. Man. 2017, 398, 48–63. [Google Scholar] [CrossRef]
- Latifi, H.; Fassnacht, F.E.; Hartig, F.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Stratified aboveground forest biomass estimation by remote sensing data. Int. J. Appl. Earth. Obs. 2015, 38, 229–241. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Neumann, M.; Saatchi, S.S.; Ulander, L.M.H.; Fransson, J.E.S. Assessing performance of L- and P-band polarimetric interferometric SAR data in estimating boreal forest above-ground biomass. IEEE Trans. Geosci. Remote Sens. 2012, 50, 714–726. [Google Scholar] [CrossRef]
- Faraway, J.J. Linear Models with R, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2014; 274p. [Google Scholar]
- Montgomery, D.C.; Peck, E.A.; Vining, G.G. Introduction to Linear Regression Analysis, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012; 688p. [Google Scholar]
- Segura, M.; Andrade, H. ¿Cómo hacerlo? ¿Cómo construir modelos alométricos de volumen, biomasa o carbono de especies leñosas perennes? Agroforestería Américas 2008, 1, 89–96. [Google Scholar]
- Li, L.X.; Hao, Y.H.; Zhang, Y. The application of dummy variables in statistic analysis. J. Math. Med. 2006, 19, 51–52. (In Chinese) [Google Scholar]
- Fox, J. Applied Regression Analysis and Generalized Linear Models, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2016; 791p. [Google Scholar]
- Weisberg, S. Applied Linear Regression, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2014; 368p. [Google Scholar]
- Zeng, W.S.; Zhang, H.R.; Tang, S.Z. Using the dummy variable model approach to construct compatible single-tree biomass equations at different scales—A case study for Masson pine (Pinus massoniana) in southern China. Can. J. For. Res. 2011, 41, 1547–1554. [Google Scholar] [CrossRef]
- Pando-Moreno, M.; Pulido, R.; Castillo, D.; Jurado, E.; Jimenez, J. Estimating fiber for lechuguilla (Agave lecheguilla Torr., Agavaceae), a traditional non-timber forest product in Mexico. For. Ecol. Man. 2008, 255, 3686–3690. [Google Scholar] [CrossRef]
- Narcia, V.M.; Castillo, Q.D.; Vázquez, R.J.; Berlanga, R.C. Turno técnico de la lechuguilla (Agave lechuguilla Torr.) en el noreste de México. Rev. Mex. Cienc. For. 2012, 3, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Molina-Guerra, V.M.; Soto-Mata, B.; Cervantes-Balderas, J.M.; Alanís, R.E.; Marroquín-Castillo, J.J.; Sarmiento-Muñoz, T.I. Composición y estructura del matorral desértico rosetófilo del sureste de Coahuila, México. Polibotánica. 2017, 1, 67–77. [Google Scholar] [CrossRef]
- Alanís-Rodríguez, E.; Mora-Olivo, A.; Jiménez-Pérez, J.; González-Tagle, M.A.; Yerena Yamallel, J.I.; Martínez-Ávalos, J.G.; González-Rodríguez, L.E. Composición y diversidad del matorral desértico rosetófilo en dos tipos de suelo en el noreste de México. Acta Bot. Mex. 2015, 110, 105–117. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Statistic and Geography (INEGI). Conjuntos de Datos Vectoriales de Uso del Suelo y Vegetación, Escala 1:250,000 Serie VI; INEGI: Aguascalientes, México, 2016. [Google Scholar]
- Rzedowski, J. Vegetación de México, 1st ed.; Digital; Comisión Nacional para el Uso y Conservación de la Biodiversidad: Mexico City, México, 2006; 504p. [Google Scholar]
- García, E. Climas, Clasificación de Kóeppen, Modificado por García. Carta de Climas, Escala 1:1 000 000; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO): Mexico City, México, 1998. [Google Scholar]
- Schumacher, F.X.; Hall, F.S. Logarithmic expression of timber-tree volume. J. Agric. Res. 1933, 47, 719–734. [Google Scholar]
- Villavicencio, G.E.; Mendoza-Morales, S.; Méndez, G.J. Modelo para predecir biomasa foliar seca de Litsea parvifolia (Hemsl.) Mez. Rev. Mex. Cienc. For. 2020, 11, 112–133. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Ramos, A.; Cano-Pineda, A.; Flores-López, C.; Hernández-Ramos, J.; García-Cuevas, X.; Martínez-Salvador, M.; Martínez, Á.L. Modelos para estimar biomasa de Euphorbia antisyphilitica Zucc. en seis municipios de Coahuila. Madera Bosques 2019, 25, 1–13. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 February 2020).
- Sprugel, D.G. Correcting for bias in log-transformed allometric equations. Ecology 1983, 64, 209–210. [Google Scholar] [CrossRef]
- Ortiz, P.J.; Gil, D. Transformaciones logarítmicas en regresión simple. Comun. Estadística 2014, 7, 89–98. [Google Scholar]
- Nikolai, S.A. qpcR: Modelling and Analysis of Real-Time PCR Data. R Package Version 1.4–1. 2018. Available online: https://CRAN.R-project.org/package=qpcR (accessed on 7 July 2020).
- Hong, X.; Sharkey, P.M.; Warwick, K. A robust nonlinear identification algorithm using PRESS statistic and forward regression. IEEE Trans. Neural Netw. 2003, 14, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Khaleelur, R.S.; Mohamed, S.M.; Senthamarai, K.K. Multiple linear regression models in outlier detection. Int. J. Adv. Res. Comput. Sci. 2012, 2, 23–28. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; 798p. [Google Scholar]
- Gross, J.; Ligges, U. Nortest: Tests for Normality. R Package Version 1.0–4. 2015. Available online: https://CRAN.R-project.org/package=nortest (accessed on 7 April 2020).
- Zeileis, A.; Hothorn, T. Diagnostic checking in regression relationships. R News 2002, 2, 7–10. [Google Scholar]
- Yan, X.; Gang Su, X. Linear Regression Analysis: Theory and Computing, 1st ed.; World Scientific: Tuck Link, Singapore, 2009; 348p. [Google Scholar]
- Yohai, V.J. High breakdown-point and high efficiency robust estimates for regression. Ann. Stat. 1987, 15, 642–656. [Google Scholar] [CrossRef]
- Huber, P.J. Robust Statistics; Wiley: New York, NY, USA, 1981. [Google Scholar]
- Maechler, M.; Rousseeuw, P.; Croux, C.; Todorov, V.; Ruckstuhl, A.; Salibian-Barrera, M.; Verbeke, T.; Koller, K.; Conceicao, E.; di Palma, A.M. robustbase: Basic Robust Statistics R Package Version 0.93-6. 2020. Available online: http://CRAN.R-project.org/package=robustbase (accessed on 2 April 2020).
- Pollar, D. Asymptotics for least absolute deviation regression estimators. Econ. Theory 1991, 7, 186–199. [Google Scholar] [CrossRef]
- Osorio, F.; Wolodzko, T. Routines for L1 Estimation. R Package Version 0.38.19. 2017. Available online: http://l1pack.mat.utfsm.cl (accessed on 3 April 2020).
- Goldstein, H. Restricted unbiased iterative generalized least-squares estimation. Biometrika 1989, 76, 622–623. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Versión 3.1–147. 2020. Available online: https://CRAN.R-project.org/package=nlme (accessed on 4 April 2020).
- Conti, G.; Enrico, L.; Casanoves, F.; Diaz, S. Shrub biomass estimation in the semiarid Chaco forest: A contribution to the quantification of an underrated carbon stock. Ann. For. Sci. 2013, 70, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Aquino-Ramírez, M.; Velázquez-Martínez, A.; Castellanos-Bolaños, J.F.; De los Santos-Posadas, H.; Etchevers-Barra, J.D. Partición de la biomasa aérea en tres especies arbóreas tropicales. Agrociencia 2015, 49, 299–314. [Google Scholar]
- Cortés-Sánchez, B.G.; Ángeles-Pérez, G.; De los Santos-Posadas, H.M.; Ramírez-Maldonado, H. Ecuaciones alométricas para estimar biomasa en especies de encino en Guanajuato, México. Madera Bosques 2019, 25, e2521799. [Google Scholar] [CrossRef]
- Rasch, D.; Verdooren, R.; Pilz, J. Applied Statistics: Theory and Problem Solutions with R; John Wiley & Sons: Oxford, UK, 2020; 512p. [Google Scholar]
- Ali, A.; Xu, M.S.; Zhao, Y.T.; Zhang, Q.Q.; Zhou, L.L.; Yang, X.D.; Yan, E.R. Allometric biomass equations for shrub and small tree species in subtropical China. Silva Fennica 2015, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio, G.E.; Hernández, R.A.; García, C.X. Estimación de la biomasa foliar seca de Lippia graveolens Kunth del sureste de Coahuila. Rev. Mex. Cienc. For. 2018, 9, 187–207. [Google Scholar] [CrossRef]
- Araújo, E.J.G.; Loureiro, G.H.; Sanquetta, C.R.; Sanquetta, M.N.I.; Corte, A.P.D.; Péllico Netto, S.; Behling, A. Allometric models to biomass in restoration areas in the Atlantic rain forest. Floresta Ambiente 2018, 25, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Méndez, G.J.; Turlán, M.O.A.; Ríos, S.J.C.; Nájera, L.J.A. Ecuaciones alométricas para estimar biomasa aérea de Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst. Rev. Mex. Cienc. For. 2012, 3, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.Q.; Liu, Q.J.; Feng, Z.W.; Ma, Z.Q. Biomass equations for four shrub species in subtropical China. J. For. Res. 2010, 15, 83–90. [Google Scholar] [CrossRef]
- Bondé, L.; Ganamé, M.; Ouédraogo, O.; Nacoulma, B.M.; Thiombiano, A.; Boussim, J.I. Allometric models to estimate foliage biomass of Tamarindus indica in Burkina Faso, Southern Forests. J. For. Sci. 2018, 80, 143–150. [Google Scholar] [CrossRef]
- Búrquez, A.; Martínez-Yrízar, A. Accuracy and bias on the estimation of aboveground biomass in the woody vegetation of the Sonoran Desert. Botany 2011, 89, 625–633. [Google Scholar] [CrossRef]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- Cunia, T. Error of forest inventory estimates: Its main components. In Estimating Tree Biomass Regressions and Their Error. Proceedings of the Workshop on Tree Biomass Regression Functions and Their Contribution to the Error of Forest Inventory Estimates; General Technical Bulletin NE-GTR-117; USDA Forest Service: Broomall, PA, USA, 1987; pp. 1–13. [Google Scholar]
- Alonso, J.C.; Montenegro, S. Estudio de Monte Carlo para comparar 8 pruebas de normalidad sobre residuos de mínimos cuadrados ordinarios en presencia de procesos autorregresivos de primer orden. Estud. Gerenc. 2015, 31, 253–265. [Google Scholar] [CrossRef] [Green Version]
- Cancino, C.J. Dendrometría Básica; Departamento Manejo de Bosques y Medio Ambiente, Facultad de Ciencias Forestales, Universidad de Concepción: Concepción, Chile, 2012; Available online: http://repositorio.udec.cl/bitstream/11594/407/2/Dendrometria_Basica.pdf (accessed on 8 May 2020).
- Owate, O.A.; Mware, M.J.; Kinyanjui, M.J. Allometric equations for estimating silk oak (Grevillea robusta) biomass in agricultural landscapes of Maragua Subcounty, Kenya. Int. J. Forest Res. 2018. [Google Scholar] [CrossRef]
- Moore, J.R. Allometric equations to predict the total above-ground biomass of radiata pine trees. Ann. For. Sci. 2010, 67, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ponce, A.M. Medidas de influencia que se basan en la curva de influencia. Pesquimat 2000, 3, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.R.; Montgomery, D.C. A robust regression technique using compound estimation. Nav. Res. Logist. 1998, 45, 125–139. [Google Scholar] [CrossRef]
- Alma, O.G. Comparison of robust regression methods in linear regression. Int. J. Contemp. Math. Sci. 2011, 6, 409–421. [Google Scholar]
- Smucler, E.; Yohai, V.J. Robust and sparse estimators for linear regression models. Comput. Stat. Data Anal. 2017, 111, 116–130. [Google Scholar] [CrossRef] [Green Version]
- Smucler, E. Estimadores Robustos para el Modelo de Regresión Lineal con Datos de Alta Dimensión. Ph.D. Thesis, Universidad de Buenos Aires, Buenos Aires, Argentina, 2016. [Google Scholar]
- Van Aelst, S.; Willems, G.; Zamar, R.H. Robust and efficient estimation of the residual scale in linear regression. J. Multivar. Anal. 2013, 116, 278–296. [Google Scholar] [CrossRef]
- Alfons, A.; Croux, C.; Gelper, S. Sparse least trimmed squares regression for analyzing high-dimensional large data sets. Ann. Appl. Stat. 2013, 7, 226–248. [Google Scholar] [CrossRef]
- Susanti, Y.; Pratiwi, H.; Sulistijowati, H.S.; Liana, T. M estimation, S estimation, and Mm estimation in robust regression. Int. J. Pure Appl. Math. 2014, 91, 349–360. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coahuila (n = 175) | San Luis Potosí (n = 178) | Zacatecas (n = 180) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Cd | H | AGB | Cd | H | AGB | Cd | H | AGB |
| Minimum | 7.50 | 9.00 | 0.01 | 5.10 | 6.10 | 0.00 | 3.50 | 3.50 | 0.00 |
| Maximum | 128.50 | 95.00 | 2.03 | 166.50 | 118.00 | 8.17 | 127.50 | 87.00 | 2.91 |
| Mean | 48.66 | 44.02 | 0.49 | 54.82 | 45.60 | 0.89 | 49.40 | 41.75 | 0.45 |
| Standard deviation | 25.90 | 16.90 | 0.47 | 36.24 | 23.50 | 1.28 | 29.32 | 17.89 | 0.54 |
| C.V. | 53.22 | 38.40 | 96.16 | 66.10 | 51.53 | 144.03 | 59.35 | 42.86 | 120.24 |
| Equation | Estimator | Value | Sxy (β) | Value t | Pr (>|t|) | R2 adj. | Sxy |
|---|---|---|---|---|---|---|---|
| 3 | β0 | −8.722 | 0.132 | −65.87 | 0.0001 | 0.865 | 0.558 |
| β1 (ln Cd) | 2.001 | 0.035 | 57.866 | 0.0001 | |||
| β2 [Zac] | −0.301 | 0.051 | −5.897 | 0.0001 | |||
| 4 | β0 | −10.183 | 0.155 | −65.665 | 0.0001 | 0.901 | 0.478 |
| β1 (ln Cd) | 1.108 | 0.071 | 15.657 | 0.0001 | |||
| β2 (ln H) | 1.285 | 0.093 | 13.89 | 0.0001 | |||
| β3 [Zac] | −0.178 | 0.051 | −3.481 | 0.0001 | |||
| β4 [SLP] | 0.127 | 0.051 | 2.496 | 0.0100 |
| Equation | Estimator | Valor | IC | Pr (>|t|) | R2 adj. | Sxy | PRESS | AIC | CF |
|---|---|---|---|---|---|---|---|---|---|
| 3 | β0 | −8.762 | (± 0.249) | 0.0001 | 0.877 | 0.531 | 149.55 | 834 | 1.151 |
| β1 (ln Dp) | 2.014 | (± 0.065) | 0.0001 | ||||||
| β2 [Zac] | −0.299 | --- | 0.0001 | ||||||
| 4 | β0 | −10.182 | (± 0.285) | 0.0001 | 0.914 | 0.44 | 102.25 | 632.2 | 1.101 |
| β1 (ln Dp) | 1.158 | (± 0.130) | 0.0001 | ||||||
| β2 (ln H) | 1.236 | (± 0.169) | 0.0001 | ||||||
| β3 [Zac] | −0.178 | --- | 0.0001 | ||||||
| β4 [SLP] | 0.143 | --- | 0.01 |
| Estimator | OLS | MM | LAD | LTS | GLS |
|---|---|---|---|---|---|
| β0 | −10.183 * (±0.304) | −10.214 *(±0.300) | −10.244 * (±0.467) | −10.349 * | −9.959 * (±0.328) |
| β1 | 1.108 * (±0.139) | 1.129 * (±0.136) | 1.155 * (±0.214) | 1.181 * | 1.048 * (±0.138) |
| β2 | 1.285 * (±0.181) | 1.273 * (±0.177) | 1.245 * (±0.279) | 1.208 * | 1.285 * (±0.180) |
| β3 (Zac) | −0.178 * (±0.100) | −0.161 * (±0.098) | −0.099 ** (±0.072) | −0.098 * | −0.179 * (±0.113) |
| β4(SLP) | 0.127 ** (±0.100) | 0.147 * (±0.098) | 0.210 (±0.154) | 0.195 *** | 0.127 ** (±0.113) |
| R2 adj. | 0.901 | 0.907 | 0.901 | 0.919 | 0.901 |
| MSE | 0.226 | 0.226 | 0.228 | 0.229 | 0.228 |
| CF | 1.121 | 1.111 | ---- | 1.092 | 1.120 |
| Normality (LF) | 0.043 | 0.053 | 0.022 | 0.014 | 0.014 |
| Homogeneity (B-P) | 0.031 | 0.031 | 0.031 | 0.031 | ---- |
| Autocorrelation (LJ-B) | 0.007 | 0.013 | 0.013 | 0.015 | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Hernández, C.d.J.; Méndez-González, J.; Sánchez-Pérez, F.d.J.; Méndez-Encina, F.M.; López-Díaz, Ó.M.; López-Serrano, P.M. Allometric Equations for Predicting Agave lechuguilla Torr. Aboveground Biomass in Mexico. Forests 2020, 11, 784. https://doi.org/10.3390/f11070784
Flores-Hernández CdJ, Méndez-González J, Sánchez-Pérez FdJ, Méndez-Encina FM, López-Díaz ÓM, López-Serrano PM. Allometric Equations for Predicting Agave lechuguilla Torr. Aboveground Biomass in Mexico. Forests. 2020; 11(7):784. https://doi.org/10.3390/f11070784
Chicago/Turabian StyleFlores-Hernández, Cristóbal de J., Jorge Méndez-González, Félix de J. Sánchez-Pérez, Fátima M. Méndez-Encina, Óscar M. López-Díaz, and Pablito M. López-Serrano. 2020. "Allometric Equations for Predicting Agave lechuguilla Torr. Aboveground Biomass in Mexico" Forests 11, no. 7: 784. https://doi.org/10.3390/f11070784
APA StyleFlores-Hernández, C. d. J., Méndez-González, J., Sánchez-Pérez, F. d. J., Méndez-Encina, F. M., López-Díaz, Ó. M., & López-Serrano, P. M. (2020). Allometric Equations for Predicting Agave lechuguilla Torr. Aboveground Biomass in Mexico. Forests, 11(7), 784. https://doi.org/10.3390/f11070784