Effect of Beauveria bassiana Fungal Infection on Survival and Feeding Behavior of Pine-Tree Lappet Moth (Dendrolimus pini L.)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Insects
2.2. Fungal Isolation
2.3. Preparation of Spore Suspensions
2.4. Confirmation of Conidial Viability
2.5. Setting up the Experiments
2.6. Statistical Analyses
3. Results
3.1. Experiment 1
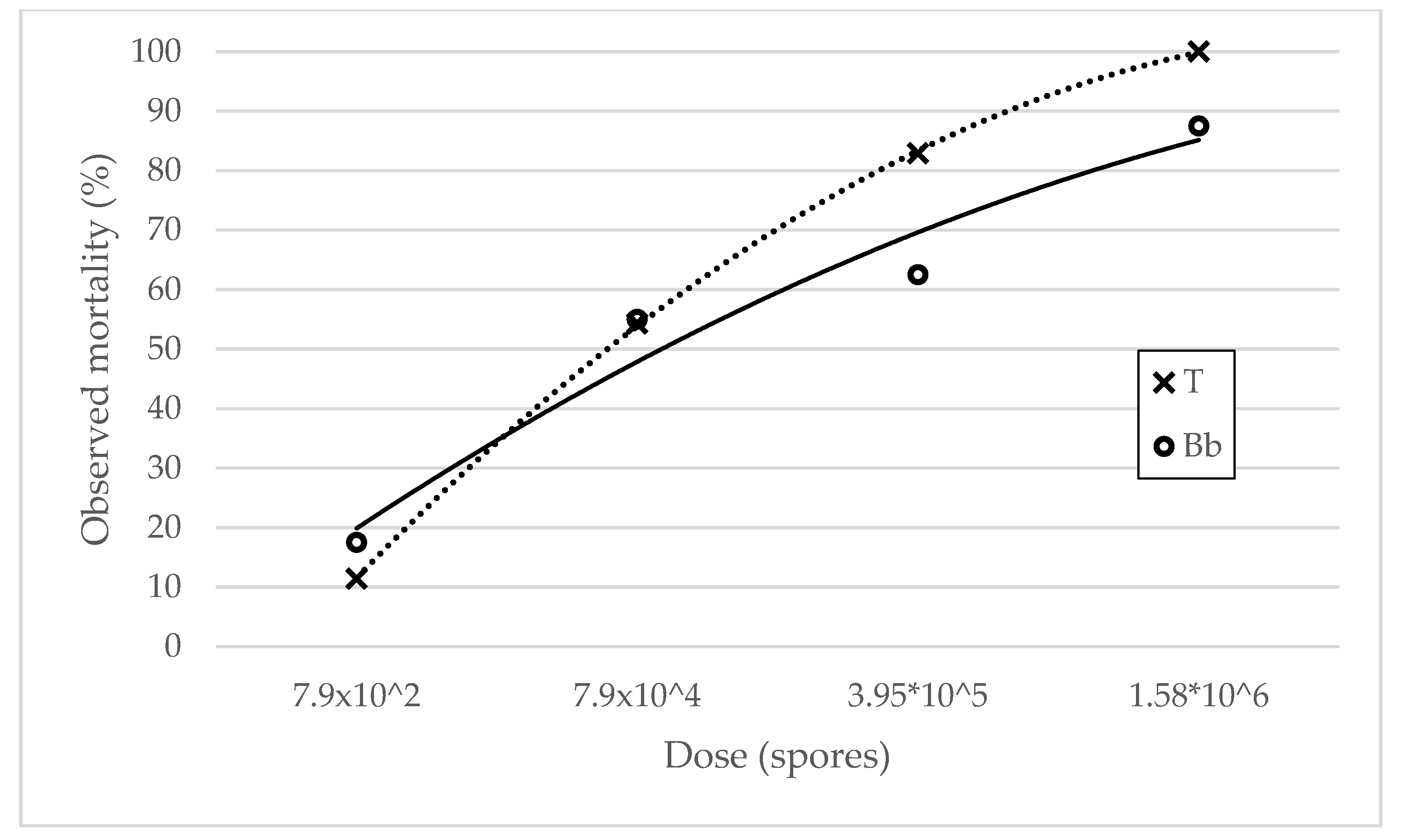
3.1.1. Mortality Rates
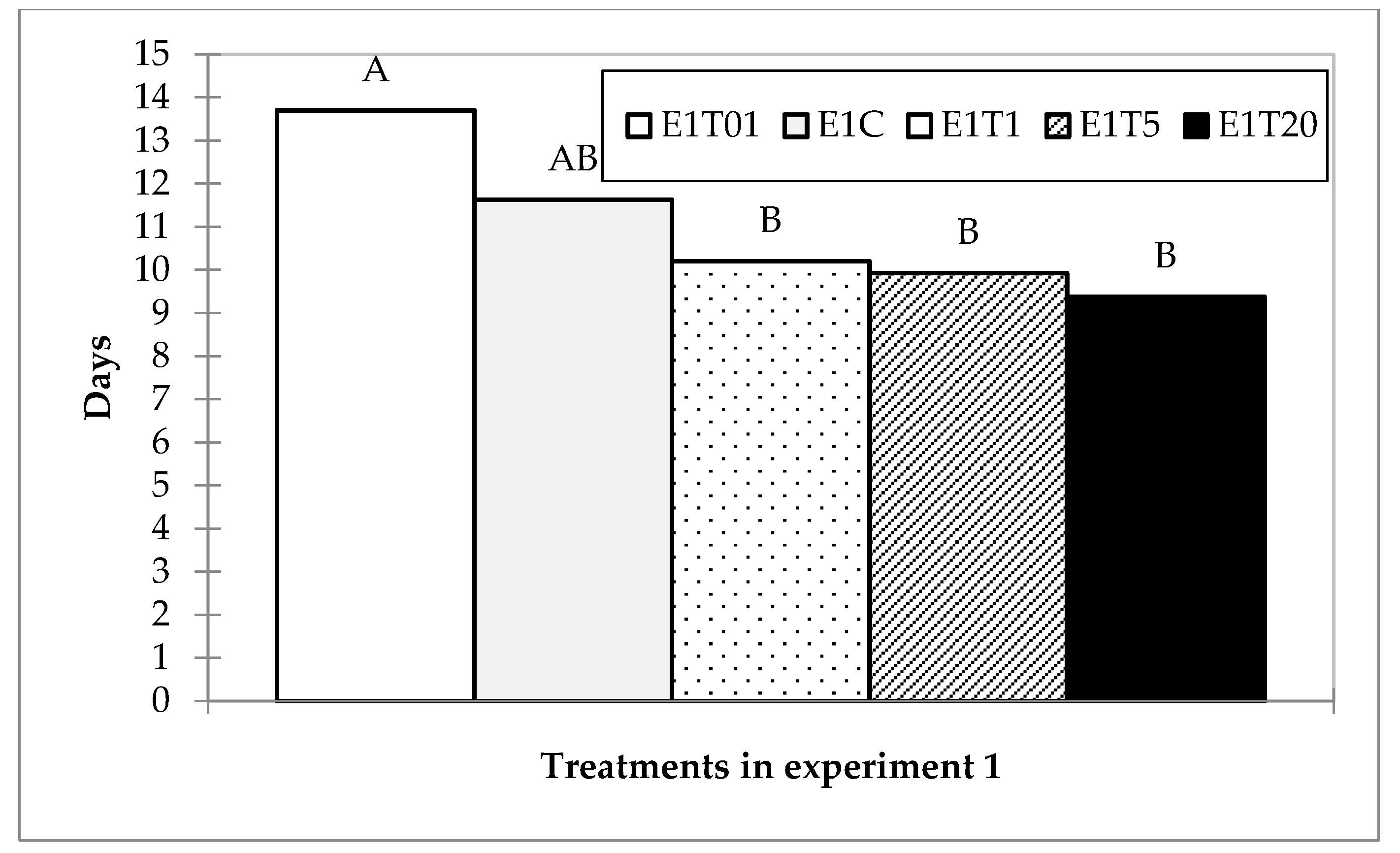
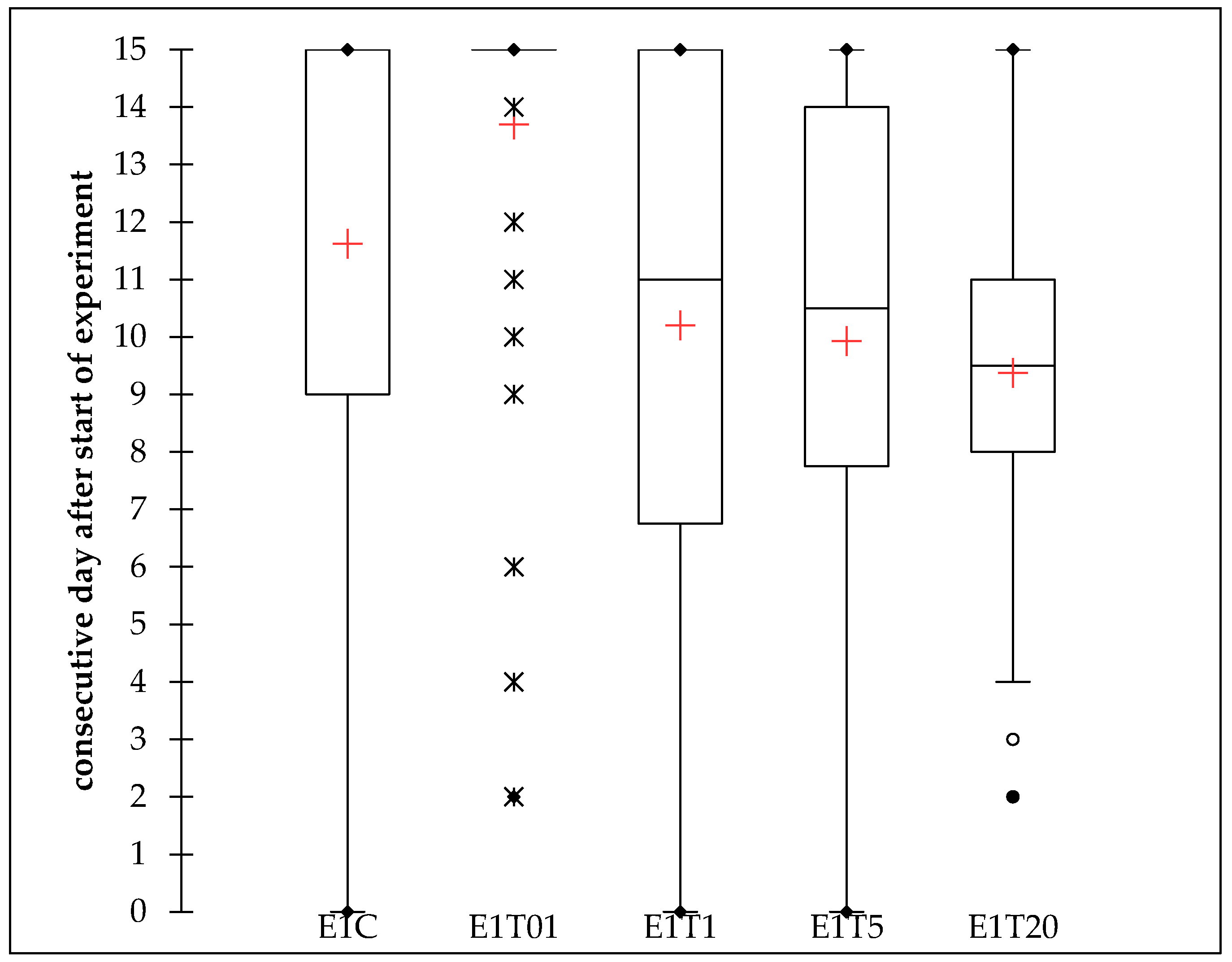
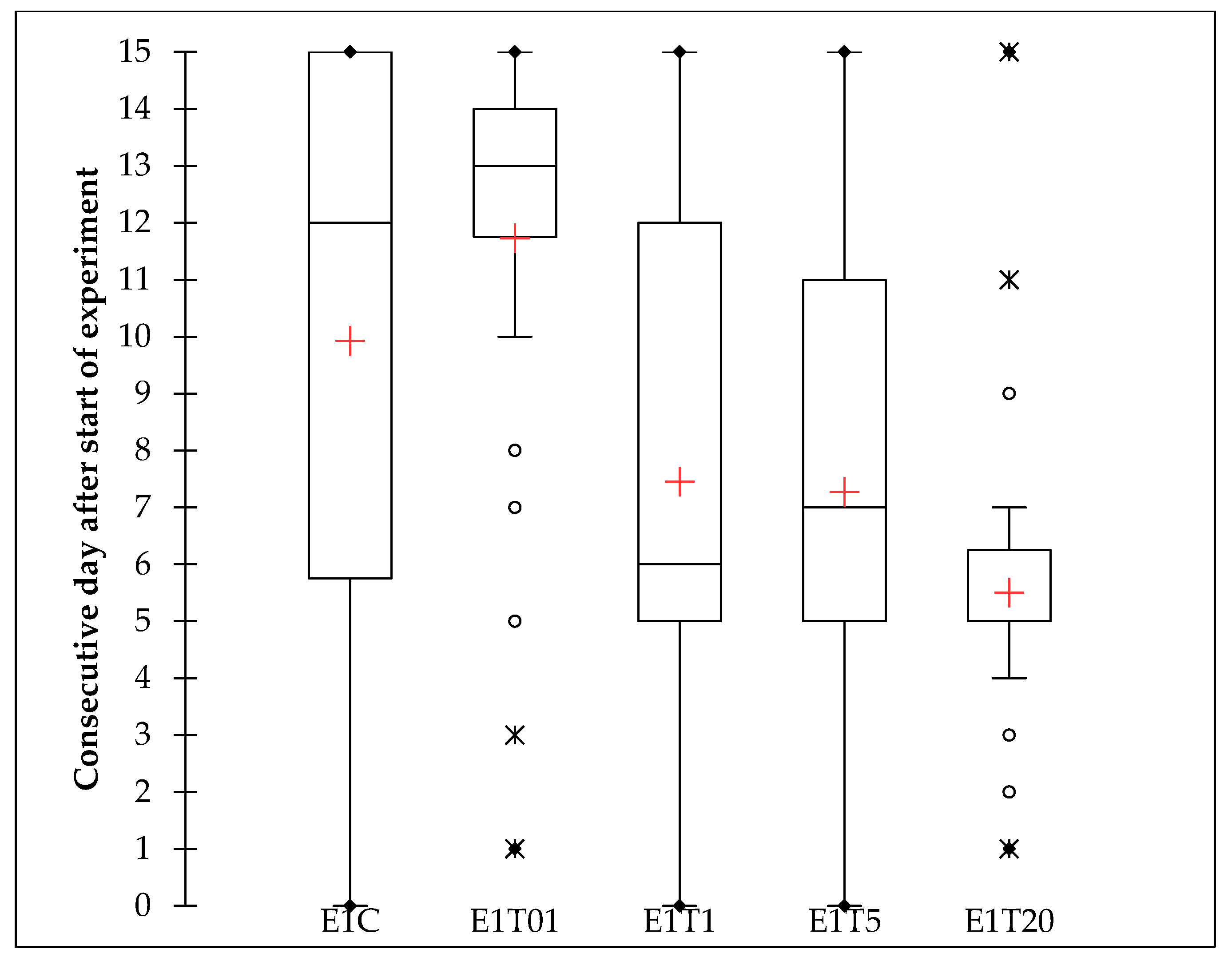
3.1.2. Lethal Time
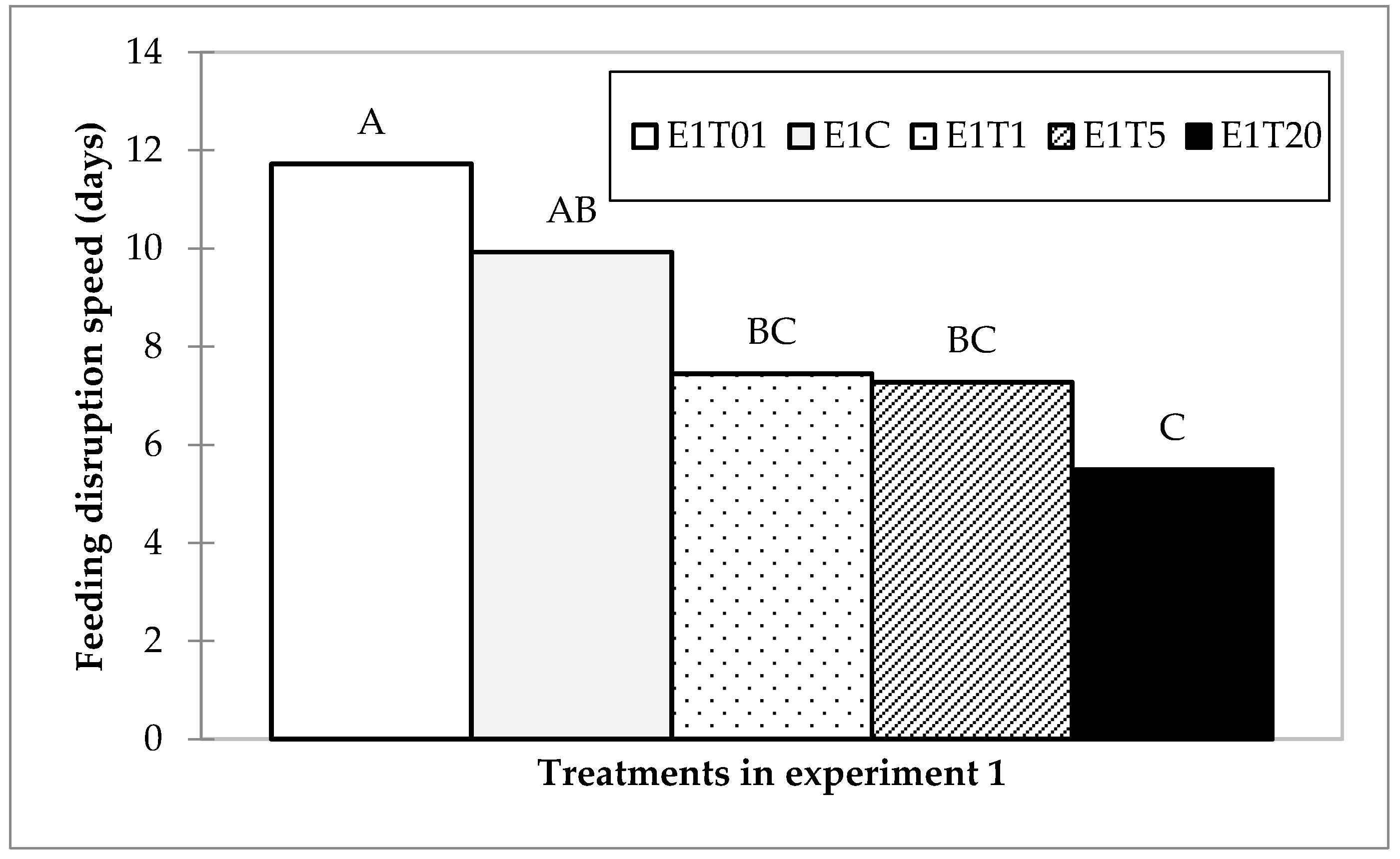
3.1.3. Feeding Disruption Speed
3.2. Experiment 2
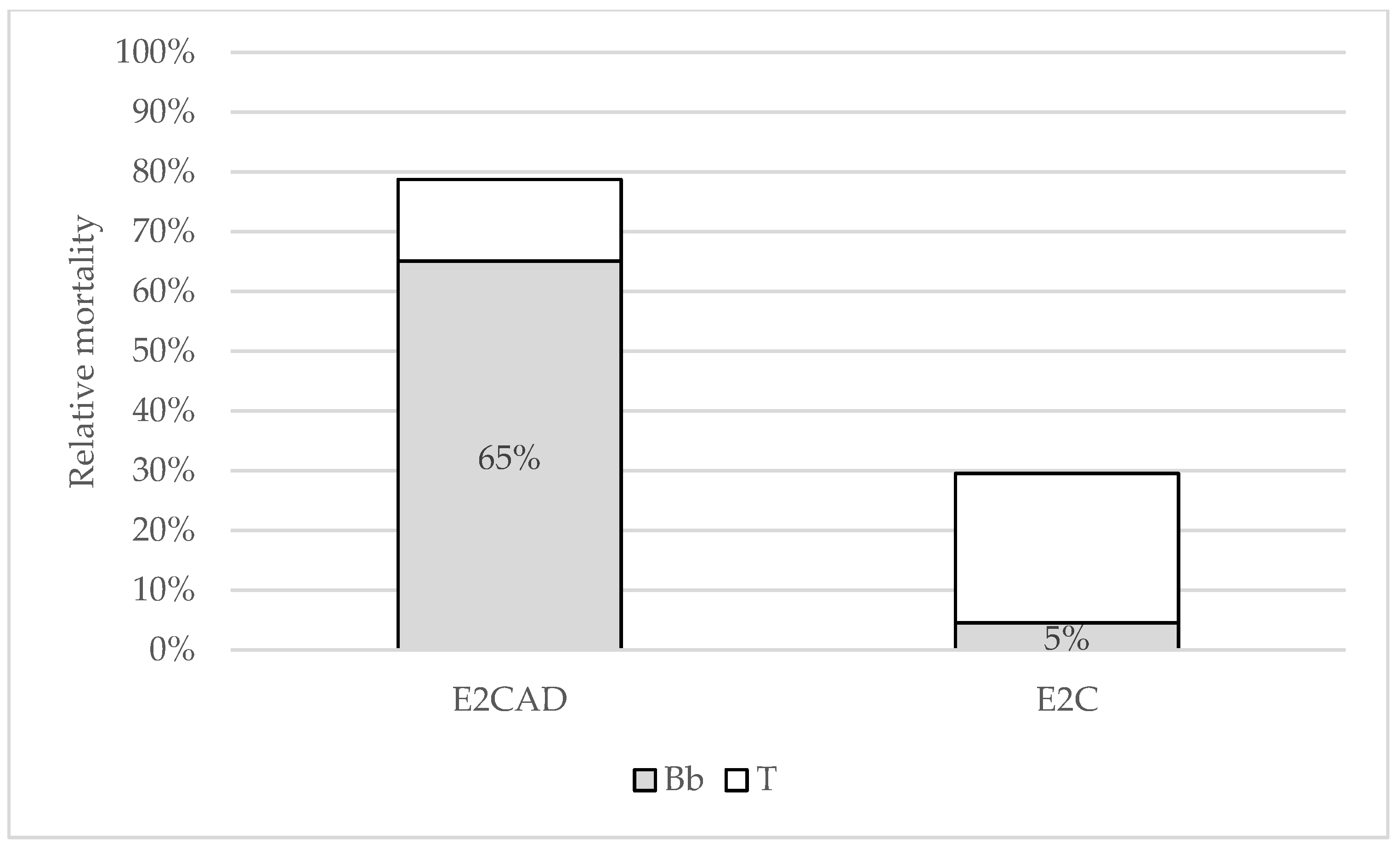
Mortality Rates
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skrzecz, I.; Ślusarski, S.; Tkaczyk, M. Integration of science and practice for Dendrolimus pini (L.) management—A review with special reference to Central Europe. For. Ecol. Manag. 2020, 455, 117697. [Google Scholar] [CrossRef]
- Ray, D.; Peace, A.; Moore, R.; Petr, M.; Grieve, Y.; Convery, C.; Ziesche, T. Improved prediction of the climate-driven outbreaks of Dendrolimus pini in Pinus sylvestris forests. Forestry 2016, 89, 230–244. [Google Scholar] [CrossRef] [Green Version]
- Björkman, C.; Lindelöw, A.; Eklund, K.; Kyrk, S.; Klapwijk, M.J.; Fedderwitz, F.; Nordlander, G. A rare event—An isolated outbreak of the pine-tree lappet moth (Dendrolimus pini) in the Stockholm archipelago. Entomol. Tidskr. 2013, 134, 1–9. [Google Scholar]
- Moore, D.; Reed, M.; Le Patourel, G.; Abraham, Y.J.; Prior, C. Reduction of feeding by the desert locust, Schistocerca gregaria, after infection with Metarhizium flavoviride. J. Invertebr. Pathol. 1992, 60, 304–307. [Google Scholar] [CrossRef]
- Matek, M.; Pernek, M. First record of Dendrolimus pini outbreak on Aleppo Pine in Croatia and severe case of population collapse caused by entomopathogen Beauveria bassiana. South East Eur. For. 2018, 9, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Lesniak, A. Climatic and meteorological conditions of the pine moth (Dendrolimus pini L.) outbreaks. Ekol. Polska 1976, 24, 515–547. [Google Scholar]
- Sierpinska, A. Towards an integrated management of Dendrolimus pini L. In Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects; McManus, M.L., Liebhold, A.M., Eds.; U.S. Department of Agriculture: Radnor, PA, USA, 1998; pp. 129–142. [Google Scholar]
- Altenkirch, W.; Majunke, C.; Ohnesorge, B. Waldschutz auf ökologischer Grundlage; Ulmer: Stuttgart, Germany, 2002; p. 434. [Google Scholar]
- Habermann, M. Auswirkungen der Anwendungsbestimmungen für die Ausbringung von Pflanzenschutzmitteln mit Luftfahrzeugen im Wald. J. Kult. 2017, 69, 249–254. [Google Scholar] [CrossRef]
- Sukovata, L.; Asztemborska, M.; Rudziński, K.J.; Cieślak, M.; Staszek, D.; Janiszewski, W.; Szmigielski, R.; Kolk, A.; Raczko, J. Effect of dispenser type, trap design and placement on catches of pine-tree lappet moth, Dendrolimus pini. Phytoparasitica 2020, 48, 63–74. [Google Scholar] [CrossRef]
- Sukovata, L.; Jaworski, T. The abundance of the nun moth and lappet moth larvae on trees of different trunk thickness in Scots pine stands in the Notec forest complex. For. Res. Pap. 2010, 71, 231–237. [Google Scholar] [CrossRef]
- Tumialis, D.; Skrzecz, I.; Mazurkiewicz, A.; Pezowicz, E. Sensitivity of caterpillars of the pine tree lappet moth Dendrolimus pini to native isolates of entomopathogenic nematodes. Int. J. Pest. Manag. 2018, 1–6. [Google Scholar] [CrossRef]
- Gedminas, A.; Lynikienė, J.; Ρovilaitienė, A. Entomopathogenic fungus Cordyceps militaris. Distribution in south Lithuania,‘in vitro’ cultivation and pathogenicity tests. Balt. For. 2015, 21, 359–368. [Google Scholar]
- Batta, Y.A. Biocontrol of almond bark beetle (Scolytus amygdali Geurin-Meneville, Coleoptera: Scolytidae) using Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: Hyphomycetes). J. Appl. Microbiol. 2007, 103, 1406–1414. [Google Scholar] [CrossRef]
- Khosravi, R.; Sendi, J.J.; Zibaee, A.; Shokrgozar, M.A. Virulence of four Beauveria bassiana (Balsamo) (Asc., Hypocreales) isolates on rose sawfly, Arge rosae under laboratory condition. J. King Saud Univ. Sci. 2015, 27, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, T.; Takken, W.; Koenraadt, C.J.M. Development of Metarhizium anisopliae and Beauveria bassiana formulations for control of malaria mosquito larvae. Parasites Vectors 2011, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.Z. List on the insect hosts of Beauveria bassiana. In Study and Application of Entomogenous Fungi in China; Academic Periodical Press: Beijing, China, 1988; Volume 1, pp. 241–255, (In Chinese with Latin names and English summary). [Google Scholar]
- Feng, M.G.; Poprawski, T.J.; Khachatourians, G.G. Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: Current status. Biocontrol Sci. Technol. 1994, 4, 3–34. [Google Scholar] [CrossRef]
- Zhang, L.W.; Liu, Y.J.; Yao, J.; Wang, B.; Huang, B.; Li, Z.Z.; Sun, J.H. Evaluation of Beauveria bassiana (Hyphomycetes) isolates as potential agents for control of Dendroctonus valens. Insect Sci. 2011, 18, 209–216. [Google Scholar] [CrossRef]
- Mc Namara, L.; Kapranas, A.; Williams, C.D.; O’Tuama, P.; Kavanagh, K.; Griffin, C.T. Efficacy of entomopathogenic fungi against large pine weevil, Hylobius abietis, and their additive effects when combined with entomopathogenic nematodes. J. Pest. Sci. 2018, 91, 1407–1419. [Google Scholar] [CrossRef]
- Singh, D.; Raina, T.K.; Singh, J. Entomopathogenic fungi: An effective biocontrol agent for management of insect populations naturally. J. Pharm. Sci. Res. 2017, 9, 830–839. [Google Scholar]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Valero-Jiménez, C.A.; Wiegers, H.; Zwaan, B.J.; Koenraadt, C.J.M.; Van Kan, J.A.L. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2016, 133, 41–49. [Google Scholar] [CrossRef]
- Kaulfuß, S. The Monitoring and Prognosis of Insect Pests: Early Recognition, Monitoring, Analysis and Action. Wald Wissen. Net, Informationen für die Fortspraxis -Forest Pest Handbook. 2012. Available online: http://www.waldwissen.net (accessed on 22 July 2020).
- Duffus, J.H.; Nordberg, M.; Templeton, D.M. Glossary of terms used in toxicology, 2nd Edition (IUPAC recommendations 2007). Pure Appl. Chem. 2007, 79, 1153–1344. [Google Scholar] [CrossRef] [Green Version]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Tefera, T.; Pringle, K.L. Food consumption by Chilo partellus (Lepidoptera: Pyralidae) larvae infected with Beauveria bassiana and Metarhizium anisopliae and effects of feeding natural versus artificial diets on mortality and mycosis. J. Invertebr. Pathol. 2003, 84, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Scholte, E.J.; Knols, B.G.J.; Takken, W. Infection of the malaria mosquito Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebr. Pathol. 2006, 91, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Blanford, S.; Shi, W.; Christian, R.; Marden, J.H.; Koekemoer, L.L.; Brooke, B.D.; Coetzee, M.; Read, A.F.; Thomas, M.B. Lethal and pre-lethal effects of a fungal biopesticide contribute to substantial and rapid control of malaria vectors. PLoS ONE 2011, 6, e23591. [Google Scholar] [CrossRef] [Green Version]
- Roditakis, E.; Couzin, I.D.; Franks, N.R.; Charnley, A.K. Effects of Lecanicillium longisporum infection on the behaviour of the green peach aphid Myzus persicae. J. Insect Physiol. 2008, 54, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.B.; Blanford, S.; Lomer, C.J. Reduction of feeding by the variegated grasshopper, Zonocerus variegatus, following infection by the fungal pathogen, Metarhizium flavoviride. Biocontrol Sci. Technol. 2010, 7, 327–334. [Google Scholar] [CrossRef]
- Conceschi, M.R.; D’Alessandro, C.P.; de Andrade Moral, R.; Demétrio, C.G.B.; Júnior, I.D. Transmission potential of the entomopathogenic fungi Isaria fumosorosea and Beauveria bassiana from sporulated cadavers of Diaphorina citri and Toxoptera citricida to uninfected D. citri adults. Biocontrol 2016, 61, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Kreutz, J.; Zimmermann, G.; Vaupel, O. Horizontal transmission of the entomopathogenic fungus Beauveria bassiana among the spruce bark beetle, Ips typographus (Col., Scolytidae) in the laboratory and under field conditions. Biocontrol Sci. Technol. 2004, 14, 837–848. [Google Scholar] [CrossRef]
- Drummond, F.A.; Groden, E. IV. Insect Pests and Natural Enemies. In The Ecology, Economics, and Management of Potato Cropping Systems: A Report of the First Four Years of the Maine Potato Ecosystem Project; Marra, M.C., Harrity, B.A., Eds.; University of Maine: Maine, ME, USA, 1996; pp. 80–118. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | E1T1 | E1T5 | E1T20 | E1T01 | E1C |
|---|---|---|---|---|---|
| Treatment | Spore suspension | Spore suspension | Spore suspension | Spore suspension | H₂O + Tween (0.1%) |
| Dose per larvae [µL] | 1 | 5 | 20 | 1 | 20 |
| Spore suspension concentration [spores/µL] | 7.9 × 104 | 7.9 × 104 | 7.9 × 104 | 7.9 × 102 (dilution 1:100) | - |
| Group | E2CAD | E2C |
|---|---|---|
| Treatment | Healthy larvae + cadaver | Only healthy larvae |
| Treatment | E1T1 | E1T5 | E1T20 | E1T01 | E1C | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mortality rate | Abs | Rel | Abs | Rel | Abs | Rel | Abs | Rel | Abs | Rel |
| Total | 24 | 54% | 34 | 83% | 40 | 100% | 9 | 11% | 5 | 13% |
| χ2 (p) | 6.40 (0.0114) | 0.90 (0.3428) | 0.00 (1.0000) | 24.03 (<0.0001) | 30.63 (<0.0001) | |||||
| B. bassiana | 22 | 55% | 25 | 63% | 35 | 88% | 7 | 18% | 0 | 0% |
| χ2 (p) | 8.10 (0.0044) | 5.63 (0.0177) | 0.63 (0.4292) | 27.23 (<0.0001) | 40.00 (0.0000) | |||||
| Treatment | N | Min | Max | Sd | ANOVA | Kruskal–Wallis | |||
|---|---|---|---|---|---|---|---|---|---|
| E1C | 40 | 0.000 | 15.000 | 11.625 | 5.092 | DF | 4 | K (Observed value) | 40.230 |
| E1T01 | 40 | 2.000 | 1.000 | 13.700 | 3.164 | Sum of squares | 484.430 | K (Critical value) | 9.488 |
| E1T1 | 40 | 0.000 | 15.000 | 10.200 | 4.794 | Mean squares | 121.108 | DF | 4 |
| E1T20 | 40 | 2.000 | 15.000 | 9.375 | 2.862 | F | 6.901 | p-value (one-tailed) | <0.0001 |
| Pr > F | <0.0001 | ||||||||
| Treatment | E2CAD | E2C | ||
|---|---|---|---|---|
| Mortality | Abs | Rel | Abs | Rel |
| Total | 51 | 85% | 13 | 30% |
| χ2 (p) | 1.35 (0.2453) | 21.84 (<0.0001) | ||
| B. bassiana | 40 | 67% | 2 | 5% |
| χ2 (p) | 6.67 (0.0098) | 40.09 (<0.0001) | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, M.; Lacković, N.; Pernek, M. Effect of Beauveria bassiana Fungal Infection on Survival and Feeding Behavior of Pine-Tree Lappet Moth (Dendrolimus pini L.). Forests 2020, 11, 974. https://doi.org/10.3390/f11090974
Kovač M, Lacković N, Pernek M. Effect of Beauveria bassiana Fungal Infection on Survival and Feeding Behavior of Pine-Tree Lappet Moth (Dendrolimus pini L.). Forests. 2020; 11(9):974. https://doi.org/10.3390/f11090974
Chicago/Turabian StyleKovač, Marta, Nikola Lacković, and Milan Pernek. 2020. "Effect of Beauveria bassiana Fungal Infection on Survival and Feeding Behavior of Pine-Tree Lappet Moth (Dendrolimus pini L.)" Forests 11, no. 9: 974. https://doi.org/10.3390/f11090974
APA StyleKovač, M., Lacković, N., & Pernek, M. (2020). Effect of Beauveria bassiana Fungal Infection on Survival and Feeding Behavior of Pine-Tree Lappet Moth (Dendrolimus pini L.). Forests, 11(9), 974. https://doi.org/10.3390/f11090974