Dynamics of Carbon Accumulation in Tropical Dry Forests under Climate Change Extremes

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
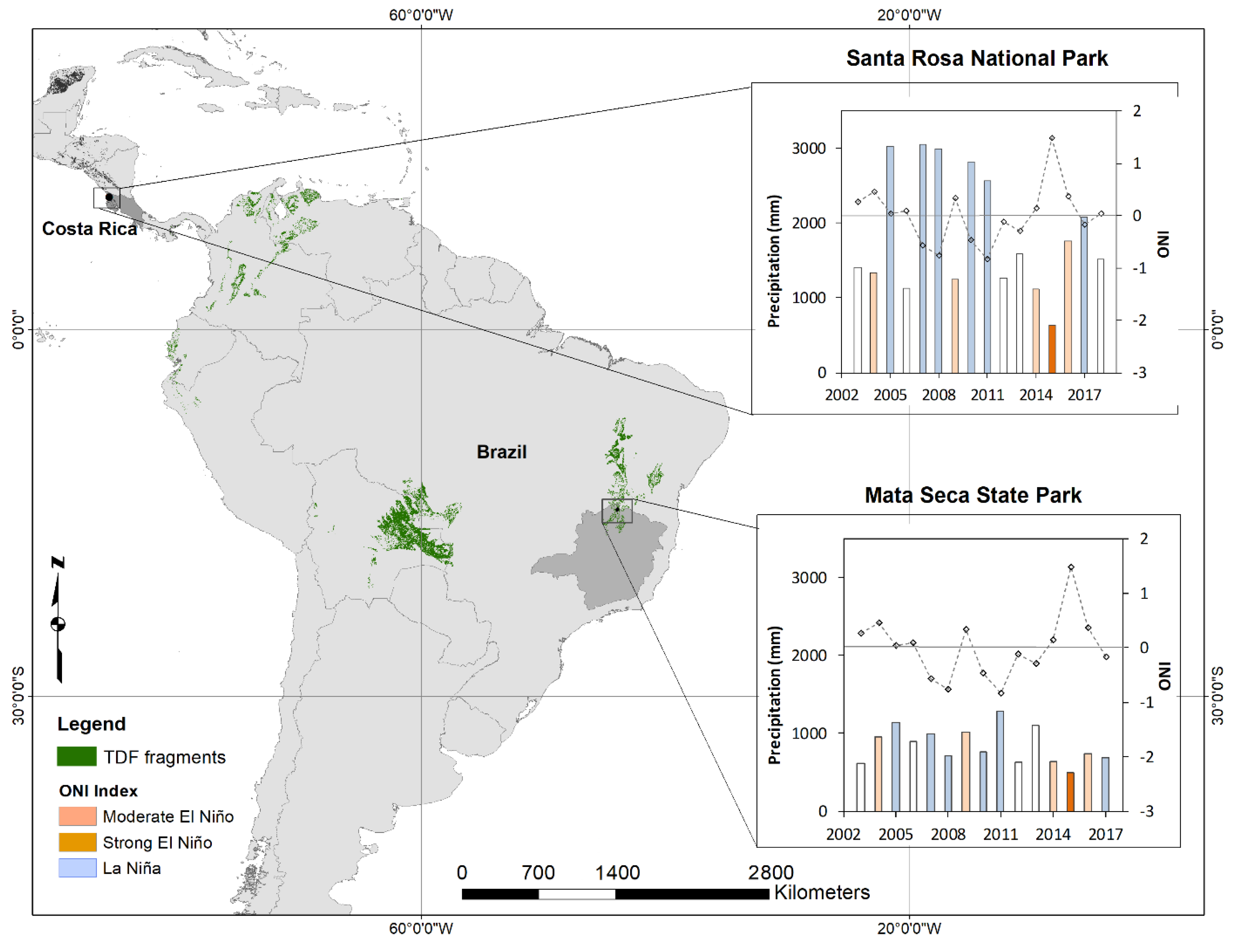
2.1. Study Site
2.2. Climatic Variables
2.3. Forest Inventory Plots
2.4. Forest Dynamics and Carbon Accumulation
2.5. Litterfall Data
2.6. Analysis
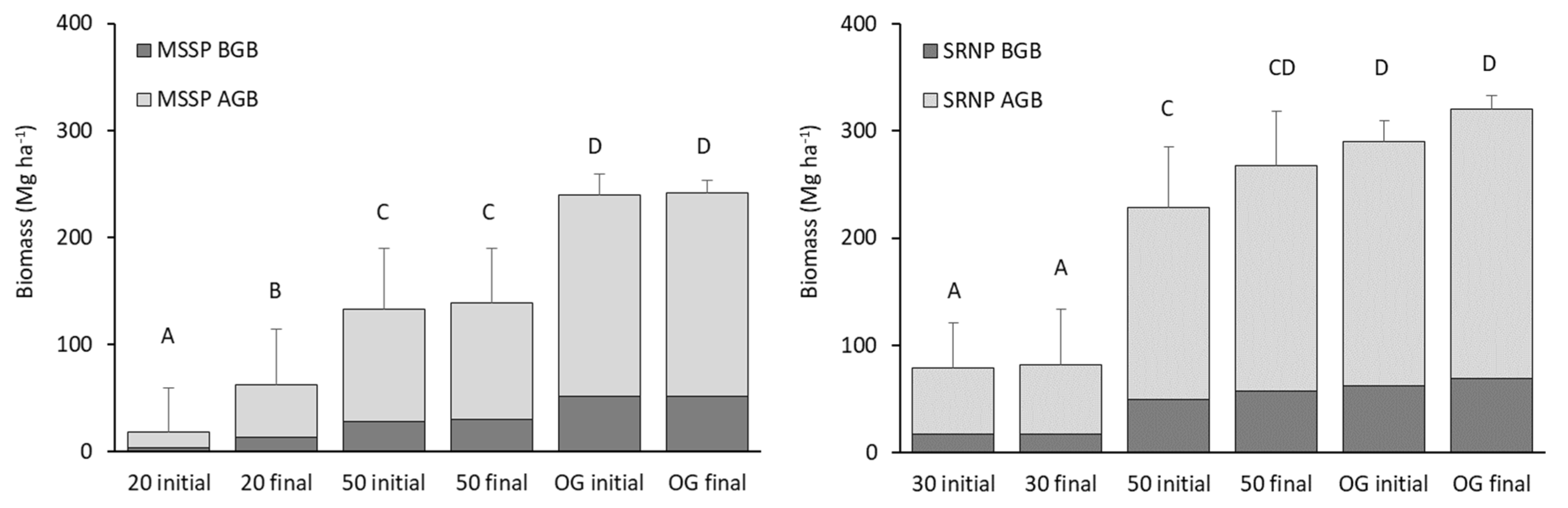
3. Results
3.1. Biomass and Carbon Dynamics
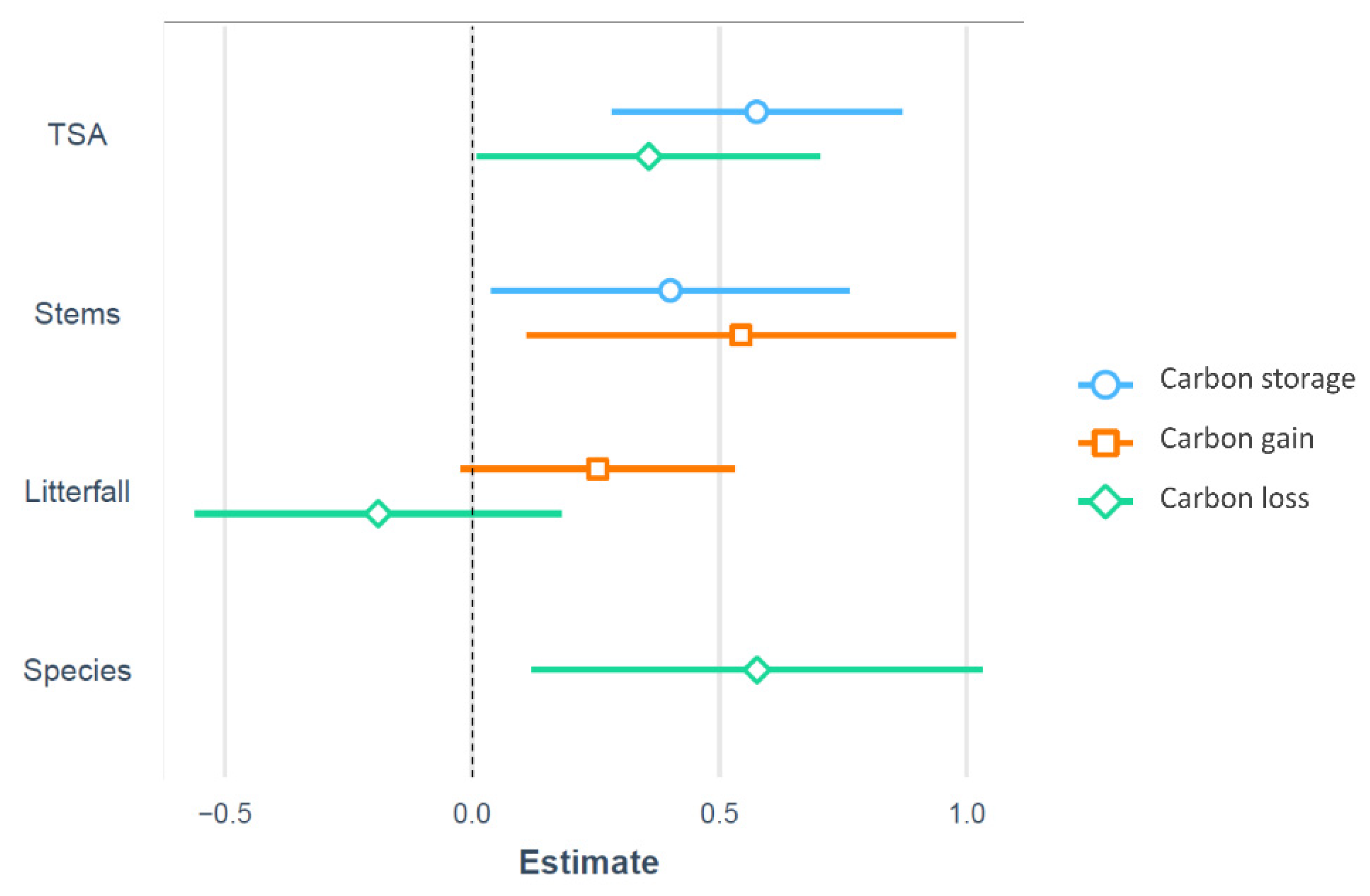
3.2. Relationship of Forest Structure and Composition on Carbon Dynamics
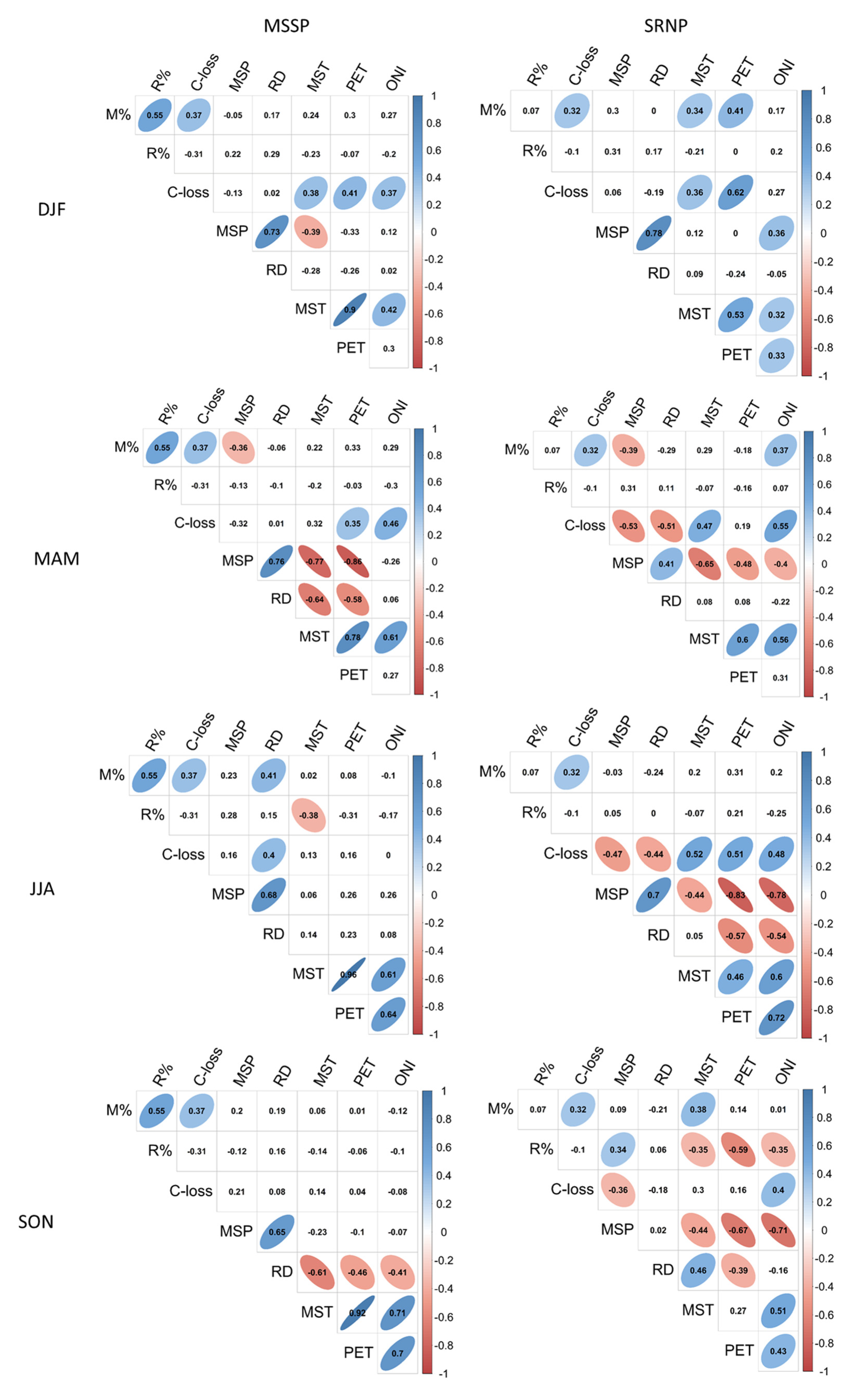
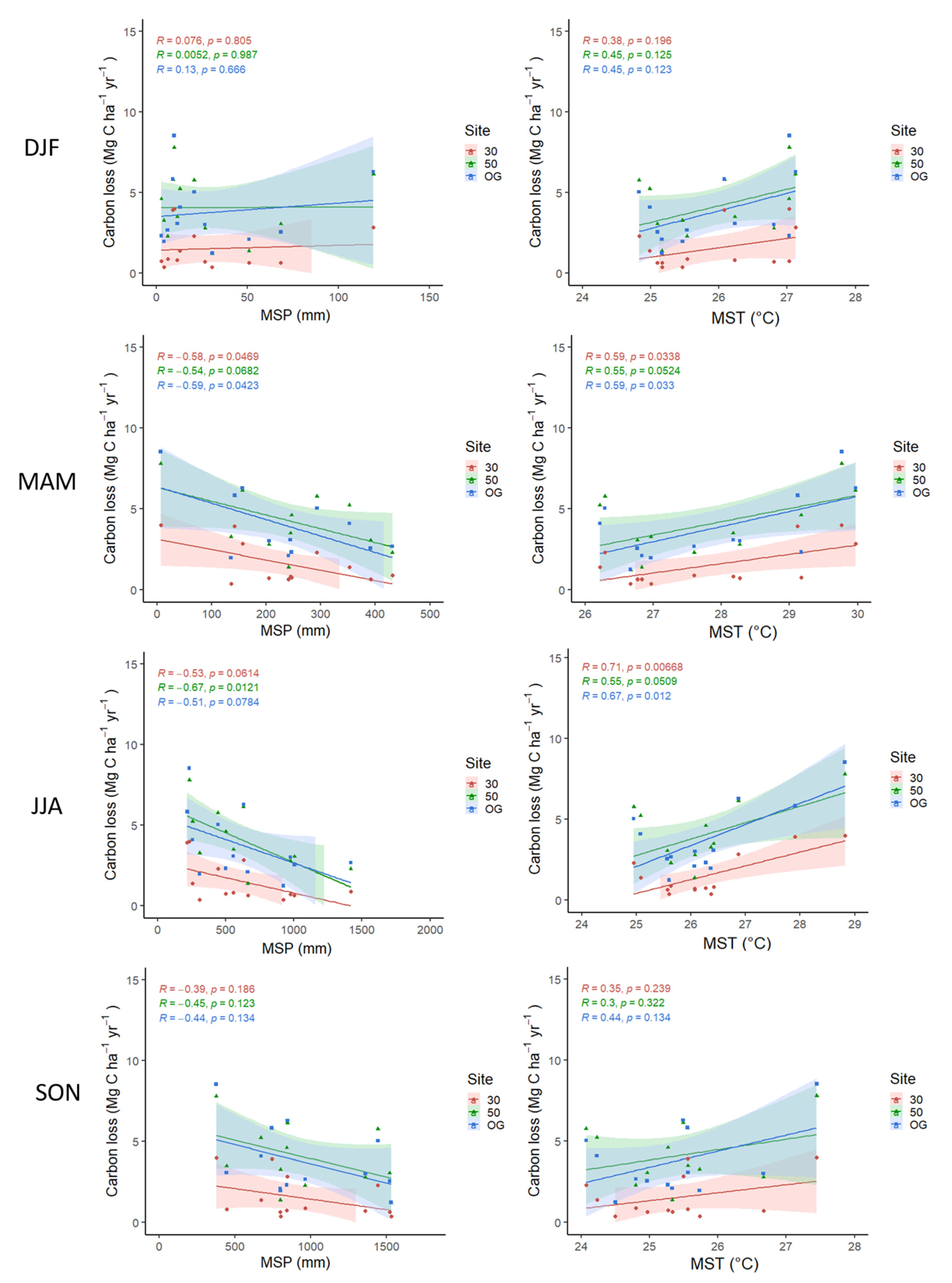
3.3. Relationship of Climate Variability on Carbon Dynamics
4. Discussion
4.1. Forests Biomass and Carbon Sequestration Dynamics
4.2. Effects of Climate Variability on Carbon Sequestration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | t Value | Pr(>|t|) | Model | ||
|---|---|---|---|---|---|---|
| (Intercept) | −0.18 | 0.13 | 0.00 | R2 = 0.58 | ||
| TSA | 0.35 | 0.16 | 2.28 | <0.05 | p < 0.001 | |
| Species | 0.57 | 0.15 | 3.87 | <0.001 | ||
| Litterfall | −0.19 | 0.14 | −1.39 | |||
| carbon gain | (Intercept) | −0.00 | 0.16 | 0.00 | R2 = 0.33 | |
| Stems | 0.54 | 0.16 | 3.38 | <0.05 | p < 0.05 | |
| Litterfall | 2.54 | 0.16 | 1.58 | |||
| carbon storage | (Intercept) | −0.00 | 0.08 | 0.00 | R2 = 0.79 | |
| TSA | 0.57 | 0.12 | 4.67 | <0.001 | p < 0.001 | |
| Stems | 0.40 | 0.12 | 3.26 | <0.05 |
References
- Portillo-Quintero, C.; Sanchez-Azofeifa, A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Ruesch, A.S.; Achard, F.; Clayton, M.K.; Holmgren, P.; Ramankutty, N.; Foley, J.A. Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Proc. Natl. Acad. Sci. USA 2010, 107, 16732–16737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, L.; Bongers, F.; Aide, T.M.; Zambrano, A.M.A.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.S.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass resilience of Neotropical secondary forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.H. Tropical dry forests. In Biodiversity; National Academy Press: Washington, DC, USA, 1988; pp. 130–137. [Google Scholar]
- Portillo-Quintero, C.; Sanchez-Azofeifa, A.; Calvo-Alvarado, J.; Quesada, M.; do Espirito Santo, M.M. The role of tropical dry forests for biodiversity, carbon and water conservation in the neotropics: Lessons learned and opportunities for its sustainable management. Reg. Environ. Chang. 2015, 15, 1039–1049. [Google Scholar] [CrossRef]
- Calvo-Rodriguez, S.; Sanchez-Azofeifa, A.; Duran, S.M.; Espírito-Santo, M.M. Assessing ecosystem services in Neotropical dry forests: A systematic review. Environ. Conserv. 2017, 44, 34–43. [Google Scholar] [CrossRef]
- Jaramillo, V.J.; Kauffman, J.B.; Rentería-Rodríguez, L.; Cummings, D.L.; Ellingson, L.J. Biomass, Carbon, and Nitrogen Pools in Mexican Tropical Dry Forest Landscapes. Ecosystems 2003, 6, 609–629. [Google Scholar] [CrossRef]
- Mora, F.; Jaramillo, V.J.; Bhaskar, R.; Gavito, M.; Siddique, I.; Byrnes, J.E.K.; Balvanera, P. Carbon Accumulation in Neotropical Dry Secondary Forests: The Roles of Forest Age and Tree Dominance and Diversity. Ecosystems 2018, 21, 536–550. [Google Scholar] [CrossRef]
- Gavito, M.E.; Sandoval-Pérez, A.L.; del Castillo, K.; Cohen-Salgado, D.; Colarte-Avilés, M.E.; Mora, F.; Santibáñez-Rentería, A.; Siddique, I.; Urquijo-Ramos, C. Resilience of soil nutrient availability and organic matter decomposition to hurricane impact in a tropical dry forest ecosystem. For. Ecol. Manage. 2018, 426, 81–90. [Google Scholar] [CrossRef]
- Becknell, J.M.; Kucek, L.K.; Powers, J.S. Aboveground biomass in mature and secondary seasonally dry tropical forests: A literature review and global synthesis. For. Ecol. Manag. 2012, 276, 88–95. [Google Scholar] [CrossRef]
- Jaramillo, V.J.; Martínez-Yrízar, A.; Sanford, R.L. Primary Productivity and Biogeochemistry of Seasonally Dry Tropical Forests. In Seasonally Dry Tropical Forests; Island Press: Washington, DC, USA, 2011; pp. 109–128. [Google Scholar] [CrossRef]
- Castro-Magnani, M.; Sanchez-Azofeifa, A.; Metternicht, G.; Laakso, K. Integration of remote-sensing based metrics and econometric models to assess the socio-economic contributions of carbon sequestration in unmanaged tropical dry forests. Environ. Sustain. Indic. 2021, 100100. [Google Scholar] [CrossRef]
- Pennington, R.T.; Lavin, M.; Prado, D.E.; Pendry, C.A.; Pell, S.K.; Butterworth, C.A. Historical climate change and speciation: Neotropical seasonally dry forest plants show patterns of both Tertiary and Quaternary diversification. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 515–538. [Google Scholar] [CrossRef] [PubMed]
- Pennington, R.T.; Lavin, M.; De Oliveira-Filho, A.T. Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Lasky, J.R.; Uriarte, M.; Muscarella, R. Synchrony, compensatory dynamics, and the functional trait basis of phenological diversity in a tropical dry forest tree community: Effects of rainfall seasonality. Environ. Res. Lett. 2016, 11, 115003. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, art129. [Google Scholar] [CrossRef]
- Castro, S.M.; Sanchez-Azofeifa, G.A.; Sato, H. Environmental Research Letters Effect of drought on productivity in a Costa Rican tropical dry forest Effect of drought on productivity in a Costa Rican tropical dry forest. Environ. Res. Lett. 2018, 13, 045001. [Google Scholar] [CrossRef]
- Cai, W.; Borlace, S.; Lengaigne, M.; Van Rensch, P.; Collins, M.; Vecchi, G.A.; Timmermann, A.; Santoso, A.; McPhaden, M.J.; Wu, L.; et al. Increasing frequency of extreme El Niño events due to greenhouse warming. Nat. Clim. Chang. 2014, 4, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Steinhoff, D.F.; Monaghan, A.J.; Clark, M.P. Projected impact of twenty-first century ENSO changes on rainfall over Central America and Northwest South America from CMIP5 AOGCMs. Clim. Dyn. 2015, 44, 1329–1349. [Google Scholar] [CrossRef]
- Wang, G.; Cai, W.; Gan, B.; Wu, L.; Santoso, A.; Lin, X.; Chen, Z.; McPhaden, M.J. Continued increase of extreme El Niño frequency long after 1.5 °C warming stabilization. Nat. Clim. Chang. 2017, 7, 568–572. [Google Scholar] [CrossRef]
- Kim, J.S.; Kug, J.S.; Jeong, S.J. Intensification of terrestrial carbon cycle related to El Nino-Southern Oscillation under greenhouse warming. Nat. Commun. 2017, 8, 1674. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.; Dupuy, J.M.; Gei, M.G.; Hulshof, C.; Medvigy, D.; Pizano, C.; Salgado-Negret, B.; Smith, C.M.; Trierweiler, A.; van Bloem, S.J. Will seasonally dry tropical forests be sensitive or resistant to future changes in rainfall regimes? Environ. Res. Lett. 2017, 12, 23001. [Google Scholar] [CrossRef] [Green Version]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C. Widespread Increase of Tree Mortality Rates in the Western United States. Science 2009, 323, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.; Lugo, A.E. The Storage and Production of Organic Matter in Tropical Forests and Their Role in the Global Carbon Cycle. Biotropica 1982, 14, 161. [Google Scholar] [CrossRef]
- Campo, J.; Vázquez-Yanes, C. Effects of nutrient limitation on aboveground carbon dynamics during tropical dry forest regeneration in Yucatán, Mexico. Ecosystems 2004, 7, 311–319. [Google Scholar] [CrossRef]
- Read, L.; Lawrence, D. Recovery of biomass following shifting cultivation in dry tropical forests of the Yucatan. Ecol. Appl. 2003, 13, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Aryal, D.R.; De Jong, B.H.J.; Ochoa-Gaona, S.; Mendoza-Vega, J.; Esparza-Olguin, L. Successional and seasonal variation in litterfall and associated nutrient transfer in semi-evergreen tropical forests of SE Mexico. Nutr. Cycl. Agroecosyst. 2015, 103, 45–60. [Google Scholar] [CrossRef]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, CA, USA, 1967. [Google Scholar]
- Sánchez-Azofeifa, A.; Calvo-Alvarado, J.; Espírito-Santo, M.M.; Fernandes, G.W.; Powers, J.S.; Quesada, M. Tropical dry forest in the Americas: The tropi-dry endeavour. In Tropical Dry Forests Americas Ecology. Ecology, Conservation, and Management; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–16. [Google Scholar]
- Madeira, B.G.; Espírito-Santo, M.M.; Neto, S.D.; Nunes, Y.R.F.; Sanchez-Azofeifa, A.; Fernandes, G.W.; Quesada, M. Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. For. Ecol. Recent Adv. Plant Ecol. 2009, 201, 291–304. [Google Scholar] [CrossRef]
- Kalacska, M.; Sanchez-Azofeifa, G.; Calvo-Alvarado, J.; Quesada, M.; Rivard, B.; Janzen, D.; Sanchez-Azofeifa, A. Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest. For. Ecol. Manag. 2004, 200, 227–247. [Google Scholar] [CrossRef]
- Souza, S.R.; Veloso, M.D.M.; Espírito-Santo, M.M.; Silva, J.O.; Sánchez-Azofeifa, A.; Brito, B.G.S.E.; Fernandes, G.W. Litterfall dynamics along a successional gradient in a Brazilian tropical dry forest. For. Ecosyst. 2019, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, D.; Lieberman, M. Forest tree growth and dynamics at La Selva, Costa Rica (1969–1982). J. Trop. Ecol. 1987, 3, 347–358. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Manokaran, N.; LaFrankie, J.V.; Hubbell, S.P.; Foster, R.B. Dynamics of the forest communities at Pasoh and Barro Colorado: Comparing two 50–ha plots. Philos. Trans. R. Soc. B Biol. Sci. 1999, 354, 1739–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.S.; Tiffin, P. Plant functional type classifications in tropical dry forests in Costa Rica: Leaf habit versus taxonomic approaches. Funct. Ecol. 2010, 24, 927–936. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.R.; Thomas, S.C. A Reassessment of Carbon Content in Tropical Trees. PLoS ONE 2011, 6, e23533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Heijden, G.M.F.; Powers, J.S.; Schnitzer, S.A. Lianas reduce carbon accumulation and storage in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 13267–13271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- 2006 IPCC Guidelines for National Greenhouse Gas Inventories—Google Scholar. Available online: https://scholar-google-ca.login.ezproxy.library.ualberta.ca/scholar?hl=en&as_sdt=0%2C5&q=2006+IPCC+Guidelines+for+National+Greenhouse+Gas+Inventories&btnG= (accessed on 14 December 2020).
- Rozendaal, D.M.A.; Chazdon, R.L.; Arreola-Villa, F.; Balvanera, P.; Bentos, T.V.; Dupuy, J.M.; Hernández-Stefanoni, J.L.; Jakovac, C.C.; Lebrija-Trejos, E.E.; Lohbeck, M.; et al. Demographic Drivers of Aboveground Biomass Dynamics during Secondary Succession in Neotropical Dry and Wet Forests. Ecosystems 2017, 20, 340–353. [Google Scholar] [CrossRef]
- Chapin, F.; Eviner, V.T. Biogeochemical Interactions Governing Terrestrial Net Primary Production. Treatise Geochem. 2013, 10, 189–216. [Google Scholar] [CrossRef]
- Martinez-Yrizar, A.; Maass, J.M.; Perez-Jimenez, L.A.; Sarukhan, J. Net primary productivity of a tropical deciduous forest ecosystem in western Mexico. J. Trop. Ecol. 1996, 12, 169–175. [Google Scholar] [CrossRef]
- Cao, S.; Sanchez-Azofeifa, A.; Duran, S.; Calvo-Rodriguez, S. Estimation of aboveground net primary productivity in secondary tropical dry forests using the Carnegie-Ames-Stanford approach (CASA) model. Environ. Res. Lett. 2016, 11, 075004. [Google Scholar] [CrossRef]
- Dupuy, J.M.; Hernández-Stefanoni, J.L.; Hernández-Juárez, R.A.; Tetetla-Rangel, E.; López-Martínez, J.O.; Leyequién-Abarca, E.; Tun-Dzul, F.J.; May-Pat, F. Patterns and Correlates of Tropical Dry Forest Structure and Composition in a Highly Replicated Chronosequence in Yucatan, Mexico. Biotropica 2012, 44, 151–162. [Google Scholar] [CrossRef]
- Becknell, J.M.; Powers, J.S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 2014, 44, 604–613. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Brenes, A.R.; Alvarado, B.V. Erratum: Effects of climate and stand age on annual tree dynamics in tropical second-growth rain forests (Ecology (2005) 86: 7 (1808–1815)). Ecology 2005, 86, 2252. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Maza-Villalobos, S.; Poorter, L.; Martínez-Ramos, M. Effects of ENSO and Temporal Rainfall Variation on the Dynamics of Successional Communities in Old-Field Succession of a Tropical Dry Forest. PLoS ONE 2013, 8, e82040. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ramos, M.; Balvanera, P.; Villa, F.A.; Mora, F.; Maass, J.M.; Méndez, S.M.-V. Effects of long-term inter-annual rainfall variation on the dynamics of regenerative communities during the old-field succession of a neotropical dry forest. For. Ecol. Manag. 2018, 426, 91–100. [Google Scholar] [CrossRef]
- De Arruda, P.H.Z.; Vourlitis, G.L.; Santanna, F.B.; Pinto, O.B., Jr.; De Almeida Lobo, F.; De Souza Nogueira, J. Large net CO2 loss from a grass? Dominated tropical savanna in South? Central Brazil in response to seasonal and interannual drought. J. Geophys. Res. Biogeosci. 2016, 121, 2110–2124. [Google Scholar] [CrossRef]
- Cavaleri, M.A.; Coble, A.P.; Ryan, M.G.; Bauerle, W.L.; Loescher, H.W.; Oberbauer, S.F. Tropical rainforest carbon sink declines during El Niño as a result of reduced photosynthesis and increased respiration rates. New Phytol. 2017, 216, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Rodriguez, S.; Kiese, R.; Sanchez-Azofeifa, A. Seasonality and Budgets of Soil Greenhouse Gas Emissions from a Tropical Dry Forest Successional Gradient in Costa Rica. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005647. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kug, J.; Yoon, J.-H.; Jeong, S. Increased Atmospheric CO2 Growth Rate during El Niño Driven by Reduced Terrestrial Productivity in the CMIP5 ESMs. J. Clim. 2016, 29, 8783–8805. [Google Scholar] [CrossRef]
- Verduzco, V.S.; Garatuza-Payán, J.; Yépez-González, E.; Watts, C.J.; Rodríguez, J.C.; Robles-Morua, A.; Vivoni, E.R. Variations of net ecosystem production due to seasonal precipitation differences in a tropical dry forest of northwest Mexico. J. Geophys. Res. Biogeosci. 2015, 120, 2081–2094. [Google Scholar] [CrossRef]






| Site | AGC Gain (Mg C ha−1 y−1) | BGC Gain (Mg C ha−1 y−1) | Litterfall (Mg C ha−1 y−1) | Carbon Accumulation (Mg C ha−1 y−1) |
|---|---|---|---|---|
| MSSP | ||||
| 20 | 1.7 ± 0.9 a | 0.4 ± 0.2 a | 1.9 ± 0.1 a | 4.0 ± 1.2 a |
| 50 | 2.1 ± 1.2 a | 0.6 ± 0.3 ab | 1.8 ± 0.1 a | 4.5 ± 1.6 a |
| OG | 2.2 ± 0.6 a | 0.6 ± 0.2 b | 2.2 ± 0.2 a | 4.9 ± 0.7 a |
| SRNP | ||||
| 30 | 1.6 ± 0.5 ª | 0.4 ± 0.1 a | 0.6 ± 0.5 a | 2.6 ± 1.1 a |
| 50 | 3.0 ± 1.1 b | 1.3 ± 1.0 b | 2.0 ± 0.5 b | 6.3 ± 2.4 b |
| OG | 2.6 ± 0.8 b | 0.7 ± 0.2 b | 1.9 ± 0.3 b | 5.2 ± 0.8 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvo-Rodriguez, S.; Sánchez-Azofeifa, G.A.; Durán, S.M.; Do Espírito-Santo, M.M.; Ferreira Nunes, Y.R. Dynamics of Carbon Accumulation in Tropical Dry Forests under Climate Change Extremes. Forests 2021, 12, 106. https://doi.org/10.3390/f12010106
Calvo-Rodriguez S, Sánchez-Azofeifa GA, Durán SM, Do Espírito-Santo MM, Ferreira Nunes YR. Dynamics of Carbon Accumulation in Tropical Dry Forests under Climate Change Extremes. Forests. 2021; 12(1):106. https://doi.org/10.3390/f12010106
Chicago/Turabian StyleCalvo-Rodriguez, Sofia, G. Arturo Sánchez-Azofeifa, Sandra M. Durán, Mario Marcos Do Espírito-Santo, and Yule Roberta Ferreira Nunes. 2021. "Dynamics of Carbon Accumulation in Tropical Dry Forests under Climate Change Extremes" Forests 12, no. 1: 106. https://doi.org/10.3390/f12010106
APA StyleCalvo-Rodriguez, S., Sánchez-Azofeifa, G. A., Durán, S. M., Do Espírito-Santo, M. M., & Ferreira Nunes, Y. R. (2021). Dynamics of Carbon Accumulation in Tropical Dry Forests under Climate Change Extremes. Forests, 12(1), 106. https://doi.org/10.3390/f12010106