
Impact of Shrimp Ponds on Mangrove Blue Carbon Stocks in Ecuador

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
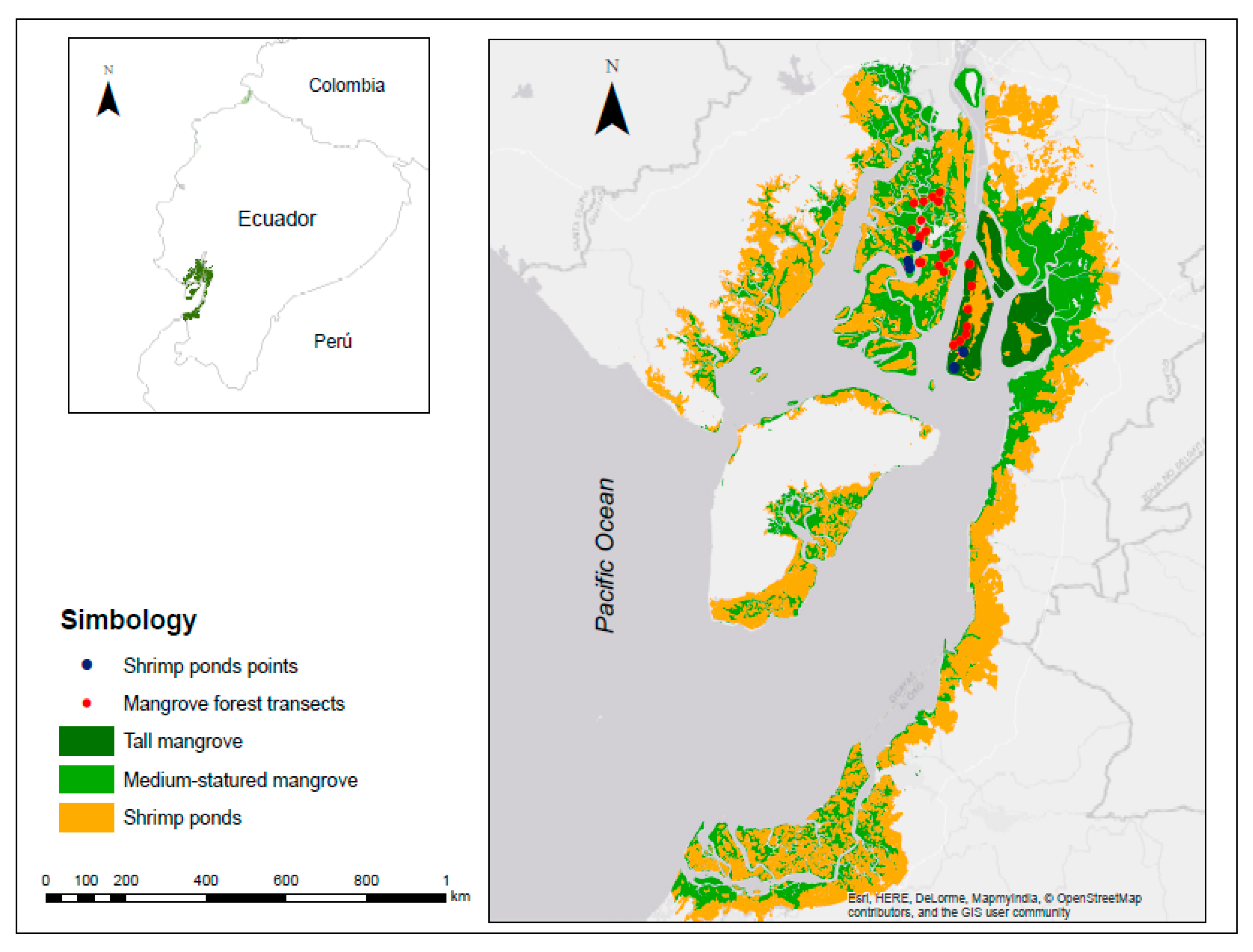
2.1. Study Region
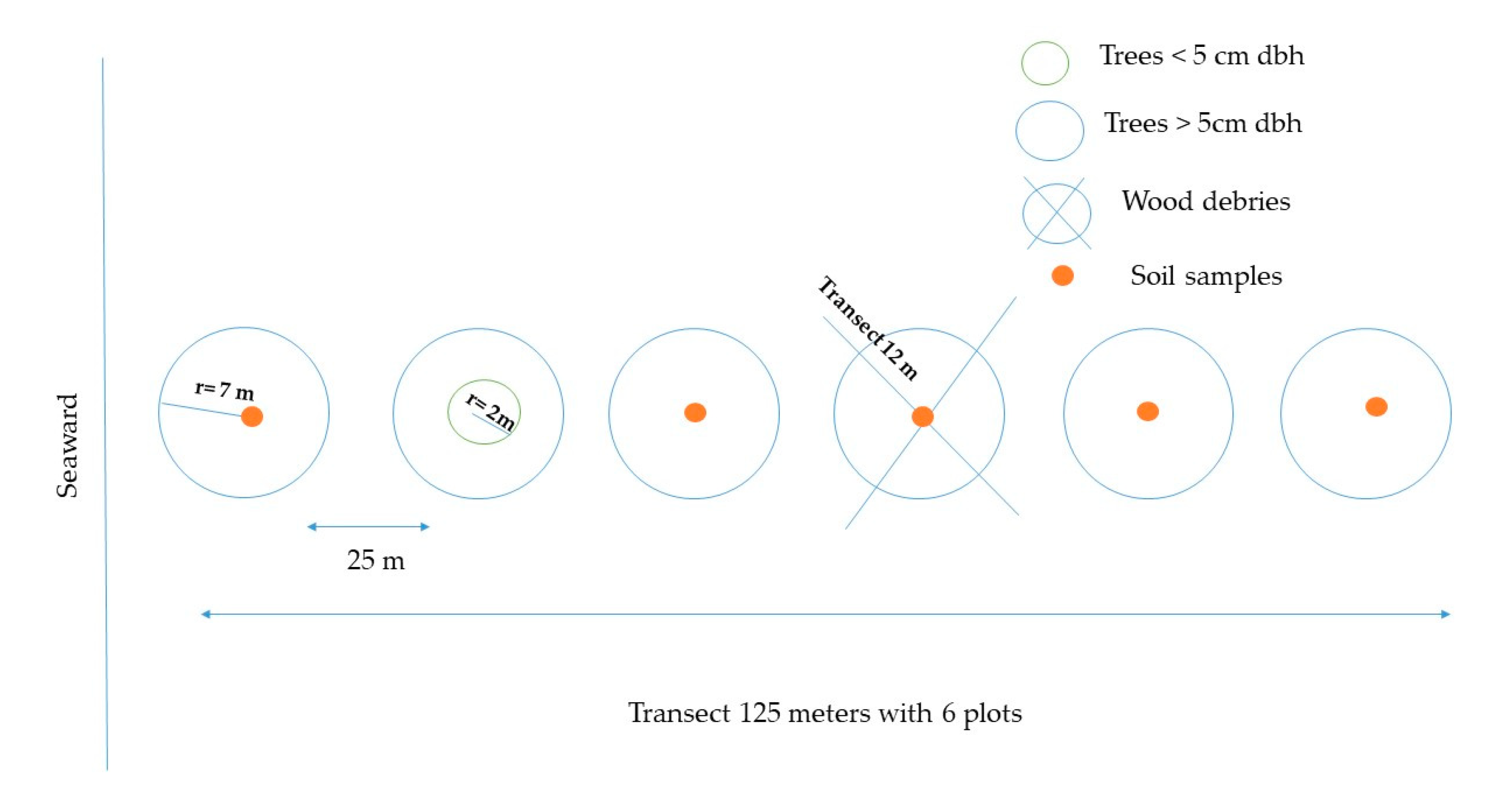
2.2. Forest Stratification and Transects Selection
2.3. Aboveground Carbon
2.4. Soil Carbon
2.5. Statistical Analysis
3. Results
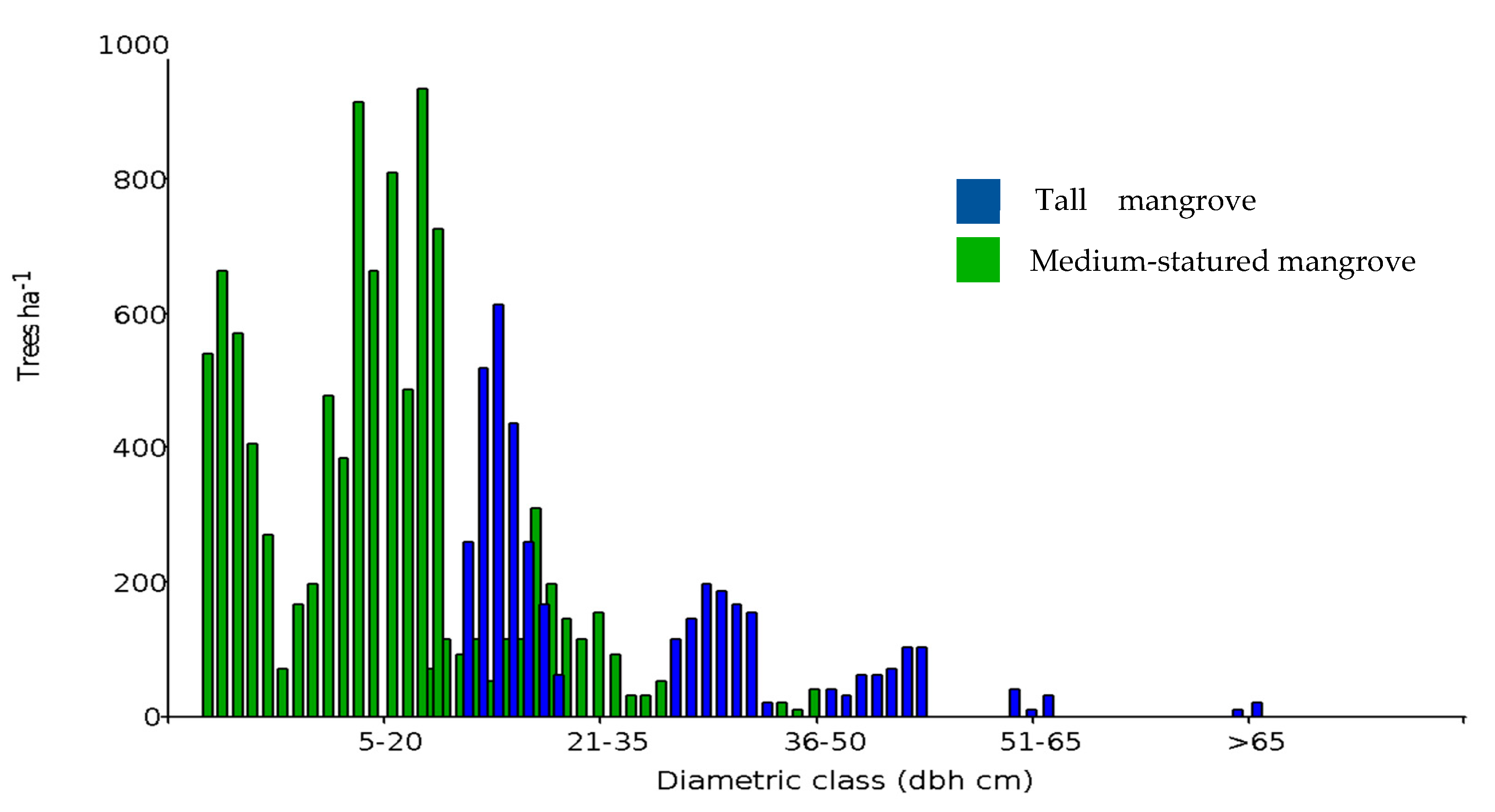
3.1. Mangrove Forest Strata and Shrimp Ponds
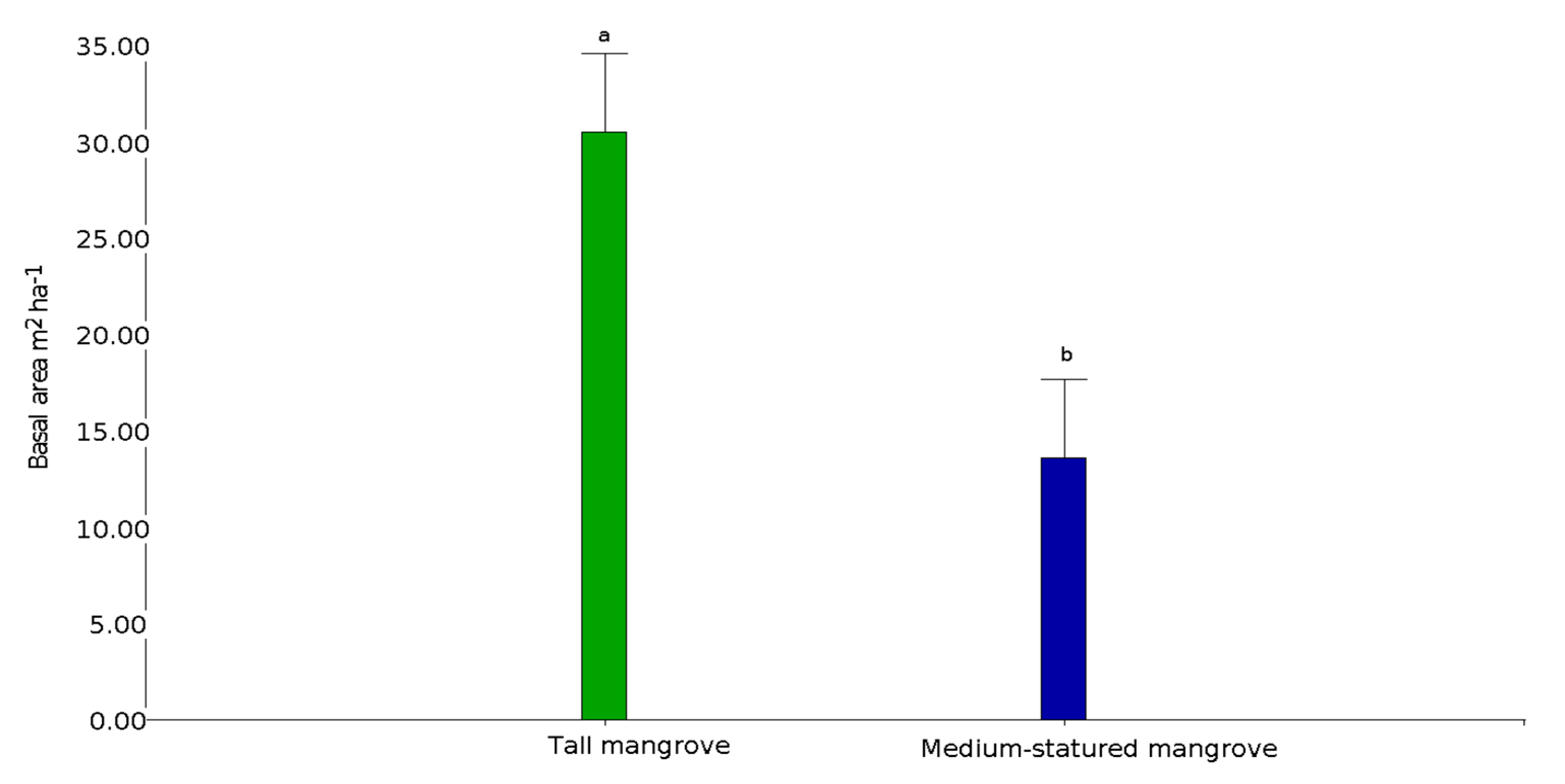
3.2. Aboveground Carbon
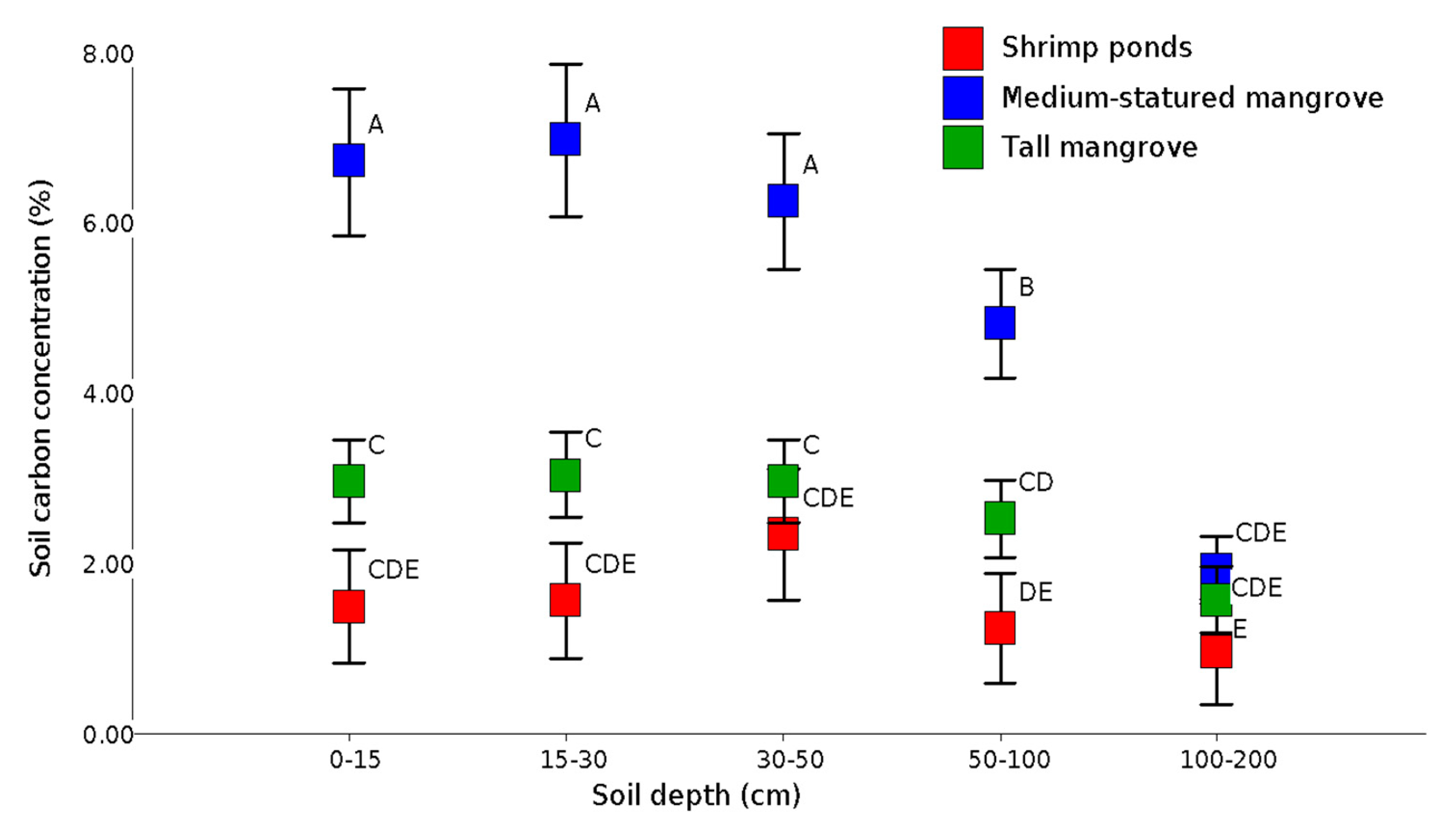
3.3. Soil Carbon
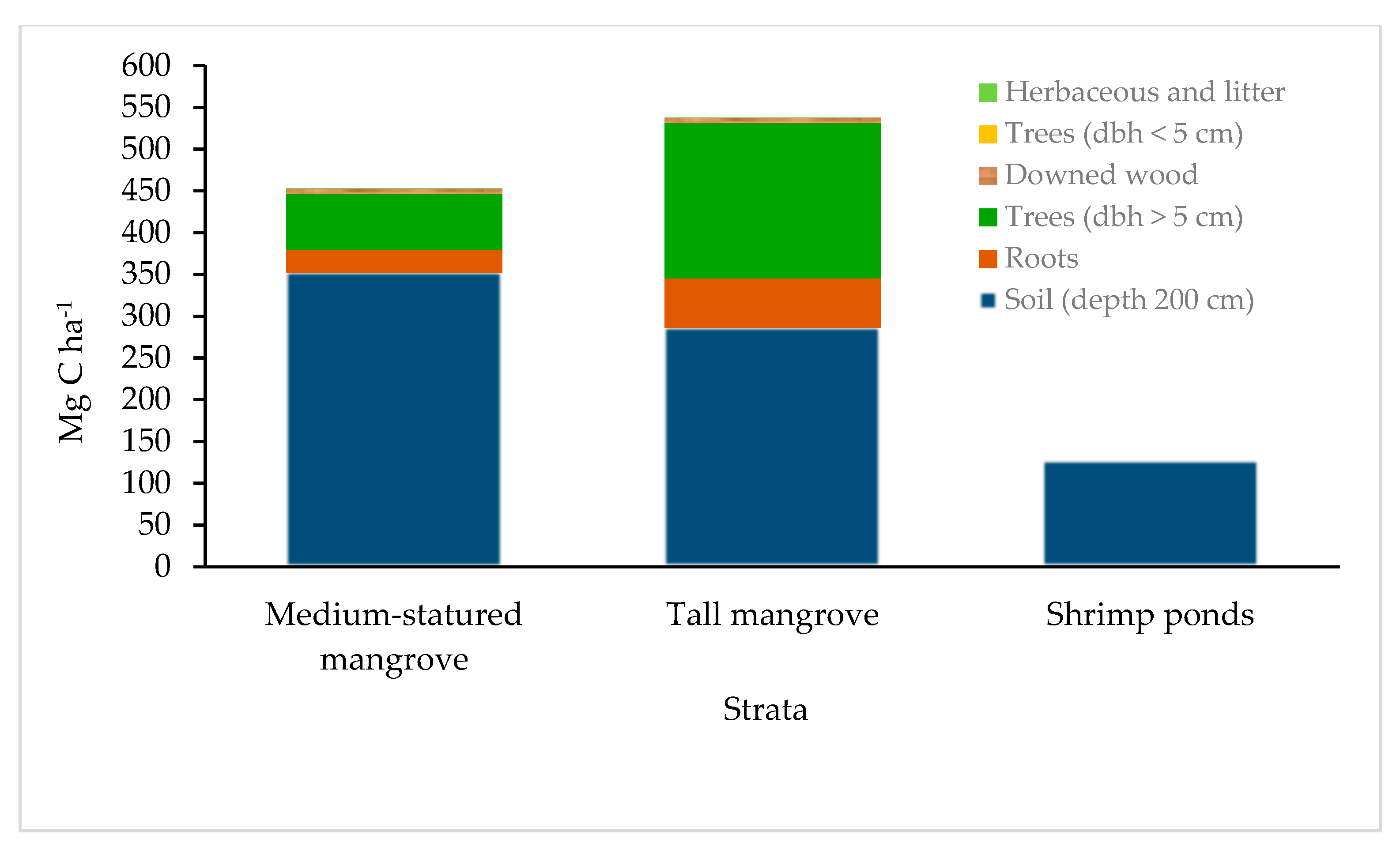
3.4. Total Carbon Stocks
4. Discussion
4.1. Mangrove Forest Strata
4.2. Aboveground Carbon
4.3. Soil Carbon
4.4. Total Carbon Stocks
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Inoue, T. Carbon Sequestration in Mangroves. In Blue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and Implementation; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2019; pp. 73–99. ISBN 9789811312953. [Google Scholar]
- Spalding, M.D.; Kainuma, M.; Collins, L. World Atlas of Mangroves; Earthscan: London, UK, 2010. [Google Scholar]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Cucalón, E. Sistemas Biofísicos y Pesca En El Golfo de Guayaquil: Componente de Oceanografía y Sstemas Físicos; Informe de Consultoría: Guayaquil, Ecuador, 1996; 103p. [Google Scholar]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Herrera-Silveira, J.A. Carbon Stocks of Tropical Coastal Wetlands within the Karstic Landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.J.; Smoak, J.M.; Naidu, A.S.; Sanders, L.M.; Patchineelam, S.R. Organic carbon burial in a mangrove forest, margin and intertidal mud flat. Estuar. Coast. Shelf Sci. 2010, 90, 168–172. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon Cycling and Storage in Mangrove Forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon Cycling in the World’s Mangrove Ecosystems Revisited: Significance of Non-Steady State Diagenesis and Subsurface Linkages between the Forest Floor and the Coastal Ocean. Forests 2020, 11, 977. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove Forests. In Blue Carbon: Coastal Sequestration for Climate Change Mitigation; Alongi, D.M., Ed.; Springer Briefs in Climate Studies; Springer International Publishing: Cham, Switzerland, 2018; pp. 23–36. ISBN 978-3-319-91698-9. [Google Scholar]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Nellemann, C.; Corcoran, E. Blue Carbon: The Role of Healthy Oceans in Binding Carbon: A Rapid Response Assessment; UNEP/Earthprint: Arendal, Norway, 2009; ISBN 978-82-7701-060-1. [Google Scholar]
- Laumonier, Y.; Edin, A.; Kanninen, M.; Munandar, A.W. Landscape-scale variation in the structure and biomass of the hill dipterocarp forest of Sumatra: Implications for carbon stock assessments. For. Ecol. Manag. 2010, 259, 505–513. [Google Scholar] [CrossRef]
- Atwood, T.B.; Connolly, R.M.; Almahasheer, H.; Carnell, P.E.; Duarte, C.M.; Ewers Lewis, C.J.; Irigoien, X.; Kelleway, J.J.; Lavery, P.S.; Macreadie, P.I.; et al. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Chang. 2017, 7, 523–528. [Google Scholar] [CrossRef]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove Forests: One of the World’s Threatened Major Tropical Environments: At least 35% of the area of mangrove forests has been lost in the past two decades, losses that exceed those for tropical rain forests and coral reefs, two other well-known threatened environments. BioScience 2001, 51, 807–815. [Google Scholar]
- Cebrian, J. Variability and control of carbon consumption, export, and accumulation in marine communities. Limnol. Oceanogr. 2002, 47, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Ruess, R.W.; Feller, I.C. CO2 Efflux from Cleared Mangrove Peat. PLoS ONE 2011, 6, e21279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- van der Werf, G.R.; Morton, D.C.; DeFries, R.S.; Olivier, J.G.J.; Kasibhatla, P.S.; Jackson, R.B.; Collatz, G.J.; Randerson, J.T. CO2 emissions from forest loss. Nat. Geosci. 2009, 2, 737–738. [Google Scholar] [CrossRef]
- Centro de Levantamientos Integrados de Recursos por Sensores Remotos (CLIRSEN). Estudio Multitemporal de los Manglares, Camaroenras y Areas Salinas de la Costa Ecuatoriana; CLIRSEN: Quito, Ecuador, 1990. [Google Scholar]
- Rossel, F.; Cadier, E. El Niño and prediction of anomalous monthly rainfalls in Ecuador. Hydrol. Process. Int. J. 2009, 23, 3253–3260. [Google Scholar] [CrossRef]
- Vera, T. Manejo Ambiental Integrado de la Acuicultura del camarón en el Golfo de Guayaquil Apoyado en Sistemas de Información Geográfica. Bachelor’s Thesis, Escuela Politécnica del Litoral, Guayaquil, Ecuador, 2003. [Google Scholar]
- MAE (Ministerio del Ambiente del Ecuador); FAO (Organización de las Naciones Unidas para la Alimentación y la Agricultura). Árboles y Arbustos de los Manglares del Ecuador; Ministerio del Ambiente del Ecuador: Quito. Ecuador, 2014; 48p. [Google Scholar]
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Donato, D.C.; Adame, M.F. Protocolo Para la Medición, Monitoreo y Reporte de la Estructura, Biomasa y Reservas de Carbono de los Manglares; Documento de trabajo 117; CIFOR: Bogor, Indonesia, 2013. [Google Scholar]
- Murdiyarso, D.; Donato, D.; Kauffman, J.B.; Kurnianto, S.; Stidham, M.; Kanninen, M. Carbon storage in mangrove and peatland ecosystems: A preliminary account from plots in Indonesia. Bogor Indonesia Cent. Int. For. Res. 2010, 35, 1–35. [Google Scholar]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change). Good Practice Guidance for Land Use, Land-Use Change and Forestry; Institute for Global Environmental Strategies: Kanagawa, Japan, 2003. [Google Scholar]
- Brown, J.K.; Roussopoulos, P.J. Eliminating Biases in the Planar Intersect Method for Estimating Volumes of Small Fuels. Forest Sci. 1974, 20, 350–356. [Google Scholar]
- Van Wagner, C.E. The Line Intersect Method in Forest Fuel Sampling. For. Sci. 1968, 14, 20–26. [Google Scholar]
- Zanne, A.E.; López-González, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a Worldwide Wood Economics Spectrum. 2009. Available online: https://datadryad.org/stash/dataset/doi:10.5061/dryad.234 (accessed on 4 February 2009).
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1996; pp. 961–1010. ISBN 978-0-89118-866-7. [Google Scholar]
- Sitoe, A.A.; Mandlate, L.J.C.; Guedes, B.S. Biomass and Carbon Stocks of Sofala Bay Mangrove Forests. Forests 2014, 5, 1967–1981. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat versión 2019. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 1 August 2019).
- Hamilton, S.E.; Lovette, J. Ecuador’s Mangrove Forest Carbon Stocks: A Spatiotemporal Analysis of Living Carbon Holdings and Their Depletion since the Advent of Commercial Aquaculture. PLoS ONE 2015, 10, e0118880. [Google Scholar] [CrossRef] [PubMed]
- Terchunian, A.; Klemas, V.; Segovia, A.; Alvarez, A.; Vasconez, B.; Guerrero, L. Mangrove mapping in Ecuador: The impact of shrimp pond construction. Environ. Manag. 1986, 10, 345–350. [Google Scholar] [CrossRef]
- Solis, M.A. Los Manglares Del Ecuador. Rev. Geográfica 1961, 28, 69–88. [Google Scholar] [CrossRef]
- Duke, N.; Ball, M.; Ellison, J. Factors influencing biodiversity and distributional gradients in mangroves. Glob. Ecol. Biogeogr. Lett. 1998, 7, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Estrada, G.C.D.; Soares, M.L.G.; Fernadez, V.; Almeida, P.M.M. de The economic evaluation of carbon storage and sequestration as ecosystem services of mangroves: A case study from southeastern Brazil. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 29–35. [Google Scholar] [CrossRef]
- DelVecchia, A.G.; Bruno, J.F.; Benninger, L.; Alperin, M.; Banerjee, O.; Morales, J.d.D. Organic carbon inventories in natural and restored Ecuadorian mangrove forests. PeerJ 2014, 2, e388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boone Kauffman, J.; Arifanti, V.B.; Hernández Trejo, H.; del Carmen Jesús García, M.; Norfolk, J.; Cifuentes, M.; Hadriyanto, D.; Murdiyarso, D. The jumbo carbon footprint of a shrimp: Carbon losses from mangrove deforestation. Front. Ecol. Environ. 2017, 15, 183–188. [Google Scholar] [CrossRef]
- Chandra, I.A.; Seca, G.; Hena, M.K.A. Aboveground Biomass Production of Rhizophora apiculata Blume in Sarawak Mangrove Forest. Am. J. Agric. Biol. Sci. 2011, 6, 469–474. [Google Scholar]
- Sasmito, S.D.; Sillanpää, M.; Hayes, M.A.; Bachri, S.; Saragi-Sasmito, M.F.; Sidik, F.; Hanggara, B.B.; Mofu, W.Y.; Rumbiak, V.I.; Taberima, S.; et al. Mangrove blue carbon stocks and dynamics are controlled by hydrogeomorphic settings and land-use change. Glob. Chang. Biol. 2020, 26, 3028–3039. [Google Scholar] [CrossRef] [Green Version]
- Urquizo, R.; Carvajal, R.; Salas, J.; Bustamante, M. Biodiversidad Del Guayas: Conociendo Nuestra Verdadera Riqueza; Gobierno Provincial Del Guayas-Dirección de Medio Ambiente: Guayaquil, Ecuador, 2011. [Google Scholar]
- Chatting, M.; LeVay, L.; Walton, M.; Skov, M.W.; Kennedy, H.; Wilson, S.; Al-Maslamani, I. Mangrove carbon stocks and biomass partitioning in an extreme environment. Estuar. Coast. Shelf Sci. 2020, 244, 106940. [Google Scholar] [CrossRef]
- Cifuentes, M.; Brenes, C.; Manrow, M.; Torres, D. Dinámica de uso de la Tierra y Potencial de Mitigación de los Manglares del Golfo de Nicoya; Conservación Internacional: San José, Costa Rica, 2014. [Google Scholar]
- Manrow-Villalobos, M. Estructura, composición florística, biomasa y carbono arriba del suelo en los manglares laguna de Gandoca y estero Limón, Limón, Costa Rica. Rev. For. Mesoam. 2011, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Nóbrega, G.N.; Ferreira, T.O.; Siqueira Neto, M.; Mendonça, E.D.S.; Romero, R.E.; Otero, X.L. The importance of blue carbon soil stocks in tropical semiarid mangroves: A case study in Northeastern Brazil. Environ. Earth Sci. 2019, 78, 369. [Google Scholar] [CrossRef]
- Stringer, C.E.; Trettin, C.C.; Zarnoch, S.J.; Tang, W. Carbon stocks of mangroves within the Zambezi River Delta, Mozambique. For. Ecol. Manag. 2015, 354, 139–148. [Google Scholar] [CrossRef]
- Kristensen, E.; Alongi, D.M. Control by fiddler crabs (Uca vocans) and plant roots (Avicennia marina) on carbon, iron, and sulfur biogeochemistry in mangrove sediment. Limnol. Oceanogr. 2006, 51, 1557–1571. [Google Scholar] [CrossRef] [Green Version]
- Burdige, D.J. Preservation of Organic Matter in Marine Sediments: Controls, Mechanisms, and an Imbalance in Sediment Organic Carbon Budgets? Chem. Rev. 2007, 107, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Marchand, C.; Lallier-Vergès, E.; Baltzer, F. The composition of sedimentary organic matter in relation to the dynamic features of a mangrove-fringed coast in French Guiana. Estuar. Coast. Shelf Sci. 2003, 56, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Kleber, M.; Eusterhues, K.; Keiluweit, M.; Mikutta, C.; Mikutta, R.; Nico, P.S. Chapter One—Mineral–Organic Associations: Formation, Properties, and Relevance in Soil Environments. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 130, pp. 1–140. [Google Scholar]
- Otero, X.L.; Méndez, A.; Nóbrega, G.N.; Ferreira, T.O.; Santiso-Taboada, M.J.; Meléndez, W.; Macías, F. High fragility of the soil organic C pools in mangrove forests. Mar. Pollut. Bull. 2017, 119, 460–464. [Google Scholar] [CrossRef]
- Banerjee, K.; Bal, G.; Mitra, A. How Soil Texture Affects the Organic Carbon Load in the Mangrove Ecosystem? A Case Study from Bhitarkanika, Odisha. In Environmental Pollution. Water Science and Technology Library; Singh, V., Yadav, S., Yadava, R., Eds.; Springer: Singapore, 2018; Volume 77, pp. 329–341. [Google Scholar]
- Sippo, J.Z.; Sanders, C.J.; Santos, I.R.; Jeffrey, L.C.; Call, M.; Harada, Y.; Maguire, K.; Brown, D.; Conrad, S.R.; Maher, D.T. Coastal carbon cycle changes following mangrove loss. Limnol. Oceanogr. 2020, 65, 2642–2656. [Google Scholar] [CrossRef]
- Johnson, J.L.; Raw, J.L.; Adams, J.B. First report on carbon storage in a warm-temperate mangrove forest in South Africa. Estuar. Coast. Shelf Sci. 2020, 235, 106566. [Google Scholar] [CrossRef]
- Eid, E.M.; Arshad, M.; Shaltout, K.H.; El-Sheikh, M.A.; Alfarhan, A.H.; Picó, Y.; Barcelo, D. Effect of the conversion of mangroves into shrimp farms on carbon stock in the sediment along the southern Red Sea coast, Saudi Arabia. Environ. Res. 2019, 176, 108536. [Google Scholar] [CrossRef] [PubMed]
- Sidik, F.; Lovelock, C.E. CO2 Efflux from Shrimp Ponds in Indonesia. PLoS ONE 2013, 8, e66329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidik, F.; Adame, M.F.; Lovelock, C.E. Carbon sequestration and fluxes of restored mangroves in abandoned aquaculture ponds. J. Indian Ocean Reg. 2019, 15, 177–192. [Google Scholar] [CrossRef]
- Baccini, A.; Goetz, S.J.; Walker, W.S.; Laporte, N.T.; Sun, M.; Sulla-Menashe, D.; Hackler, J.; Beck, P.S.A.; Dubayah, R.; Friedl, M.A.; et al. Estimated carbon dioxide emissions from tropical deforestation improved by carbon-density maps. Nat. Clim. Chang. 2012, 2, 182–185. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Adame, M.F.; Arifanti, V.B.; Schile-Beers, L.M.; Bernardino, A.F.; Bhomia, R.K.; Donato, D.C.; Feller, I.C.; Ferreira, T.O.; Jesus Garcia, M.D.C.; et al. Total ecosystem carbon stocks of mangroves across broad global environmental and physical gradients. Ecol. Monogr. 2020, 90, e01405. [Google Scholar] [CrossRef] [Green Version]
- Palacios Peñaranda, M.L.; Cantera Kintz, J.R.; Peña Salamanca, E.J. Carbon stocks in mangrove forests of the Colombian Pacific. Estuar. Coast. Shelf Sci. 2019, 227, 106299. [Google Scholar] [CrossRef]
- Marchand, C. Soil carbon stocks and burial rates along a mangrove forest chronosequence (French Guiana). For. Ecol. Manag. 2017, 384, 92–99. [Google Scholar] [CrossRef]
- Queiroz, H.M.; Artur, A.G.; Taniguchi, C.A.K.; da Silveira, M.R.S.; do Nascimento, J.C.; Nóbrega, G.N.; Otero, X.L.; Ferreira, T.O. Hidden contribution of shrimp farming effluents to greenhouse gas emissions from mangrove soils. Estuar. Coast. Shelf Sci. 2019, 221, 8–14. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marbà, N.; et al. Estimating Global “Blue Carbon” Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Aboveground Biomass | Root Biomass | n | r2 |
|---|---|---|---|---|
| Rhizophora mangle | 0.128 × (dbh) 2.6 | = 0.199 × (ρ) 0.899 × (dbh) 2.22 | 9 | 0.92 |
| Laguncularia racemosa | 0.1023 × (dbh) 2.5 | 70 | 0.97 |
| Mangrove Stratum | Transect | Island | Species | Height | Basal Area | Tree Density |
|---|---|---|---|---|---|---|
| Medium-statured | T11 | San Ignacio | Rhiz | 18.0 ± 2.4 | 10.5 ± 4.3 | 671 ± 281 |
| Medium-statured | T12 | Canoa | Rhiz | 18.2 ± 1.5 | 16.2 ± 6.8 | 942 ± 477 |
| Medium-statured | T13 | San Ignacio | Rhiz | 19.6 ± 3.6 | 16.8 ± 9.0 | 769 ± 271 |
| Medium-statured | T14 | San Ignacio | Rhiz | 16.1 ± 4.1 | 18.5 ± 9.8 | 693 ± 268 |
| Medium-statured | T15 | Chupadores Chico | Rhiz | 8.4 ± 0.8 | 3.2 ± 1.7 | 487 ± 99 |
| Medium-statured | T16 | Chupadores Chico | Rhiz | 13.0 ± 5.1 | 8.8 ± 5.7 | 379 ± 139 |
| Medium-statured | T17 | Chupadores Chico | Rhiz/Lag | 21.1 ± 2.9 | 13.9 ± 4.1 | 531 ± 156 |
| Medium-statured | T18 | Chupadores | Rhiz | 20.8 ± 3.6 | 14.0 ± 2.8 | 498 ± 140 |
| Medium-statured | T19 | San Ignacio | Rhiz | 22.7 ± 1.7 | 27.7 ± 9.9 | 1007 ± 158 |
| Medium-statured | T20 | Canoa | Rhiz | 23.0 ± 4.2 | 23.1 ± 13.0 | 736 ± 168 |
| Medium-statured | T21 | Bellavista | Rhiz | 17.7 ± 1.7 | 20.2 ± 7.9 | 1321 ± 853 |
| Medium-statured | T22 | Chupadores Chico | Rhiz | 12.9 ± 2.2 | 19.2 ± 11.2 | 1039 ± 344 |
| Medium-statured | T4 | Tortuga | Rhiz | 22.9 ± 1.5 | 26.6 ± 8.7 | 1180 ± 255 |
| Medium-statured | T5 | Tortuga | Rhiz/Lag | 19.8 ± 1.4 | 10.1 ± 3.4 | 693 ± 333 |
| Medium-statured | T6 | Tortuga | Rhiz | 18.4 ± 1.6 | 10.0 ± 4.5 | 1061 ± 333 |
| Medium-statured | T7 | Bocana | Rhiz | 14.4 ± 2.7 | 11.1 ± 4.2 | 888 ± 262 |
| Medium-statured | T8 | Bocana | Rhiz/Lag | 16.7 ± 2.9 | 12.0 ± 2.3 | 1288 ± 238 |
| Tall | TM1 | Mondragón | Rhiz | 22.4 ± 6.0 | 32.6 ± 17.9 | 585 ± 179 |
| Tall | TM10 | Mondragón | Rhiz/Lag | 11.8 ± 2.8 | 18.7 ± 9.5 | 985 ± 186 |
| Tall | TM11 | Mondragón | Rhiz/Lag | 14.5 ± 2.1 | 29.2 ± 11.5 | 1245 ± 289 |
| Tall | TM12 | Mondragón | Rhiz | 17.8 ± 2.4 | 34.3 ± 16.8 | 1104 ± 448 |
| Tall | TM3 | Mondragón | Rhiz | 17.3 ± 1.4 | 28.0 ± 14.4 | 660 ± 271 |
| Tall | TM8 | Mondragón | Rhiz | 17.7 ± 3.1 | 31.9 ± 10.0 | 812 ± 365 |
| Tall | TM9 | Mondragón | Rhiz | 23.5 ± 4.7 | 40.9 ± 20.0 | 401 ± 156 |
| Seaward Edge (m) | Aboveground Carbon (Mg·ha−1) | |
|---|---|---|
| 25 | 85.8 | a |
| 50 | 86.7 | a |
| 75 | 83.0 | a |
| 100 | 78.3 | a |
| 125 | 59.1 | b |
| 150 | 48.6 | b |
| Medium-Statured Mangroves | Tall Mangroves | Shrimp Ponds | |
|---|---|---|---|
| Transect | 17 | 7 | 5 |
| Trees > 5 cm dbh | 67.88 ± 8.39 | 186 ± 26.76 | -- |
| Trees < 5 cm dbh | 0.38 ± 0.20 | 0.06 ± 0.03 | -- |
| Woody debris | 5.36 ± 1.00 | 6.05 ± 3.61 | -- |
| Total aboveground carbon | 73.62 ± 8.39 a | 192.20 ± 29.90 b | -- |
| Roots | 26.76 ± 2.88 | 59.06 ± 6.16 | -- |
| Soil 1 m | 220.57 ± 22.89 | 168.19 ± 21.30 | 81.91 ± 13.69 |
| Soil 2 m | 352.51 ± 31.49 | 286.39 ± 38.27 | 126.98 ± 16.33 |
| Total soil and roots carbon (1 m) | 247.33 ± 21.81 a | 227.25 ± 26.57 a | 81.91 ± 13.69 b |
| Total soil and roots carbon (2 m) | 379.26 ± 30.35 a | 345.46 ± 43.57 a | 126.98 ± 16.33 b |
| Ecosystem carbon (1 m) | 320.95 ± 20.88 a | 419.4 5± 55.66 a | 81.91 ± 13.69 b |
| Ecosystem carbon (2 m) | 452.13 ± 28.32 a | 537.65 ± 66.70 a | 126.98 ± 16.33 b |
| Land Use | Area (ha) | C (Mg ha−1) | Total Carbon(Mg C) | Mg CO2e |
|---|---|---|---|---|
| Medium-statured mangrove | 117,746.0 | 320.9 | 37,790,578.7 | 138,691,423.8 |
| Tall mangrove | 16,317.1 | 419.4 | 6,844,224.3 | 25,118,303.4 |
| Shrimp ponds | 153,950.4 | 81.9 | 12,610,084.6 | 46,279,010.6 |
| Total | 288,013.6 | 57,244,887.7 | 210,088,737.8 |
| Country | Height (m) | Species | Aboveground Carbon (Mg ha−1) | Author |
|---|---|---|---|---|
| Dominican Republic | >10 | R. mangle | 133 | [40] |
| Honduras | R. mangle–L. racemosa | 143.9 | ||
| Costa Rica | R. mangle | 151 | ||
| Mexico | R. mangle | 236 | ||
| Ecuador | R. mangle–L. racemosa | 108.2 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merecí-Guamán, J.; Casanoves, F.; Delgado-Rodríguez, D.; Ochoa, P.; Cifuentes-Jara, M. Impact of Shrimp Ponds on Mangrove Blue Carbon Stocks in Ecuador. Forests 2021, 12, 816. https://doi.org/10.3390/f12070816
Merecí-Guamán J, Casanoves F, Delgado-Rodríguez D, Ochoa P, Cifuentes-Jara M. Impact of Shrimp Ponds on Mangrove Blue Carbon Stocks in Ecuador. Forests. 2021; 12(7):816. https://doi.org/10.3390/f12070816
Chicago/Turabian StyleMerecí-Guamán, Jéssica, Fernando Casanoves, Diego Delgado-Rodríguez, Pablo Ochoa, and Miguel Cifuentes-Jara. 2021. "Impact of Shrimp Ponds on Mangrove Blue Carbon Stocks in Ecuador" Forests 12, no. 7: 816. https://doi.org/10.3390/f12070816
APA StyleMerecí-Guamán, J., Casanoves, F., Delgado-Rodríguez, D., Ochoa, P., & Cifuentes-Jara, M. (2021). Impact of Shrimp Ponds on Mangrove Blue Carbon Stocks in Ecuador. Forests, 12(7), 816. https://doi.org/10.3390/f12070816