
Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove


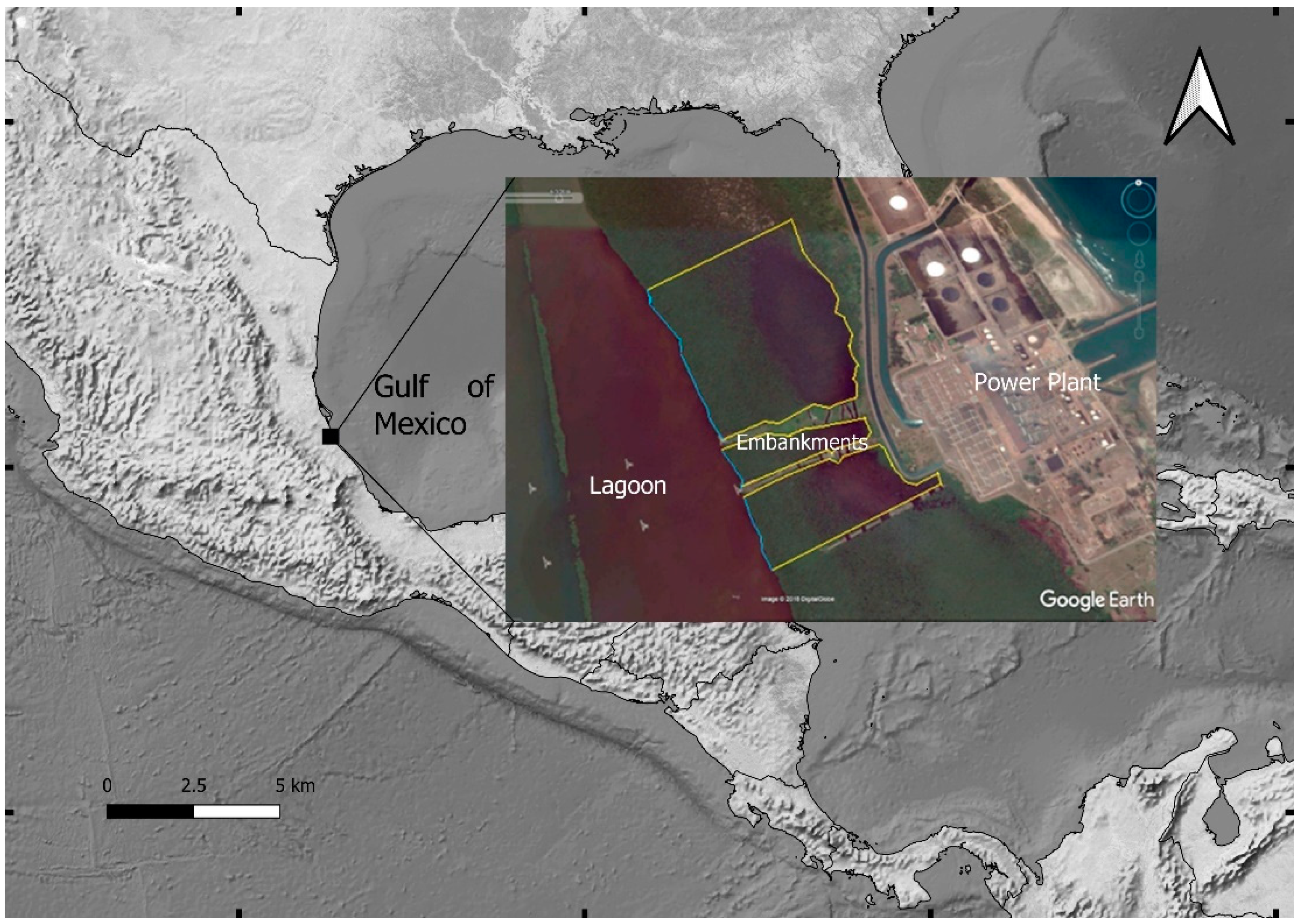{kind=link}
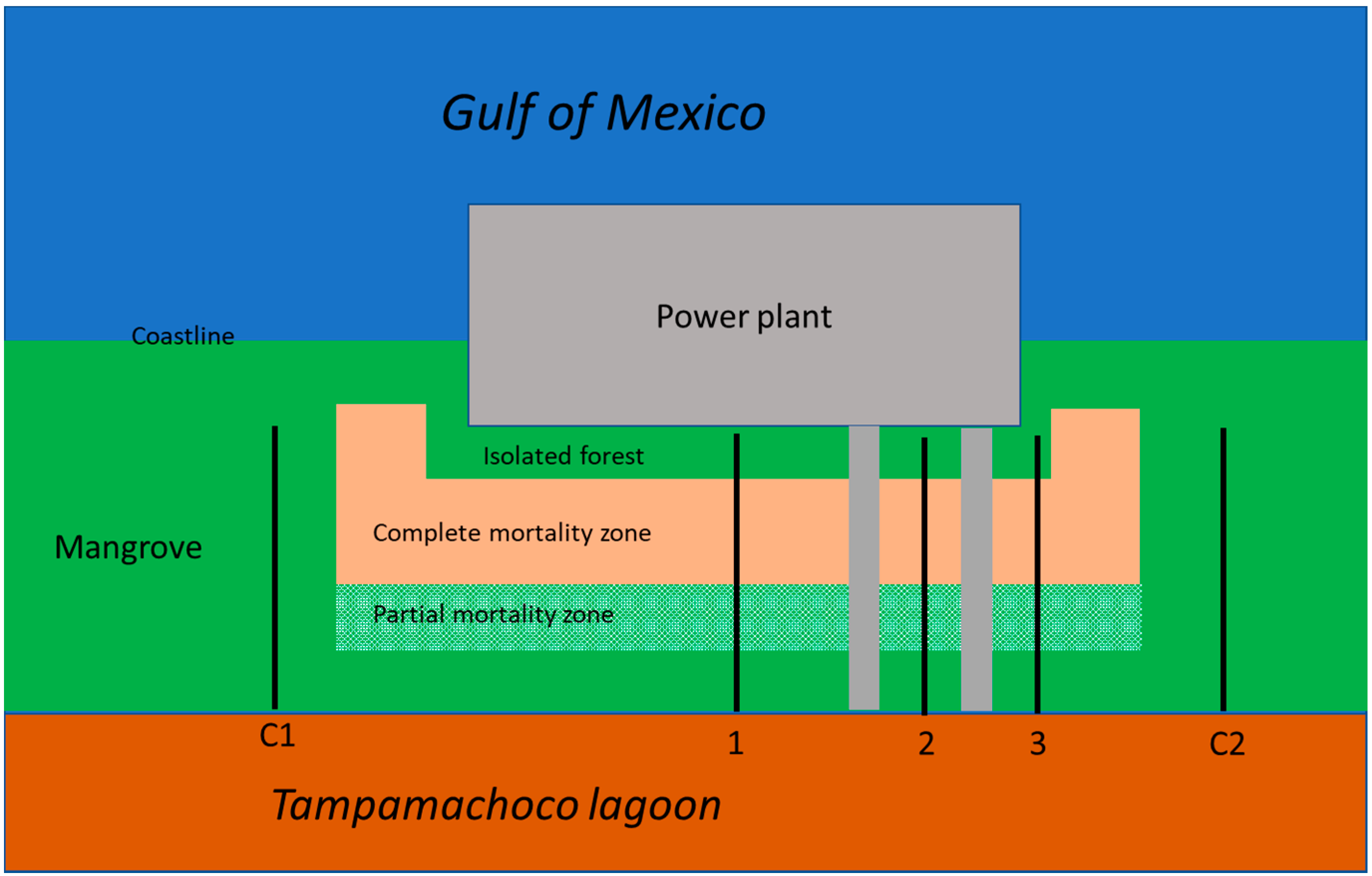{kind=link}
{kind=link}
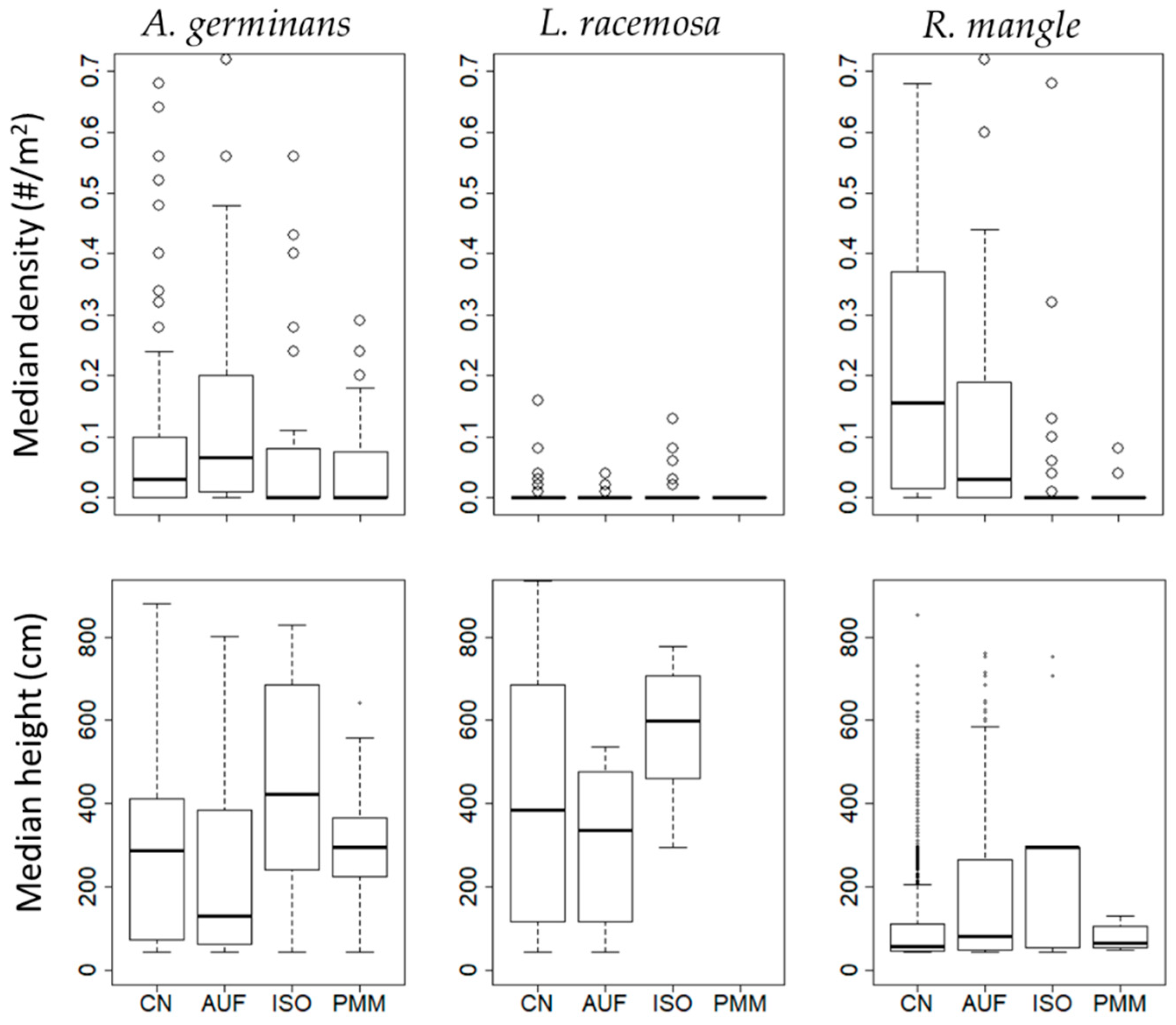{kind=link}
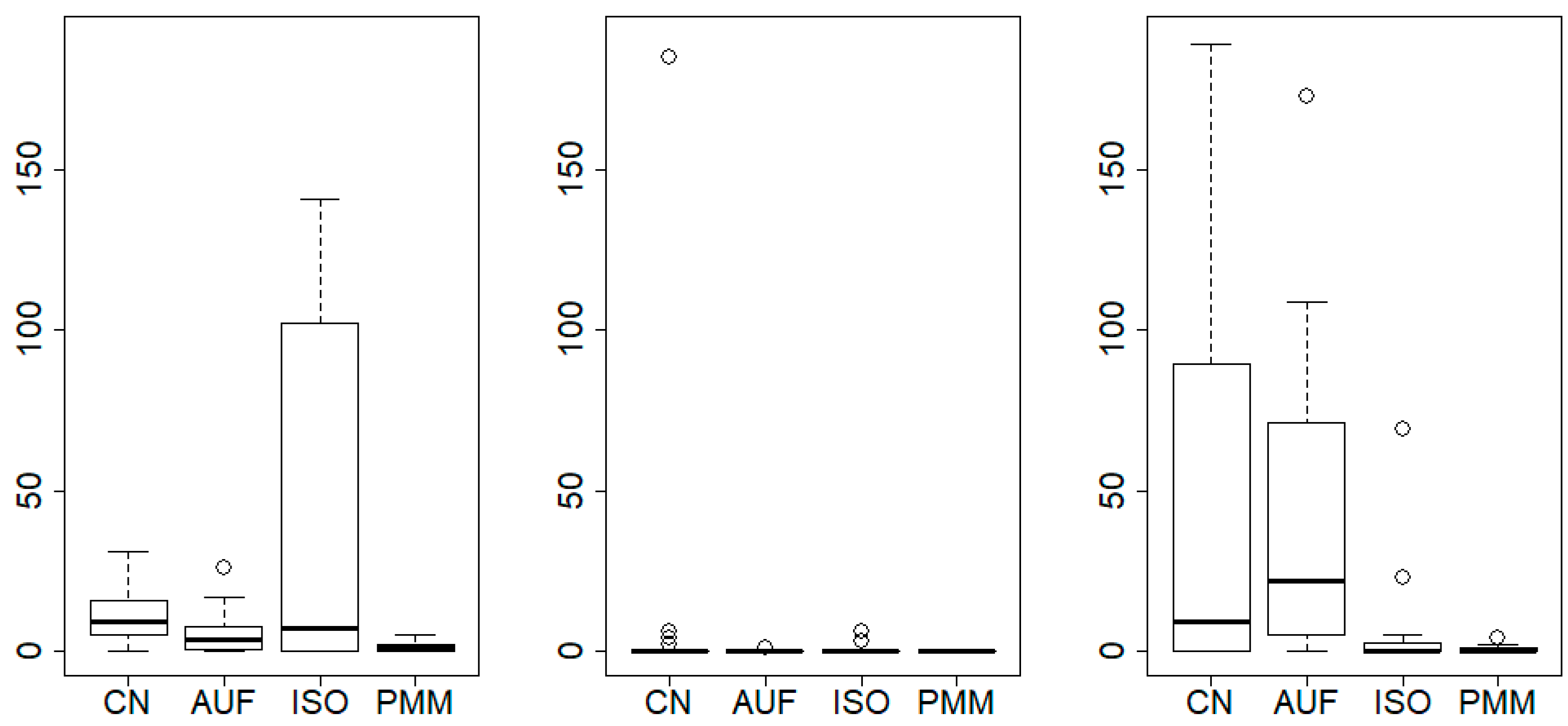{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Forest Structure
3.2. Projections of Avicennia germinans (L.) L.
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ewel, K.C.; Twilley, R.R.; Ong, J.E. Different Kinds of Mangrove Forests Provide Different Goods and Services. Glob. Ecol. Biogeogr. Lett. 1998, 7, 83–94. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemuller, R.; Roy, D.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Feher, L.C.; Griffith, K.T.; Cavanaugh, K.C.; Enwright, N.; Day, R.; Stagg, C.; Krauss, K.W.; Howard, R.J.; Grace, J.B.; et al. Climatic controls on the global distribution, abundance, and species richness of mangrove forests. Ecol. Monogr. 2017, 87, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Sasmito, S.D.; Murdiyarso, D.; Friess, D.A.; Kurnianto, S. Can mangroves keep pace with contemporary sea level rise? A global data review. Wetl. Ecol. Manag. 2016, 24, 263–278. [Google Scholar] [CrossRef]
- Lambin, E.F.; Meyfroidt, P. Land use transitions: Socio-ecological feedback versus socio-economic change. Land Use Policy 2010, 27, 108–118. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Eddy, S.; Milantara, N.; Sasmito, S.; Kajita, T.; Basyuni, M. Anthropogenic Drivers of Mangrove Loss and Associated Carbon Emissions in South Sumatra, Indonesia. Forests 2021, 12, 187. [Google Scholar] [CrossRef]
- Lara-Lara, J.R.; Arreola-Lizárraga, J.A.; Calderón-Aguilera, L.E.; Camacho-Ibar, V.F.; De la Lanza-Espino, G.; Escofet-Giansone, A.; Espejel-Carbajal, M.I.; Guzmán-Arroyo, M.; Ladah, L.B.; López-Hernández, M.; et al. Los ecosistemas costeros, insulares y epicontinentales. Cap. Nat. México 2008, 1, 109–134. [Google Scholar]
- Vazquez-Lule, A.; Colditz, R.; Herrera-Silveira, J.; Guevara, M.; Rodríguez-Zúñiga, M.T.; Cruz, I.; Ressl, R.; Vargas, R. Greenness trends and carbon stocks of mangroves across Mexico. Environ. Res. Lett. 2019, 14, 075010. [Google Scholar] [CrossRef]
- Feller, I.C.; Lovelock, C.; Berger, U.; McKee, K.; Joye, S.; Ball, M. Biocomplexity in Mangrove Ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Valderrama-Landeros, L.H.; Rodríguez-Zúñiga, M.T.; Troche-Souza, C.; Velázquez-Salazar, S.; Villeda-Chávez, E.; Alcántara-Maya, J.A.; Vázquez-Balderas, B.; Cruz-López, M.I.; Ressl, R. Manglares de México: Actualización y Exploración de los Datos del Sistema de Monitoreo 1970/1980–2015; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico city, Mexico, 2017; p. 128. [Google Scholar]
- Moreno-Casasola, P.; Cejudo, E.; Monroy, R.C.; Infante, D.M.; López, H.; Peralta, L.A.; Flores, I.N.; Castillo-Campo, G.; Madero, C.; Capistrán, A.; et al. IV. Manglares, selvas inundables y humedales herbáceos. In Servicios Ecosistémicos de las Selvas y Bosques Costeros de Veracruz; INECOL-ITTO-CONAFOR-INECC: Xalapa, Mexico, 2016; p. 358. [Google Scholar]
- Cano-Ortiz, A.; Musarella, C.; Fuentes, C.; Pinto-Gomes, C.; Río, S.; Cano, E. Diversity and conservation status of mangrove communities in two areas of Mesocaribea biogeographic region. Curr. Sci. 2018, 115, 534–540. [Google Scholar] [CrossRef]
- de Jesús Basáñez Muñoz, A.; Olmedo, G.; Rojas, P. Características estructurales y usos del manglar en el ejido Cerro de Tumilco, Tuxpan, Veracruz, México. Revista UDO Agrícola 2006, 6, 114–120. [Google Scholar]
- de Jesús Basáñez Muñoz, A.; Cruz, M.A.; Domínguez, C.; González, C.; Serrano, A.; Hernández, A. Estructura y producción de Conocarpus erectus L. en el Sitio Ramsar “Manglares y Humedales de Tuxpan”, Veracruz, México. Revista UDO Agrícola 2008, 8, 78–87. [Google Scholar]
- López Portillo, J.A.; Lara, A.L.; Sáinz, E.; Vásquez, V.M.; Rodríguez, M.; Martínez, M.C.; Bartolo, O.; Ortiz, I.I.; Alvarado, G. Restauración Hidráulica en la Laguna de Tampamachoco en el Estado de Veracruz para la Rehabilitación del Manglar y de sus Servicios Ambientales; Informe Final SNIB-CONABIO. Proyecto No.HH025; Instituto de Ecología A.C.: Xalapa, Mexico, 2014; 28p. [Google Scholar]
- Vovides, A.G.; López-Portillo, J.; Bashan, Y. N2-fixation along a gradient of long-term disturbance in tropical mangroves bordering the Gulf of Mexico. Biol. Fertil. Soils 2011, 47, 567–576. [Google Scholar] [CrossRef]
- de Jesús Basáñez Muñoz, A.; Cuervo-López, L.; Serrano-Solis, A.; Cárdenas-del Ángel, S.I. Ensayos de reforestación con plantas de mangle en la Reserva Ecológica del Complejo Termoeléctrico “Presidente Adolfo López Mateos”, Tuxpan, Veracruz. Polibotánica 2016, 42, 91–101. [Google Scholar]
- Osland, M.J.; Feher, L.C.; López-Portillo, J.; Day, R.; Suman, D.O.; Menéndez, J.M.G.; Rivera-Monroy, V.H. Mangrove forests in a rapidly changing world: Global change impacts and conservation opportunities along the Gulf of Mexico coast. Estuar. Coast. Shelf Sci. 2018, 214, 120–140. [Google Scholar] [CrossRef]
- Cintrón-Molero, G.; Schaeffer-Novelli, Y. Ecology and management of New World mangroves. In Coastal Plant Communities of Latin America; Academic Press: Cambridge, MA, USA, 1992; pp. 233–258. [Google Scholar]
- Kairo, J.G.; Dahdouh-Guebas, F.; Gwada, P.O.; Ochieng, C.; Koedam, N. Regeneration status of mangrove forests in Mida Creek Kenya: A compromised or secured future? Ambio 2002, 31, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Clough, B.; Scott, K. Allometric relationships for estimating above-ground biomass in six mangrove species. For. Ecol. Manag. 1989, 27, 117–127. [Google Scholar] [CrossRef]
- Jones, A.R.; Raja Segaran, R.; Clarke, K.D.; Waycott, M.; Goh, W.S.H.; Gillanders, B.M. Estimating Mangrove Tree Biomass and Carbon Content: A Comparison of Forest Inventory Techniques and Drone Imagery. Front. Mar. Sci. 2020, 6, 784. [Google Scholar] [CrossRef] [Green Version]
- Clarke, P.J. The population dynamics of the mangrove Avicennia marina; demographic synthesis and predictive modelling. Hydrobiologia 1995, 295, 83–88. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001; 175p. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Clarke, P.J. Predispersal mortality and fecundity in the grey mangrove (Avicennia marina) in southeastern Australia. Aust. J. Ecol. 1992, 17, 161–168. [Google Scholar] [CrossRef]
- Stubben, C.J.; Milligan, B.G. Estimating and Analyzing Demographic Models Using the popbio Package in R. J. Stat. Softw. 2007, 22. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Keyfitz, N.; Caswell, H. Applied Mathematical Demography, 3rd ed.; Springer: New York, NY, USA, 2005; p. 555. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Day, J.W., Jr.; Conner, W.H.; Ley-Lou, F.; Day, R.H.; Navarro, A.M. The productivity and composition of mangrove forests, Laguna de Terminos, Mexico. Aquat. Bot. 1987, 27, 267–284. [Google Scholar] [CrossRef]
- Satyanarayana, B.; Raman, A.V.; Mohd-Lokman, H.; Dehairs, F.; Sharma, V.S.; Dahdouh-Guebas, F. Multivariate methods distinguishing mangrove community structrue of Coringa in the Godavari Delta, East coast of India. Acuat. Ecosyst. Heath Manag. 2009, 12, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Méndez, C.; Jiménez, J. Análisis de la asociación entre la estructura y composición de la comunidad vegetal de manglar, y los parámetros físico-químicos del agua en la Bahía La Graciosa, Izabal, Guatemala. Revista Científica Instituto Investigaciones Químicas Biológicas 2015, 25, 50–64. [Google Scholar]
- Natividad, E.M.C.; Hingabay, V.S.; Lipae, R.S.; Requieron, E.A.; Abulunan, A.J.; Tagaloguin, P.M.; Flamiano, R.S.; Jumawan, J.H.; Jurnawan, J.C. Vegetation analysis and community structure of mangroves in Alabel and Maasim Sarangani Province, Philippines. ARPN J. Agric. Biol. Sci. 2015, 10, 97–102. [Google Scholar]
- Pool, D.J.; Snedaker, S.C.; Lugo, A.E. Structure of Mangrove Forests in Florida, Puerto Rico, Mexico, and Costa Rica. Biotropica 1977, 9, 195–212. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.R.; Milbrandt, E.; Brown, B.; Krauss, K.W.; Rovai, A.; Beever, J.W.; Flynn, L.L. Stress in mangrove forests: Early detection and preemptive rehabilitation are essential for future successful worldwide mangrove forest management. Mar. Pollut. Bull. 2016, 109, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Macamo, C.D.C.F.; Adams, J.B.; Bandeira, S.O.; Mabilana, H.A.; António, V.M. Spatial Dynamics and Structure of Human Disturbed Mangrove Forests in Contrasting Coastal Communities in Eastern Africa. Wetlands 2018, 38, 509–523. [Google Scholar] [CrossRef]
- Ross, M.S.; Ruiz, P.L.; Sah, J.P.; Hanan, E. Chilling damage in a changing climate in coastal landscapes of the subtropical zone: A case study from south Florida. Glob. Chang. Biol. 2009, 15, 1817–1832. [Google Scholar] [CrossRef]
- Madrid, E.N.; Armitage, A.R.; Portillo, J.L. Avicennia germinans (black mangrove) vessel architecture is linked to chilling and salinity tolerance in the Gulf of Mexico. Front. Plant Sci. 2014, 5, 503. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Krauss, K.W.; Osland, M.J.; Reef, R.; Ball, M.C. The physiology of mangrove trees with changing climate. In Tropical Tree Physiology; Springer: Cham, Switzerland, 2016; pp. 149–179. [Google Scholar]
- Landsberg, J.J. Physiological Ecology of Forest Production; Academic Press: London, UK, 1986; 282p. [Google Scholar]
- Ali, A.; Alfarhan, A.; Robinson, E.; Aldjain, I. Pattern of Survival and Mortality of Mangrove Populations Grown at Al-Jubail Area (Saudi Arabia) of the Arabian Gulf. Am. J. Agric. Biol. Sci. 2008, 3, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Clarke, P.J.; Allaway, W.G. The regeneration niche of the grey mangrove (Avicennia marina): Effects of salinity, light and sediment factors on establishment, growth and survival in the field. Oecologia 1993, 93, 548–556. [Google Scholar] [CrossRef]
- McKee, K. Soil Physicochemical Patterns and Mangrove Species Distribution—Reciprocal Effects? J. Ecol. 1993, 81, 477–487. [Google Scholar] [CrossRef]
- Condit, R.; Sukumar, R.; Hubbell, S.P.; Foster, R.B. Predicting population trends from size distributions: A direct test in a tropical tree community. Am. Nat. 1998, 152, 495–509. [Google Scholar] [CrossRef]
- Burslem, D.F.R.P. Natural disturbance in forest environments. In Encyclopedia of Forest Sciences; Elsevier: Amsterdam, The Netherlands, 2004; pp. 80–85. [Google Scholar]
- Riascos, J.M.; Cantera, J.R.; Blanco-Libreros, J.F. Growth and mortality of mangrove seedlings in the wettest neotropical mangrove forests during ENSO: Implications for vulnerability to climate change. Aquat. Bot. 2018, 147, 34–42. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Yessoufou, K. Strong seedling recruitment does not limit mangrove vulnerability to harvest. Environ. Res. Lett. 2019, 14, 064019. [Google Scholar] [CrossRef]
- Delgado, P.; Jiménez, J.; Justic, D. Population dynamics of mangrove Avicennia bicolor on the Pacific coast of Costa Rica. Wetl. Ecol. Manag. 1999, 7, 113–120. [Google Scholar] [CrossRef]
- Devaney, J.L.; Lehmann, M.; Feller, I.C.; Parker, J.D. Mangrove microclimates alter seedling dynamics at the range edge. Ecology 2017, 98, 2513–2520. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basáñez-Muñoz, A.d.J.; Jordán-Garza, A.G.; Serrano, A. Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove. Forests 2021, 12, 989. https://doi.org/10.3390/f12080989
Basáñez-Muñoz AdJ, Jordán-Garza AG, Serrano A. Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove. Forests. 2021; 12(8):989. https://doi.org/10.3390/f12080989
Chicago/Turabian StyleBasáñez-Muñoz, Agustín de Jesús, Adán Guillermo Jordán-Garza, and Arturo Serrano. 2021. "Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove" Forests 12, no. 8: 989. https://doi.org/10.3390/f12080989
APA StyleBasáñez-Muñoz, A. d. J., Jordán-Garza, A. G., & Serrano, A. (2021). Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove. Forests, 12(8), 989. https://doi.org/10.3390/f12080989