
Species-Specific Allometric Equations for Predicting Belowground Root Biomass in Plantations: Case Study of Spotted Gums (Corymbia citriodora subspecies variegata) in Queensland


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Plantation Establishment

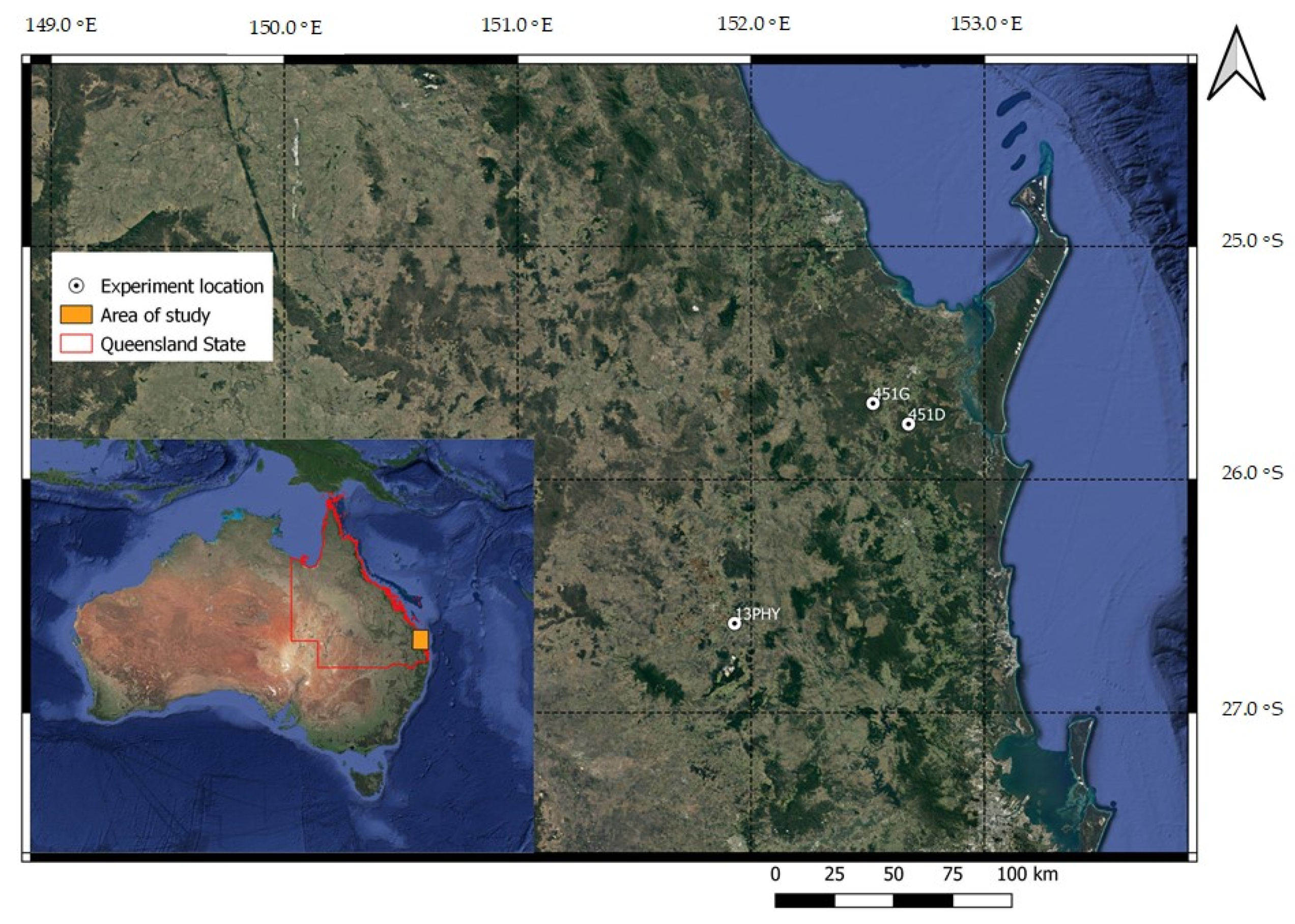{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | 451D | 451G | 13PHY |
|---|---|---|---|
| Planting year | 2000 | 2002 | 2001 |
| Latitude | 25°45′45″ S | 25°40′24″ S | 26°37′4″ S |
| Longitude | 152°40′30″ E | 152°31′22″ E | 151°55′46″ E |
| Mean annual rainfall () | 1111 | 949 | 725 |
| Initial stocking () Current sampling ) | 1000 206 | 1111 240 | 1000 300 |
| Soil type [38] | Grey Kurosol | Red Ferrosol | Red Ferrosol and Brown Dermosol |
| Initial spacing (m) | 5 × 2 | 5 × 1.8 | 5 × 2 |
2.2. Sample Size
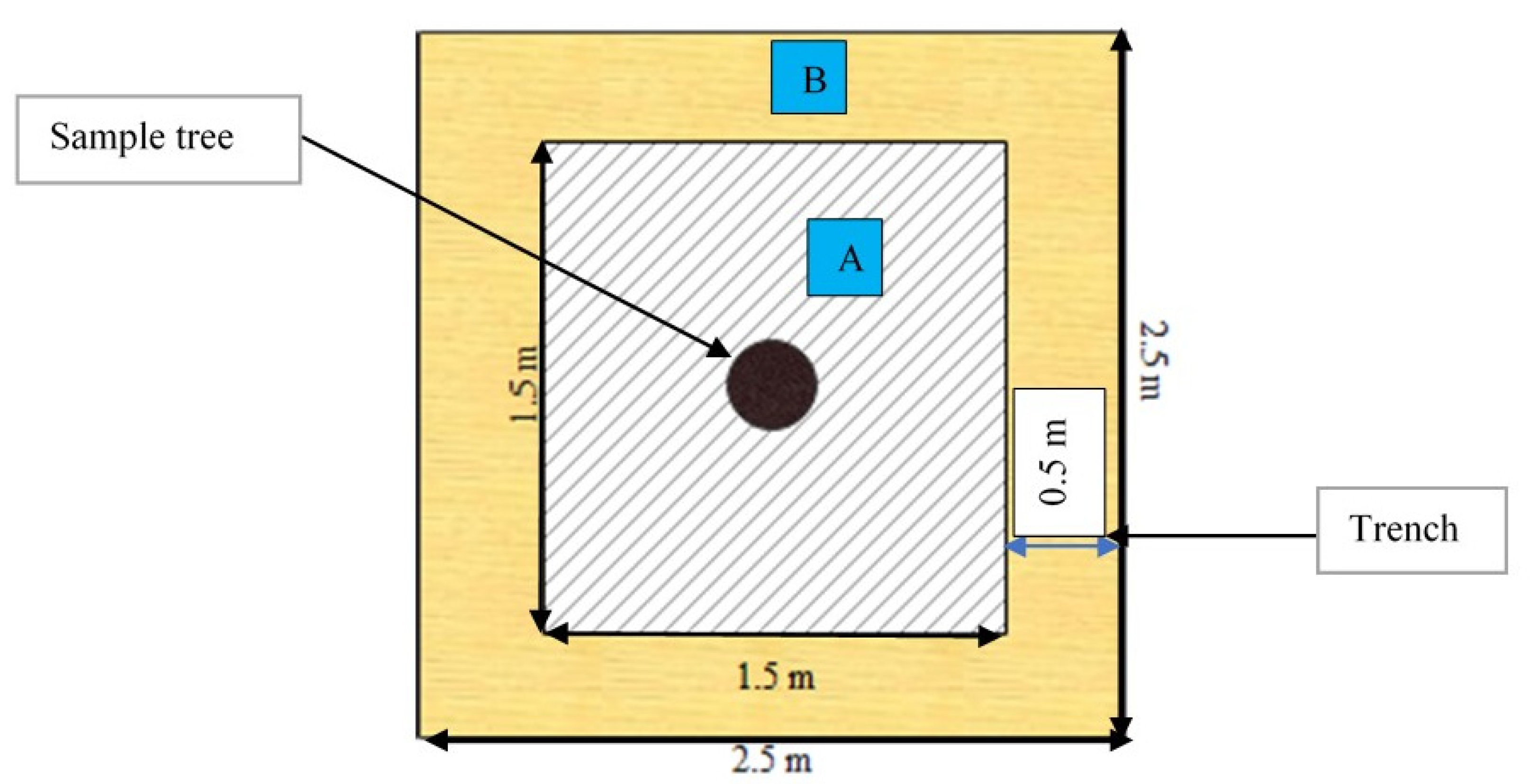
2.3. Destructive Sampling Procedure
2.4. Data Analysis
2.4.1. Biomass Model Development
2.4.2. Model Comparison and Selection
2.4.3. Model Cross Validation
3. Results
3.1. Descriptive Statistics
3.2. Regression Equations Fitted to Natural Log Transformed Data
3.3. Weighted Nonlinear Maximum Likelihood Models
3.4. Model Comparison and Selection
3.5. Model Cross Validation
4. Discussion
4.1. Model Fitting and Cross Validation
4.2. Predictors for BGB Models
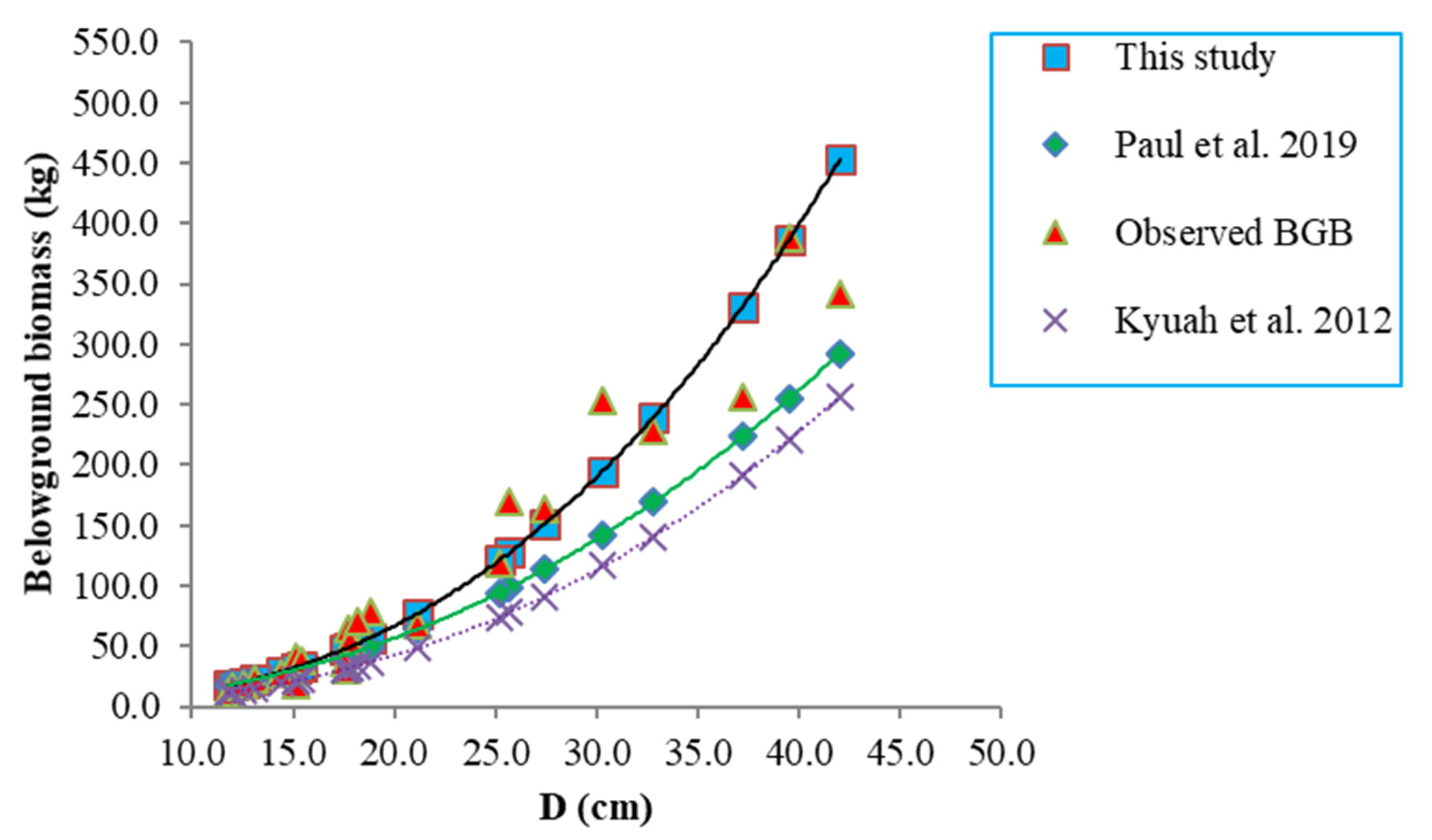
4.3. Biomass Model Comparisons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, S.; Sathaye, J.; Cannell, M.; Kauppi, P.E. Management of Forests for Mitigation of Greenhouse Gas Emissions; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Houghton, R.A.; Davidson, E.A.; Woodwell, G.M. Missing sinks, feedbacks, and understanding the role of terrestrial ecosystems in the global carbon balance. Glob. Biogeochem. Cycles 1998, 12, 25–34. [Google Scholar] [CrossRef]
- Houghton, R.A. Aboveground forest biomass and the global carbon balance. Glob. Chang. Biol. 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Grassi, G.; House, J.; Dentener, F.; Federici, S.; den Elzen, M.; Penman, J. The key role of forests in meeting climate targets requires science for credible mitigation. Nat. Clim. Chang. 2017, 7, 220–226. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Forest; Forest, Biodiversity and People; FAO: Rome, Italy, 2020; ISBN 978-92-5-132419-6. [Google Scholar]
- Canadell, J.G.; Kirschbaum, M.U.; Kurz, W.A.; Sanz, M.J.; Schlamadinger, B.; Yamagata, Y. Factoring out natural and indirect human effects on terrestrial carbon sources and sinks. Environ. Sci. Policy 2007, 10, 370–384. [Google Scholar] [CrossRef]
- van Niekerk, P.B.; Drew, D.M.; Dovey, S.B.; Du Toit, B. Allometric relationships to predict aboveground biomass of 8–10-year-old Eucalyptus grandis × E. nitens in south-eastern Mpumalanga, South Africa. South. For. J. For. Sci. 2020, 82, 15–23. [Google Scholar] [CrossRef]
- Eamus, D.; Chen, X.; Kelley, G.; Hutley, L.B. Root biomass and root fractal analyses of an open Eucalyptus forest in a savanna of north Australia. Aust. J. Bot. 2002, 50, 31–41. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Guidelines for national greenhouse gas inventories. In Agriculture, Forestry and Other Land Use; IPCC: Geneva, Switzerland, 2006; Volume 4, pp. 1–66. [Google Scholar]
- Applegate, G.B. Biomass of Blackbutt (Euclayptus pilularis Sm.) Forests on Fraser Island. Master’s Thesis, University of New England, Armadale, Australia, 1982. [Google Scholar]
- Wildy, D.T.; Pate, J.S. Quantifying above-and below-ground growth responses of the western Australian oil mallee, Eucalyptus kochii subsp. plenissima, to contrasting decapitation regimes. Ann. Bot. 2002, 90, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Grant, J.C.; Nichols, J.D.; Yao, R.L.; Smith, R.G.B.; Brennan, P.D.; Vanclay, J.K. Depth distribution of roots of Eucalyptus dunnii and Corymbia citriodora subsp. variegata in different soil conditions. For. Ecol. Manag. 2012, 269, 249–258. [Google Scholar] [CrossRef]
- Macinnis-Ng, C.M.O.; Fuentes, S.; O’Grady, A.P.; Palmer, A.R.; Taylor, D.; Whitley, R.J.; Yunusa, I.; Zeppel, M.J.B.; Eamus, D. Root biomass distribution and soil properties of an open woodland on a duplex soil. Plant Soil 2010, 327, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Brassard, B.W.; Chen, H.Y.; Bergeron, Y.; Paré, D. Coarse root biomass allometric equations for Abies balsamea, Picea mariana, Pinus banksiana, and Populus tremuloides in the boreal forest of Ontario, Canada. Biomass Bioenergy 2011, 35, 4189–4196. [Google Scholar] [CrossRef]
- Keller, M.; Palace, M.; Hurtt, G. Biomass estimation in the Tapajos National Forest, Brazil: Examination of sampling and allometric uncertainties. For. Ecol. Manag. 2001, 154, 371–382. [Google Scholar] [CrossRef]
- Finér, L.; Helmisaari, H.S.; Lõhmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konôpka, B.; et al. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Yuen, J.Q.; Fung, T.; Ziegler, A.D. Review of allometric equations for major land covers in SE Asia: Uncertainty and implications for above-and below-ground carbon estimates. For. Ecol. Manag. 2016, 360, 323–340. [Google Scholar] [CrossRef]
- Snowdon, P.; Eamus, D.; Gibbons, P.; Keith, H.; Raison, J.; Kirschbaum, M. Synthesis of Allometrics, Review of Root Biomass, and Design of Future Woody Biomass Sampling Strategies; Australian Greenhouse Office: Canberra, Australia, 2000. [Google Scholar]
- Paul, K.I.; Larmour, J.; Specht, A.; Zerihun, A.; Ritson, P.; Roxburgh, S.H.; Sochacki, S.; Lewis, T.; Barton, C.V.; England, J.R.; et al. Testing the generality of below-ground biomass allometry across plant functional types. For. Ecol. Manag. 2019, 432, 102–114. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, W.; Alice, F.E.; Rey-Benayas, J.M. Carbon accumulation in aboveground and belowground biomass and soil of different age native forest plantations in the humid tropical lowlands of Costa Rica. New For. 2012, 43, 197–211. [Google Scholar] [CrossRef]
- Razakamanarivo, R.H.; Razakavololona, A.; Razafindrakoto, M.A.; Vieilledent, G.; Albrecht, A. Below-ground biomass production and allometric relationships of eucalyptus coppice plantation in the central highlands of Madagascar. Biomass Bioenergy 2012, 45, 1–10. [Google Scholar] [CrossRef]
- Waring, B.G.; Powers, J.S. Overlooking what is underground: Root: Shoot ratios and coarse root allometric equations for tropical forests. For. Ecol. Manag. 2017, 385, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Keith, H.; Lindenmayer, D.B.; Mackey, B.G.; Blair, D.; Carter, L.; McBurney, L.; Okada, S.; Konishi-Nagano, T. Accounting for biomass carbon stock change due to wildfire in temperate forest landscapes in Australia. PLoS ONE 2014, 9, e107126. [Google Scholar] [CrossRef] [Green Version]
- Basuki, T.M.; Van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Paul, K.I.; Larmour, J.S.; Roxburgh, S.H.; England, J.R.; Davies, M.J.; Luck, H.D. Measurements of stem diameter: Implications for individual-and stand-level errors. Environ. Monit. Assess. 2017, 189, 416. [Google Scholar] [CrossRef] [PubMed]
- Fayolle, A.; Ngomanda, A.; Mbasi, M.; Barbier, N.; Bocko, Y.; Boyemba, F.; Couteron, P.; Fonton, N.; Kamdem, N.; Katembo, J.; et al. A regional allometry for the Congo basin forests based on the largest ever destructive sampling. For. Ecol. Manag. 2018, 430, 228–240. [Google Scholar] [CrossRef]
- Bates, D.M. Lme4: Mixed-Effects Modeling with R. 2010. Available online: http://lme4.r-forge.r-project.org/book (accessed on 8 April 2015).
- Eamus, D.; Burrows, W.; McGuinness, K. Review of Allometric Relationships for Estimating Woody Biomass for Queensland, The Northern Territory and Western Australia; Australian Greenhouse Office: Canberra, Australia, 2000. [Google Scholar]
- Paul, K.I.; Roxburgh, S.H.; England, J.R.; Brooksbank, K.; Larmour, J.S.; Ritson, P.; Wildy, D.; Sudmeyer, R.; Raison, R.J.; Hobbs, T.; et al. Root biomass of carbon plantings in agricultural landscapes of Southern Australia: Development and testing of allometrics. For. Ecol. Manag. 2014, 318, 216–227. [Google Scholar] [CrossRef]
- McMahon, L.; George, B.; Hean, R. Corymbia maculata, Corymbia citriodora subsp, variegata and Corymbia henryi; Industry and Investment, New South Wales Government: Sydney, Australia, 2010. [Google Scholar]
- Lee, D.J. Achievements in forest tree genetic improvement in Australia and New Zealand 2: Development of Corymbia species and hybrids for plantations in eastern Australia. Aust. For. 2007, 70, 11–16. [Google Scholar] [CrossRef]
- Lee, D.J.; Huth, J.R.; Osbourne, D.O.; Hogg, B.W. Selecting hardwood varieties for fibre production in Queensland’s subtropics. In 2nd Australasian Forest Genetics Conference: Book of Abstracts; Forest Products Commission: Kensington, Australia, 2009. [Google Scholar]
- Lee, D.J.; Brawner, J.; Smith, T.E.; Hogg, B.W.; Meder, R.; Osborne, D.O. Productivity of Plantation Hardwood Tree Species in North-Eastern Australia: A Report from the Forest Adaptation and Sequestration Alliance; Department of Agriculture, Fisheries and Forestry (DAFF), Australia Government: Sydney, Australia, 2011. [Google Scholar]
- Smith, H.J.; Boyton, S.; Henson, M. Developing elite trees for economically viable forest plantations in low rainfall sites. In Proceedings of the ANZIF Conference: Growing Forest Values, Coffs Harbour, NSW, Australia, 3–7 June 2008. [Google Scholar]
- Gardner, R.A.; Little, K.M.; Arbuthnot, A. Wood and fibre productivity potential of promising new eucalypt species for coastal Zululand, South Africa. Aust. For. 2007, 70, 37–47. [Google Scholar] [CrossRef]
- Isbell, R. The Australian Soil Classification; CSIRO Publishing: Clayton, Australia, 2016. [Google Scholar]
- Ovington, J.D.; Forrest, W.G.; Armstrong, J.S. Tree Biomass Estimation; Department of Forestry, Australian National University: Canberra, Australia, 1967. [Google Scholar]
- Snowdon, P.; Raison, R.J.; Keith, H.; Ritson, P.; Grierson, P.; Adams, M.; Montagu, K.; Bi, H.Q.; Burrows, W.; Eamus, D. Protocol for Sampling Tree and Stand. Biomass; Australian Greenhouse Office: Canberra, Australia, 2002. [Google Scholar]
- Huynh, T.; Lee, D.J.; Applegate, G.; Lewis, T. Field methods for above and belowground biomass estimation in plantation forests. MethodsX 2021, 8, 101192. [Google Scholar] [CrossRef]
- Moore, J.R. Allometric equations to predict the total above-ground biomass of radiata pine trees. Ann. For. Sci. 2010, 67, 806. [Google Scholar] [CrossRef] [Green Version]
- Picard, N.; Saint-André, L.; Henry, M. Manual for Building Tree Volume and Biomass Allometric Equations: From Field Measurement to Prediction; FAO, Food and Agricultural Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Huy, B.; Thanh, G.T.; Poudel, K.P.; Temesgen, H. Individual plant allometric equations for estimating aboveground biomass and its components for a common bamboo species (Bambusa procera A. Chev. and A. Camus) in tropical forests. Forests 2019, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- Jara, M.C.; Henry, M.; Réjou-Méchain, M.; Wayson, C.; Zapata-Cuartas, M.; Piotto, D.; Guier, F.A.; Lombis, H.C.; López, E.C.; Lara, R.C.; et al. Guidelines for documenting and reporting tree allometric equations. Ann. For. Sci. 2015, 72, 763–768. [Google Scholar] [CrossRef]
- Ketterings, Q.M.; Coe, R.; van Noordwijk, M.; Palm, C.A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For. Ecol. Manag. 2001, 146, 199–209. [Google Scholar] [CrossRef]
- Xiao, X.; White, E.P.; Hooten, M.B.; Durham, S.L. On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Ecology 2011, 92, 1887–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Cheng, Z.; Gamarra, J.G.P.; Birigazzi, L. Inventory of Allometric Equations for Estimation Tree Biomass—A Database for China; FAO: Rome, Italy, 2014. [Google Scholar]
- Burrows, W.H.; Hoffmann, M.B.; Compton, J.F.; Back, P.V. Allometric Relationships and Community Biomass Stocks in White Cypress Pine (Callitris glaucophylla) and Associated Eucalypts of the Carnarvon Area—South Central Queensland; National Carbon Accounting System Technical Report 33; Australian Greenhouse Office: Canberra, Australia, 2001. [Google Scholar]
- Huy, B.; Kralicek, K.; Poudel, K.P.; Phuong, V.T.; Van Khoa, P.; Hung, N.D.; Temesgen, H. Allometric equations for estimating tree aboveground biomass in evergreen broadleaf forests of Viet Nam. For. Ecol. Manag. 2016, 382, 193–205. [Google Scholar] [CrossRef]
- Montagu, K.D.; Düttmer, K.; Barton, C.V.M.; Cowie, A.L. Developing general allometric relationships for regional estimates of carbon sequestration—An example using Eucalyptus pilularis from seven contrasting sites. For. Ecol. Manag. 2005, 204, 115–129. [Google Scholar] [CrossRef]
- Williams, R.J.; Zerihun, A.; Montagu, K.D.; Hoffman, M.; Hutley, L.B.; Chen, X. Allometry for estimating aboveground tree biomass in tropical and subtropical eucalypt woodlands: Towards general predictive equations. Aust. J. Bot. 2005, 53, 607–619. [Google Scholar] [CrossRef]
- Ximenes, F.A.; Gardner, W.D.; Richards, G.P. Total above-ground biomass and biomass in commercial logs following the harvest of spotted gum (Corymbia maculata) forests of SE NSW. Aust. For. 2006, 69, 213–222. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Baskerville, G.L. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Development Core Team. Linear and Nonlinear Mixed Effects Models. R Package Version. 2007, Volume 3, pp. 1–89. Available online: http://132.180.15.2/math/statlib/R/CRAN/doc/packages/nlme.pdf (accessed on 5 August 2021).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. R Package Version 2(1). 2016. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 5 August 2021).
- Kenzo, T.; Ichie, T.; Hattori, D.; Itioka, T.; Handa, C.; Ohkubo, T.; Kendawang, J.J.; Nakamura, M.; Sakaguchi, M.; Takahashi, N.; et al. Development of allometric relationships for accurate estimation of above-and below-ground biomass in tropical secondary forests in Sarawak, Malaysia. J. Trop. Ecol. 2009, 371–386. [Google Scholar] [CrossRef]
- Roxburgh, S.H.; Paul, K.I.; Clifford, D.; England, J.R.; Raison, R.J. Guidelines for constructing allometric models for the prediction of woody biomass: How many individuals to harvest? Ecosphere 2015, 6, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Temesgen, H.; Zhang, C.H.; Zhao, X.H. Modelling tree height–diameter relationships in multi-species and multi-layered forests: A large observational study from Northeast China. For. Ecol. Manag. 2014, 316, 78–89. [Google Scholar] [CrossRef]
- Zeileis, A.; Hothorn, T. Diagnostic Checking in Regression Relationships. R News 2002, 2, 7–10. Available online: http://CRAN.R-project.org/doc/Rnews/ (accessed on 5 August 2011).
- Furnival, G.M. An index for comparing equations used in constructing volume tables. For. Sci. 1961, 7, 337–341. [Google Scholar] [CrossRef]
- Fordjour, P.A.; Rahmad, Z.B. Development of allometric equation for estimating above-ground liana biomass in tropical primary and secondary forest, Malaysia. Int. J. Ecol. 2013, 2013, 658140. [Google Scholar]
- Picard, R.R.; Cook, R.D. Cross-validation of regression models. J. Am. Stat. Assoc. 1984, 79, 575–583. [Google Scholar] [CrossRef]
- Xu, Q.S.; Liang, Y.Z.; Du, Y.P. Monte Carlo cross-validation for selecting a model and estimating the prediction error in multivariate calibration. J. Chemom. J. Chemom. Soc. 2004, 18, 112–120. [Google Scholar] [CrossRef]
- Fonseca-Delgado, R.; Gómez-Gil, P. An assessment of ten-fold and Monte Carlo cross validations for time series forecasting. In Proceedings of the 2013 10th International Conference on Electrical Engineering, Computing Science and Automatic Control (CCE), Mexico City, Mexico, 30 September–4 October 2013; IEEE: Piscataway, NJ, USA, 2013; pp. 215–220. [Google Scholar] [CrossRef]
- Xu, Y.; Goodacre, R. On splitting training and validation set: A comparative study of cross-validation, bootstrap and systematic sampling for estimating the generalization performance of supervised learning. J. Anal. Test. 2018, 2, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Specht, A.; West, P.W. Estimation of biomass and sequestered carbon on farm forest plantations in northern New South Wales, Australia. Biomass Bioenergy 2003, 25, 363–379. [Google Scholar] [CrossRef]
- Tamang, B.; Andreu, M.G.; Staudhammer, C.L.; Rockwood, D.L.; Jose, S. Equations for estimating aboveground biomass of cadaghi (Corymbia torelliana) trees in farm windbreaks. Agrofor. Syst. 2012, 86, 255–266. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuri, C.; van Noordwijk, M.; Neufeldt, H. Allometry and partitioning of above-and below-ground biomass in farmed eucalyptus species dominant in Western Kenyan agricultural landscapes. Biomass Bioenergy 2013, 55, 276–284. [Google Scholar] [CrossRef]
- Saint-André, L.; M’Bou, A.T.; Mabiala, A.; Mouvondy, W.; Jourdan, C.; Roupsard, O.; Deleporte, P.; Hamel, O.; Nouvellon, Y. Age-related equations for above-and below-ground biomass of a Eucalyptus hybrid in Congo. For. Ecol. Manag. 2005, 205, 199–214. [Google Scholar] [CrossRef]
- Herrero, C.; Juez, L.; Tejedor, C.; Pando, V.; Bravo, F. Importance of root system in total biomass for Eucalyptus globulus in northern Spain. Biomass Bioenergy 2014, 67, 212–222. [Google Scholar] [CrossRef]
- Huy, B.; Tinh, N.T.; Poudel, K.P.; Frank, B.M.; Temesgen, H. Taxon-specific modeling systems for improving reliability of tree aboveground biomass and its components estimates in tropical dry dipterocarp forests. For. Ecol. Manag. 2019, 437, 156–174. [Google Scholar] [CrossRef]
- Paul, K.I.; Radtke, P.J.; Roxburgh, S.H.; Larmour, J.; Waterworth, R.; Butler, D.; Brooksbank, K.; Ximenes, F. Validation of allometric biomass models: How to have confidence in the application of existing models. For. Ecol. Manag. 2018, 412, 70–79. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuri, C.; Jamnadass, R.; Mwangi, P.; Coe, R.; Neufeldt, H. Allometric equations for estimating biomass in agricultural landscapes: II. Belowground biomass. Agric. Ecosyst. Environ. 2012, 158, 225–234. [Google Scholar] [CrossRef]
- Resh, S.C.; Battaglia, M.; Worledge, D.; Ladiges, S. Coarse root biomass for eucalypt plantations in Tasmania, Australia: Sources of variation and methods for assessment. Trees 2003, 17, 389–399. [Google Scholar] [CrossRef]
- Ruark, G.A.; Bockheim, J.G. Below-ground biomass of 10-, 20-, and 32-year-old Populus tremuloides in Wisconsin. Pedobiologia 1987, 30, 201. [Google Scholar]
- Marziliano, P.A.; Lafortezza, R.; Medicamento, U.; Lorusso, L.; Giannico, V.; Colangelo, G.; Sanesi, G. Estimating belowground biomass and root/shoot ratio of Phillyrea latifolia L. in the Mediterranean forest landscapes. Ann. For. Sci. 2015, 72, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Millikin, C.S.; Bledsoe, C.S. Biomass and distribution of fine and coarse roots from blue oak (Quercus douglasii) trees in the northern Sierra Nevada foothills of California. Plant Soil 1999, 214, 27–38. [Google Scholar] [CrossRef]
- Misra, R.K.; Turnbull, C.R.A.; Cromer, R.N.; Gibbons, A.K.; LaSala, A.V. Below-and above-ground growth of Eucalyptus nitens in a young plantation: I. Biomass. For. Ecol. Manag. 1998, 106, 283–293. [Google Scholar] [CrossRef]
- DesRochers, A.; Lieffers, V.J. The coarse-root system of mature Populus tremuloides in declining stands in Alberta, Canada. J. Veg. Sci. 2001, 12, 355–360. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Johnsen, K.; Stokes, T. Production, allocation, and stemwood growth efficiency of Pinus taeda L. stands in response to 6 years of intensive management. For. Ecol. Manag. 2004, 192, 59–70. [Google Scholar] [CrossRef]
- Lavigne, M.B.; Krasowski, M.J. Estimating coarse root biomass of balsam fir. Can. J. For. Res. 2007, 37, 991–998. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]




| Location (Age) | N | D (cm) | H (m) | RB (kg) | MR (kg) | BGB (kg) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Min. | Max. | Min. | Max. | Min. | Max. | Min. | Max. | Min. | Max. | ||
| 451D (20) | 11 | 17.7 | 42.0 | 20.2 | 32.0 | 48.0 | 319.8 | 16.4 | 67.7 | 64.4 | 387.6 |
| 451D (9) | 3 | 12.0 | 17.8 | 16.5 | 17.5 | 13.1 | 49.6 | 2.8 | 5.8 | 16.0 | 55.4 |
| 451G (7) | 3 | 11.8 | 17.6 | 16.6 | 20.4 | 10.3 | 29.1 | 1.0 | 2.9 | 11.2 | 30.3 |
| 13 PHY (8) | 6 | 12.5 | 18.2 | 13.1 | 16.4 | 15.6 | 51.9 | 0.7 | 18.4 | 18.1 | 70.2 |
| Total | 23 | 11.8 | 42.0 | 13.1 | 32.0 | 10.3 | 319.8 | 0.7 | 67.7 | 11.2 | 387.6 |
| Equation No. | Model Form | Parameter Estimates | CF | Weight Variable | AIC | Adj. R2 | Bias | RMSE | MAPE | FI | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| α | β | ||||||||||
| Logarithmic transformed models | |||||||||||
| (3) | ln(BGB) = ln(α) + β × ln(D) | 0.02354 | 2.64328 | 1.035 | - | 234.2 | 0.875 | −6.3787 | 37.738 | 22.179 | 16.306 |
| (4) | ln(BGB) = ln(α) + β × ln(H) | 0.00302 | 3.32052 | 1.096 | - | 227.9 | 0.905 | −1.9043 | 32.876 | 39.416 | 26.668 |
| (5) | ln(BGB) = ln(α) + β × ln(DH) | 0.00757 | 1.50922 | 1.047 | - | 222.5 | 0.928 | −4.2685 | 27.987 | 25.677 | 18.898 |
| (6) | ln(BGB) = ln(α) + β × ln(D2H) | 0.01101 | 0.96482 | 1.040 | - | 226.9 | 0.913 | −5.0478 | 30.780 | 23.487 | 17.478 |
| (7) | ln(BGB) = ln(α) + β × ln(DH2) | 0.00531 | 1.04515 | 1.058 | - | 220.5 | 0.934 | −3.4478 | 26.824 | 29.197 | 20.918 |
| Weighted nonlinear models | |||||||||||
| (11) | BGB = α × D β | 0.02933 | 2.5805 | - | 1/D δ | 206.9 | 0.903 | −0.00003 | 0.014 | 0.029 | 0.014 |
| (12) | BGB = α × H β | 0.00269 | 3.3789 | - | 1/H δ | 226.8 | 0.906 | 0.00093 | 0.065 | 0.132 | 0.066 |
| (13) | BGB = α × (DH) β | 0.01150 | 1.44752 | - | 1/(DH) δ | 209.9 | 0.944 | −0.00048 | 0.058 | 0.120 | 0.058 |
| (14) | BGB = α × (D2H) β | 0.01687 | 0.92222 | - | 1/(D2H) δ | 207.5 | 0.939 | −0.00033 | 0.037 | 0.079 | 0.037 |
| (15) | BGB = α × (DH2) β | 0.00722 | 1.01661 | - | 1/(DH2) δ | 214.0 | 0.940 | −0.00016 | 0.077 | 0.158 | 0.777 |
| Equation No. | Model Form | AIC | Adj. R2 | Bias | RMSE | MAPE |
|---|---|---|---|---|---|---|
| (11) | BGB = α × D β | 146.4 | 0.854 | 0.040 | 0.063 | 0.090 |
| (12) | BGB = α × H β | 160.9 | 0.916 | 0.422 | 2.631 | 1.213 |
| (13) | BGB = α × (DH) β | 149.0 | 0.921 | 0.194 | 0.308 | 0.346 |
| (14) | BGB = α × (D2H) β | 147.6 | 0.910 | 0.136 | 0.175 | 0.224 |
| (15) | BGB = α × (DH2) β | 151.6 | 0.922 | 0.223 | 0.748 | 0.574 |
| Reference | Forest Type | Site | Species (Diameter Range, cm) | Bias | RMSE | MAPE |
|---|---|---|---|---|---|---|
| This study | P | Australia | Corymbia citriodora subsp. variegata (11.8–42) | −0.003 | 0.014 | 0.029 |
| Paul et al. (2019) | N, P | Australia | Mixed Eucalyptus spp. (1.1–139) | 8.4 | 29.6 | 26.5 |
| Kuyah et al. (2012) | P | Kenya | Eucalyptus spp. (3–102) | 32.2 | 42.2 | 34.2 |
| Eamus et al. (2002) | N | Australia | Eucalyptus spp. (3–25) | 8.4 | 58.5 | 42.2 |
| Resh et al. (2003) | P | Australia | E. globulus and E. nitens (10–25) | 61.2 | 78.0 | 61.2 |
| Saint-André et al. (2005) | P | Congo | E. alba (3–25) | 66.9 | 85.7 | 67.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huynh, T.; Applegate, G.; Lewis, T.; Pachas, A.N.A.; Hunt, M.A.; Bristow, M.; Lee, D.J. Species-Specific Allometric Equations for Predicting Belowground Root Biomass in Plantations: Case Study of Spotted Gums (Corymbia citriodora subspecies variegata) in Queensland. Forests 2021, 12, 1210. https://doi.org/10.3390/f12091210
Huynh T, Applegate G, Lewis T, Pachas ANA, Hunt MA, Bristow M, Lee DJ. Species-Specific Allometric Equations for Predicting Belowground Root Biomass in Plantations: Case Study of Spotted Gums (Corymbia citriodora subspecies variegata) in Queensland. Forests. 2021; 12(9):1210. https://doi.org/10.3390/f12091210
Chicago/Turabian StyleHuynh, Trinh, Grahame Applegate, Tom Lewis, Anibal Nahuel A. Pachas, Mark A. Hunt, Mila Bristow, and David J. Lee. 2021. "Species-Specific Allometric Equations for Predicting Belowground Root Biomass in Plantations: Case Study of Spotted Gums (Corymbia citriodora subspecies variegata) in Queensland" Forests 12, no. 9: 1210. https://doi.org/10.3390/f12091210
APA StyleHuynh, T., Applegate, G., Lewis, T., Pachas, A. N. A., Hunt, M. A., Bristow, M., & Lee, D. J. (2021). Species-Specific Allometric Equations for Predicting Belowground Root Biomass in Plantations: Case Study of Spotted Gums (Corymbia citriodora subspecies variegata) in Queensland. Forests, 12(9), 1210. https://doi.org/10.3390/f12091210