


Allocation of Phosphorus Fractions in Chinese Fir in Response to Low Phosphorus Availability Using 32P Tracer

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Study Design
2.3. Determination Methods
2.3.1. Determination of Dry Weight
2.3.2. Radioactivity Determination of Total Exogenous P and Different P Fractions in Plantlets
2.3.3. Determination of Specific Activity
2.4. Data Analyses
3. Results
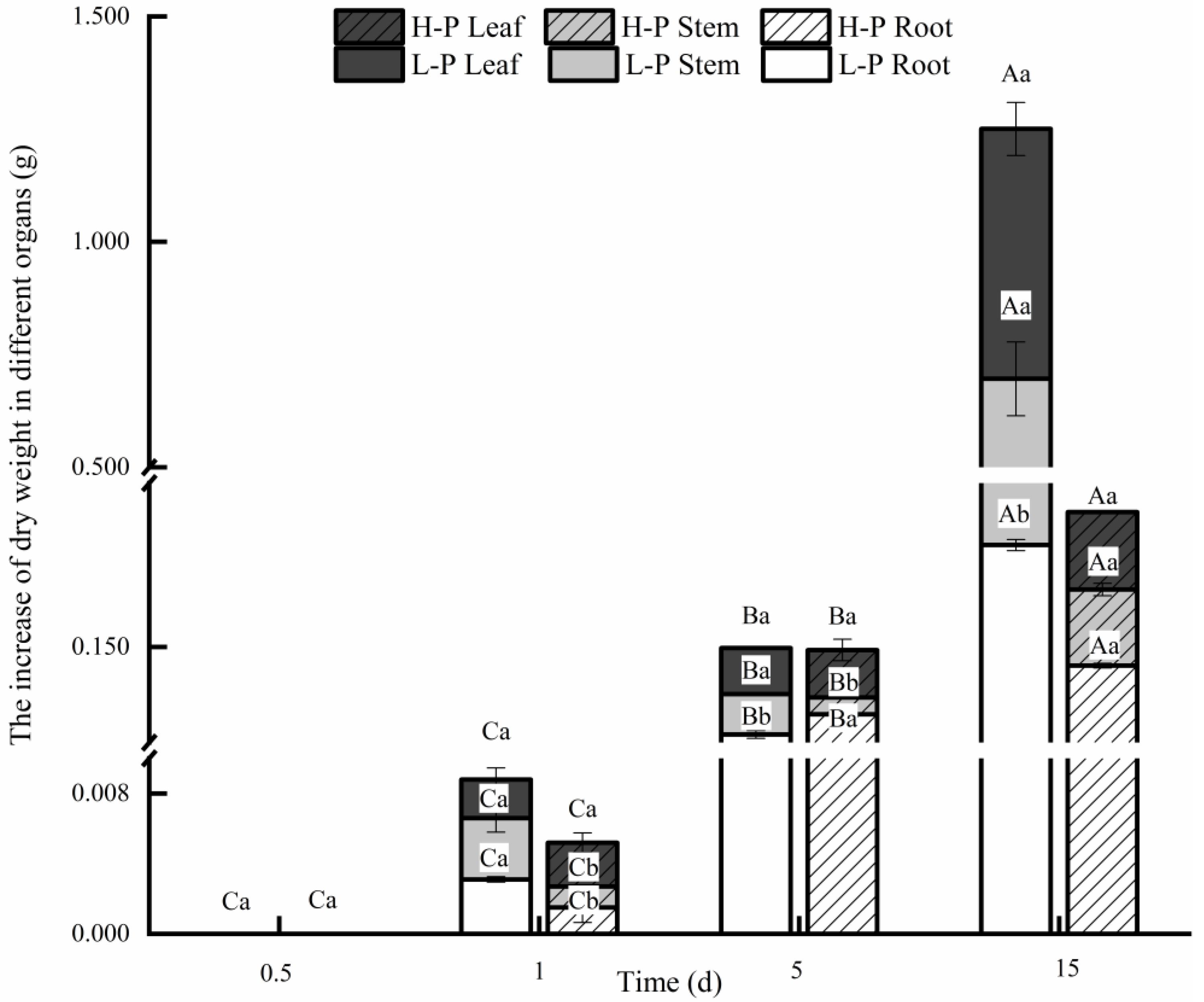
3.1. Comparison of the Increase in Dry Weight after Different P Treatments
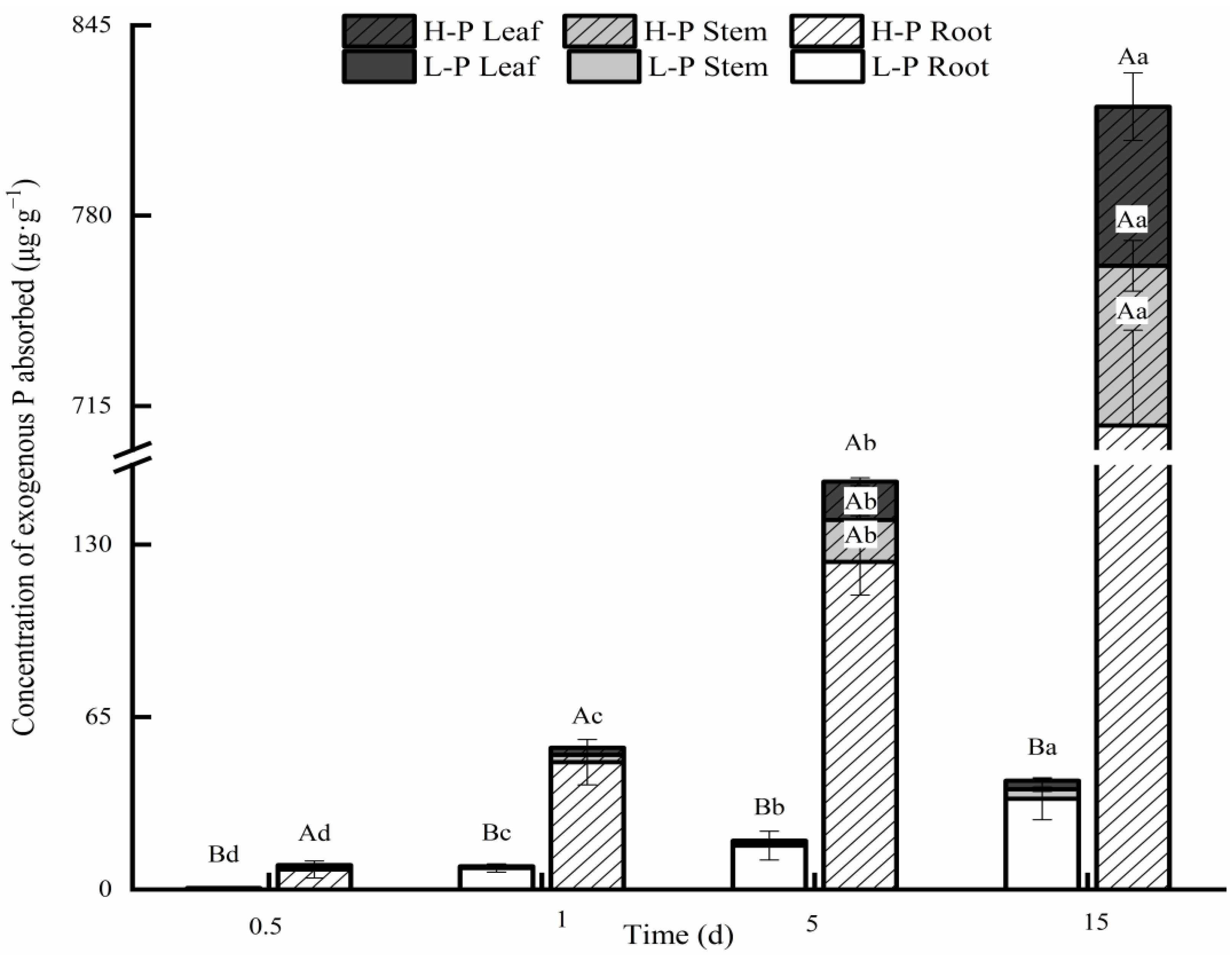
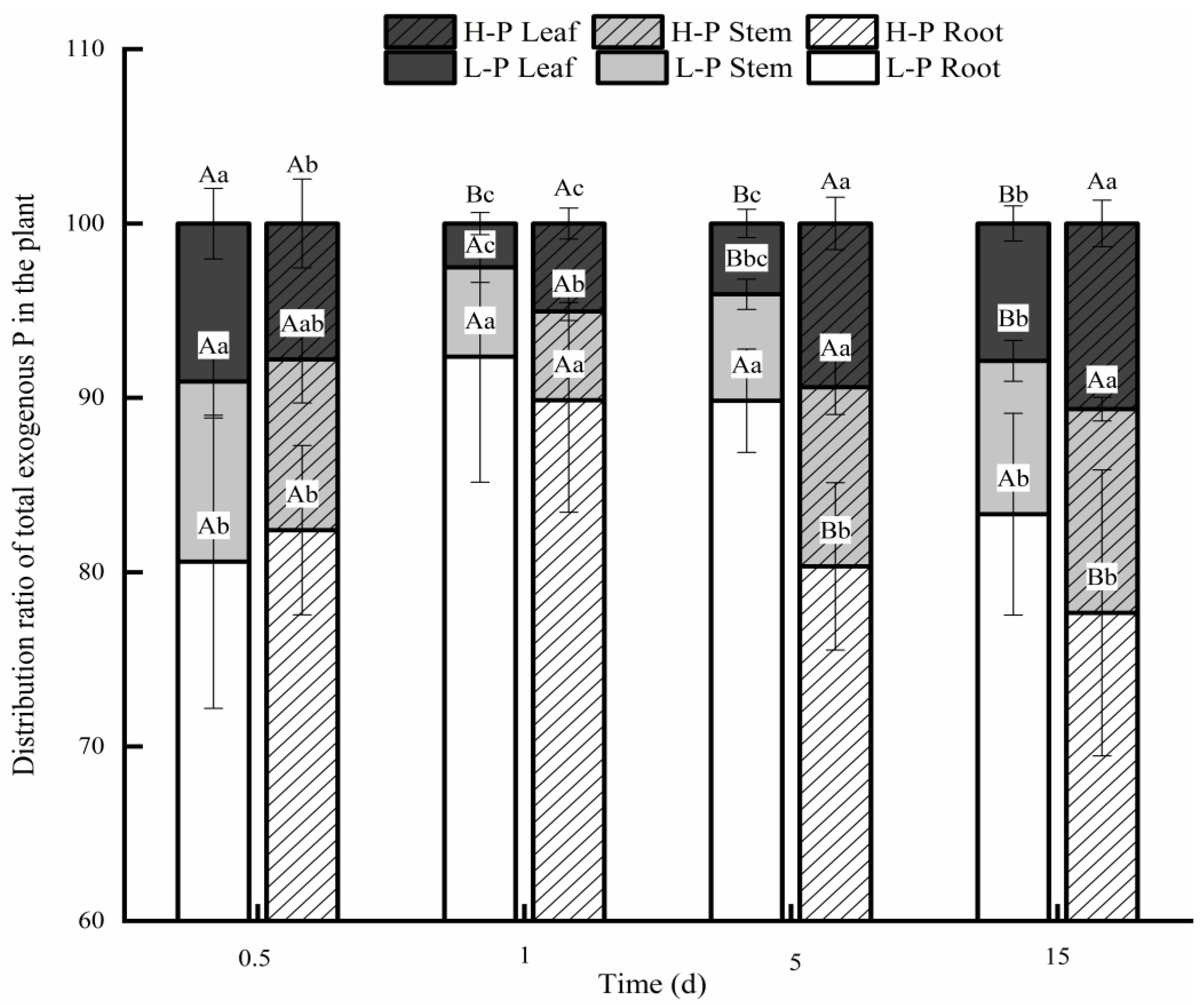
3.2. Comparison of Exogenous P Distributions after Different P Treatments
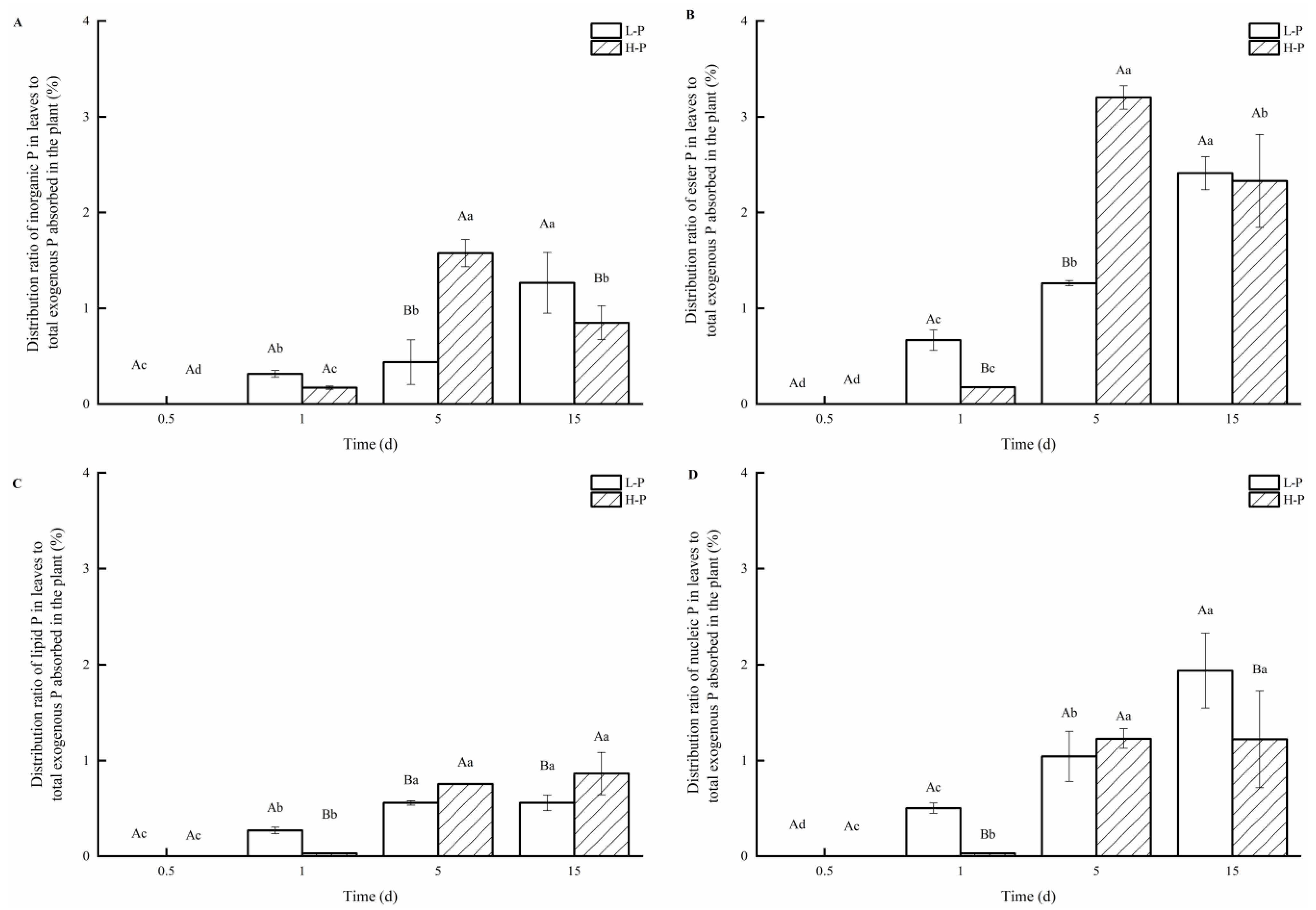
3.3. Distribution of P Fractions in Leaves under Different P Treatments
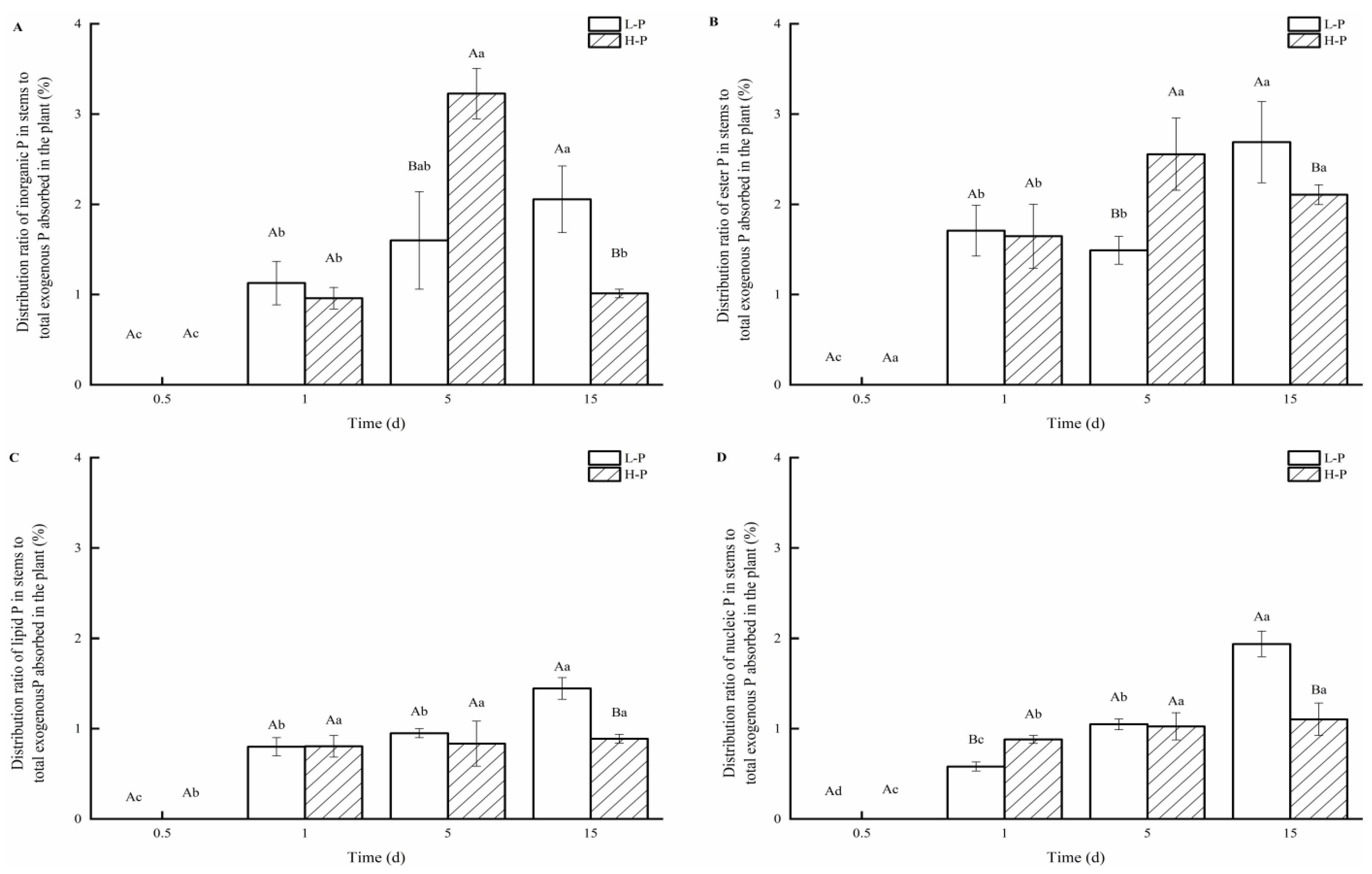
3.4. Distribution of P Fractions in Stems under Different P Treatments
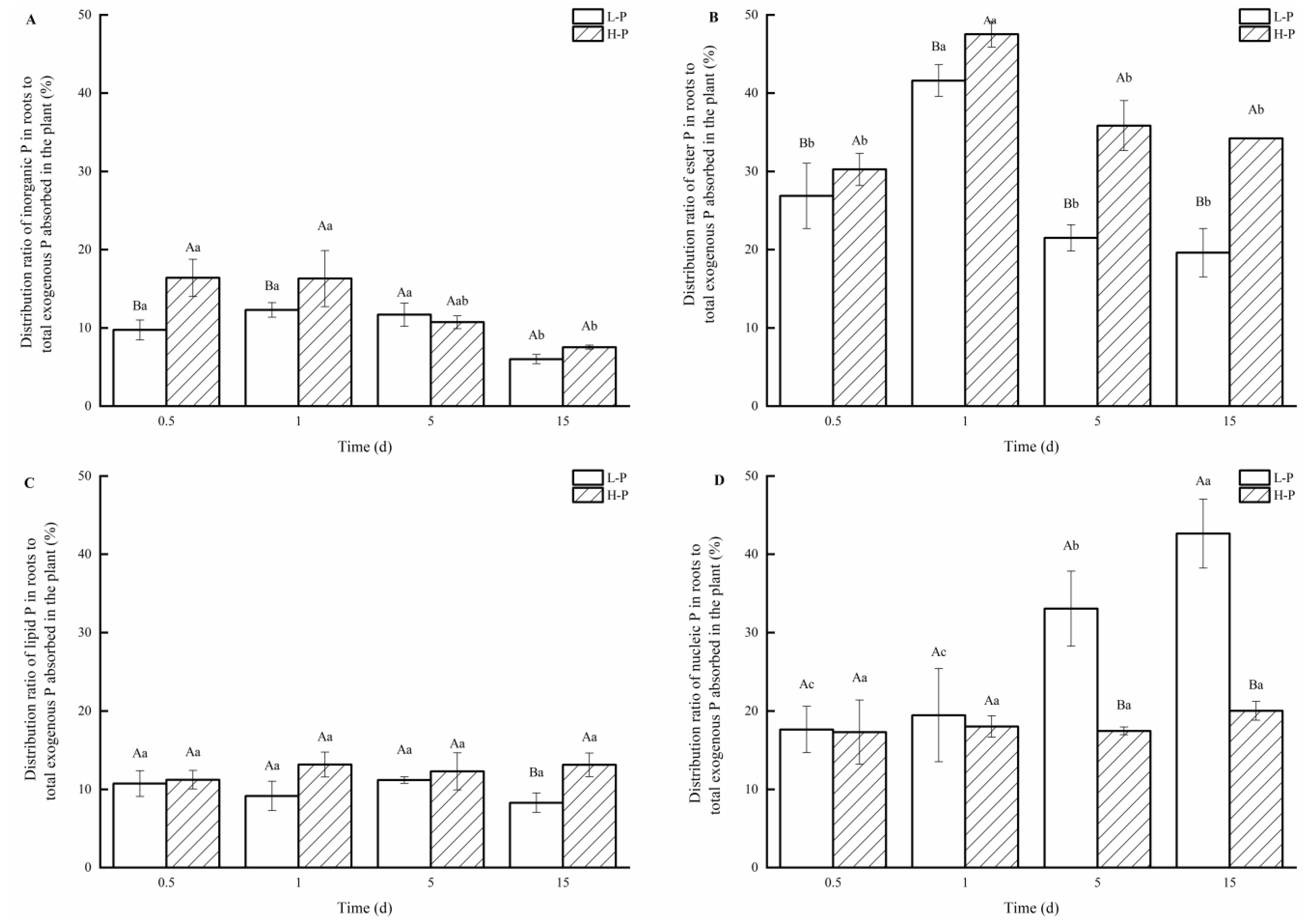
3.5. Distribution of P Fractions in Roots under Different P Treatments
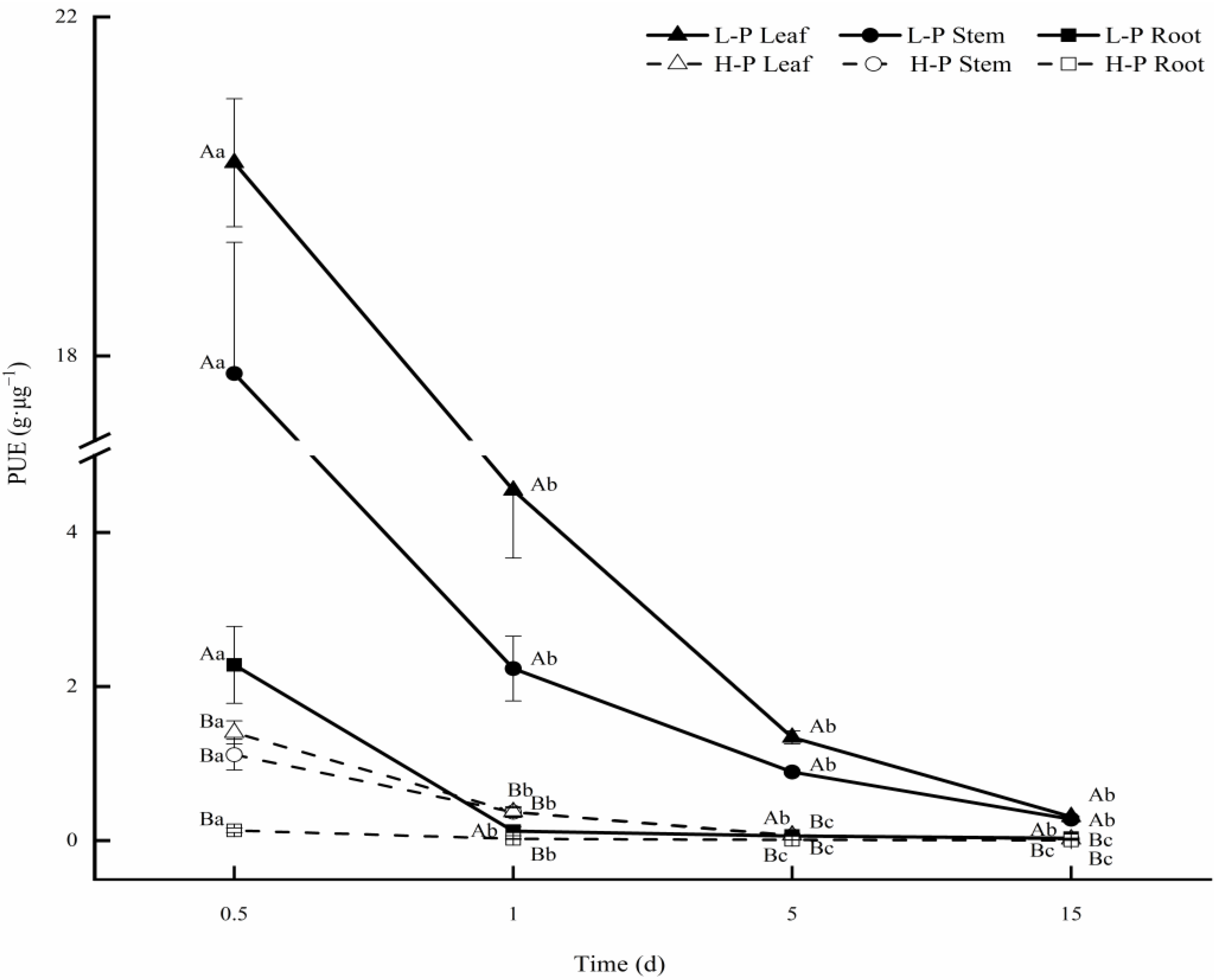
3.6. PUE in Different Organs under Different P Treatments
3.7. Relationship between P Fractions and PUE in Different Organs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambers, H.; Brundrett, M.C.; Raven, J.A.; Hopper, S.D. Plant mineral nutrition in ancient landscapes: High plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 2011, 348, 7–27. [Google Scholar] [CrossRef]
- Ragothama, K.G.; Karthikeyan, A.S. Phosphate acquisition. Plant Soil 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Fink, J.R.; Inda, A.V.; Bayer, C.; Torrent, J.; Barrón, V. Mineralogy and phosphorus adsorption in soils of south and central-west Brazil under conventional and no-tillage systems. Acta Sci. Agron. 2014, 36, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Bortoluzzi, E.C.; Pérez, C.A.S.; Ardisson, J.D.; Tiecher, T.; Caner, L. Occurrence of iron and aluminum sesquioxides and their implications for the P sorption in subtropical soils. Appl. Clay Sci. 2015, 104, 196–204. [Google Scholar] [CrossRef]
- Freitas, E.C.S.D.; Paiva, H.N.D.; Leite, H.G.; Oliveira Neto, S.N.D. Effect of phosphate fertilization and base saturation of substrate on the seedlings growth and quality of Plathymenia foliolosa Benth. Rev. Árvore 2017, 41, e410111. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P. Root phenes that reduce the metabolic costs of soil exploration: Opportunities for 21st century agriculture. Plant Cell Environ. 2014, 38, 1775–1784. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Huang, C.Y.; Shirley, N.; Genc, Y.; Shi, B.; Langridge, P. Phosphate utilization efficiency correlates with expression of low-affinity phosphate transporters and noncoding RNA, IPS1, in Barley. Plant Physiol. 2011, 156, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Hammond, J.P.; Broadley, M.R.; White, P.J.; King, G.J.; Bowen, H.C.; Hayden, R.; Meacham, M.C.; Mead, A.; Overs, T.; Spracklen, W.P.; et al. Shoot yield drives phosphorus use efficiency in Brassica oleracea and correlates with root architecture traits. J. Exp. Bot. 2009, 60, 1953–1968. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Liang, H.L.; Xu, F.S.; Wang, Y.H. Genotypic variation in phosphorus fractions and its relation to phosphorus efficiency in seedlings of Brassica napus L. Plant Nutr. Fert. Sci. 2008, 14, 351–356. [Google Scholar] [CrossRef]
- Rouached, H.; Stefanovic, A.; Secco, D.; Arpat, A.B.; Gout, E.; Bligny, R.; Bligny, R.; Poirier, Y. Uncoupling phosphate deficiency from its major effects on growth and transcriptome via PHO1 expression in Arabidopsis. Plant J. 2011, 65, 557–570. [Google Scholar] [CrossRef]
- Hidaka, A.; Kitayama, K. Allocation of foliar phosphorus fractions and leaf traits of tropical tree species in response to decreased soil phosphorus availability on Mount Kinabalu, Borneo. J. Ecol. 2011, 99, 849–857. [Google Scholar] [CrossRef]
- Piccin, R.; Couto, R.D.R.; Bellinaso, R.J.S.; Gatiboni, L.C.; Conti, L.D.; Rodrigues, L.A.T.; Somavilla, L.M.; Brunetto, G. Phosphorus forms in leaves and their relationships with must composition and yield in grapevines. Pesqui. Agropecu. Bras. 2017, 52, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Kulmann, M.S.S.; Stefanello, L.O.S.; Schwalbert, R.A.; Berghetti, L.L.P.; Brunetto, G. Effects of phosphorus fertilizer application on phosphorus fractions in different organs of Cordia trichotoma. J. For. Res. 2020, 32, 725–732. [Google Scholar] [CrossRef]
- Elser, J.; Bennett, E. Phosphorus cycle: A broken biogeochemical cycle. Nature 2011, 478, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Wieneke, J. Phosphorus efficiency and phosphorus remobilization in two sorghum (Sorghum bicolor (L.) Moench) cultivars. Plant Soil 1990, 123, 139–145. [Google Scholar] [CrossRef]
- Moro, H.; Park, H.D.; Kunito, T. Organic phosphorus substantially contributes to crop plant nutrition in soils with low phosphorus availability. Agronomy 2021, 11, 903. [Google Scholar] [CrossRef]
- Mayers, X.G.; Turner, B.L.; Laliberté, E. Greater root phosphatase activity of tropical trees at low phosphorus despite strong variation among species. Ecology 2020, 101, e03090. [Google Scholar] [CrossRef]
- Wang, F.C.; Fang, X.M.; Wang, G.G.; Rong, M.; Lin, X.F.; Wang, H.M.; Chen, F.S. Effects of nutrient addition on foliar phosphorus fractions and their resorption in different-aged leaves of Chinese fir in subtropical China. Plant Soil 2019, 443, 41–54. [Google Scholar] [CrossRef]
- Fan, S.H.; Shen, W.T.; Ma, X.Q.; Lin, K.M.; Zhang, X.Q. Effect of successive planting on productivity of Chinese fir of different age plantations. For. Res. 2003, 16, 560–567. [Google Scholar] [CrossRef]
- Wu, P.F. Adaptation Mechanism of Chinese Fir Clones with High Phosphorus-Use-Efficiency to Envrironmental Phosphorus Stress. Ph.D. Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2009. [Google Scholar]
- Wu, P.F.; Wang, G.Y.; Farooq, T.H.; Li, Q.; Zou, X.H.; Ma, X.Q. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? J. Soils Sediments 2017, 17, 2775–2785. [Google Scholar] [CrossRef]
- Wu, P.F.; Lai, H.Y.; Mulualem, T.; Wu, W.; Wang, P.; Wang, G.Y.; Ma, X.Q. Does phosphorus deficiency induce formation of root cortical aerenchyma maintaining growth of Cunninghamia lanceolata? Trees 2018, 32, 1633–1642. [Google Scholar] [CrossRef]
- Zou, X.H.; Wei, D.; Wu, P.F.; Zhang, Y.; Hu, Y.N.; Chen, S.T.; Ma, X.Q. Strategies of organic acid production and exudation in response to low-P stress in Chinese fir genotypes differing in P-use efficiencies. Trees 2018, 32, 897–912. [Google Scholar] [CrossRef]
- Chen, W.T.; Zhou, M.Y.; Zhao, M.Z.; Chen, R.H.; Mulualem, T.; Wu, P.F.; Li, M.; Ma, X.Q. Transcriptome analysis provides insights into the root response of Chinese fir to phosphorus deficiency. BMC Plant Biol. 2021, 21, 525. [Google Scholar] [CrossRef]
- Lai, H.Y.; Wu, K.; Wang, N.M.; Wu, W.J.; Wu, P.F. Relationship between volatile organic compounds released and growth of Cunninghamia lanceolata roots under low-P conditions. iForest 2018, 11, 713–720. [Google Scholar] [CrossRef]
- Liang, X.; Liu, A.Q.; Ma, X.Q.; Feng, L.Z.; Chen, Y.L. The effect of phosphorus deficiency stress on activities of acid phosphatase in different clones of Chinese fir. Chin. J. Plant Ecol. 2005, 29, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.H. The Different Stages Response and its Signal Regulation of Different Chinese Fir with Different P-Use Efficiency to Low-Phosphorus Stress. Ph.D. Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2018. [Google Scholar]
- Tian, J.; Wang, X.R.; Tong, Y.P.; Chen, X.P.; Liao, H. Bioengineering and management for efficient phosphorus utilization in crops and pastures. Curr. Opin. Biotech. 2012, 23, 866–871. [Google Scholar] [CrossRef]
- Liu, T.; Chen, H.Y.; Yu, H.Y.; Li, T.X.; Gao, S.Q.; Chen, G.D. Characterization of phosphorus utilization in Barley leaf under low phosphorus stress. Chin. Bull. Bot. 2016, 51, 504–514. [Google Scholar] [CrossRef]
- Feike, A.D.; He, M.; Johansen, M.P.; Jennifer, J.H.; Claudia, K. Plant and microbial uptake of nitrogen and phosphorus affected by drought using 15N and 32P tracers. Soil Biol. Biochem. 2015, 82, 135–142. [Google Scholar] [CrossRef]
- Mildaryani, W.; Mujiyo, M.; Dewi, W.S.; Poernomo, D. Isotopic tracing of phosphorus uptake in Oil Palm seedlings leaf axil using 32P labelled. Int. J. Adv. Sci. Eng. Inf. Tech. 2020, 10, 368–373. [Google Scholar] [CrossRef]
- Sheng, W.T.; Fan, S.H. Soil changes of Chinese fir plantation. In Long-Term Productivity of Chinese Fir Plantations; China Science Publishing: Beijing, China, 2005; pp. 78–80. [Google Scholar]
- Wu, P.F.; Ma, X.Q.; Mulualem, T.; Wang, C.; Liu, A.Q.; Odén, P.C. Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can. J. For. Res. 2011, 41, 228–234. [Google Scholar] [CrossRef]
- Howard, G. The early history of 32P as a radioactive tracer in biochemical research. Biochem. Mol. Biol. Edu. 2005, 33, 159–164. [Google Scholar] [CrossRef]
- Kedrowski, R.A. Extraction and analysis of nitrogen, phosphorus and carbon fractions in plant material. J. Plant Nutr. 1983, 6, 989–1011. [Google Scholar] [CrossRef]
- Nurmayulis, N.; Citraresmini, A.; Anas, I. The use of 32P method to evaluate the growth of lowland rice cultivated in a system of rice intensification (SRI). At. Indones. 2013, 39, 88. [Google Scholar] [CrossRef]
- Yang, X.L.; Liu, Y.X.; Wu, F.K.; Jiang, X.J.; Yu, L.; Wang, Z.Q.; Zhang, Z.L.; Ma, J.; Chen, G.D.; Wei, Y.M.; et al. Quantitative trait loci analysis of root traits under phosphorus deficiency at the seedling stage in wheat. Genome 2018, 61, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Tsujii, Y.; Onoda, Y.; Kitayama, K. Phosphorus and nitrogen resorption from different chemical fractions in senescing leaves of tropical tree species on Mount Kinabalu, Borneo. Oecologia 2017, 185, 171–180. [Google Scholar] [CrossRef]
- Hidaka, A.; Kitayama, K. Relationship between photosynthetic phosphorus-use efficiency and foliar phosphorus fractions in tropical tree species. Ecol. Evol. 2013, 3, 4872–4880. [Google Scholar] [CrossRef]
- Mo, Q.F.; Li, Z.A.; Sayer, E.J.; Lambers, H.; Li, Y.; Zou, B.; Tang, J.W.; Heske, M.; Ding, Y.Z.; Wang, F.M. Foliar phosphorus fractions reveal how tropical plants maintain photosynthetic rates despite low soil phosphorus availability. Funct. Ecol. 2019, 33, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Perkins, M.C.; Woods, H.A.; Harrison, J.F.; Elser, J.J. Dietary phosphorus affects the growth of larval Manduca sexta. Arch. Insect Biochem. Physiol. 2004, 55, 153–168. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef]
- Valverde-Barrantes, O.J.; Gregoire, T.F.; Roumet, C.; Blackwood, C.B. A worldview of root traits: The influence of ancestry, growth form, climate and mycorrhizal association on the functional trait variation of fine-root tissues in seed plants. New Phytol. 2017, 215, 1295–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimeová, M.; Rindi, F.; Kaloud, P. There is more than meets the eye: DNA cloning demonstrates high genetic heterogeneity in populations of the subaerial green alga Trentepohlia (Trentepohliales, Chlorophyta). J. Phycol. 2018, 55, 224–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Cresswell, T.; Johansen, M.P.; Harrison, J.J.; Jiang, Y.; Keitel, C.; Cavagnaro, T.R.; Dijkstra, F.A. Re-allocation of nitrogen and phosphorus from roots drives regrowth of grasses and sedges after defoliation under deficit irrigation and nitrogen enrichment. J. Ecol. 2021, 109, 4071–4080. [Google Scholar] [CrossRef]
- Meyer, G.; Bell, M.J.; Doolette, C.L.; Brunetti, G.; Zhang, Y.Q.; Lombi, E.; Kopittke, P.M. Plant-Available Phosphorus in Highly Concentrated Fertilizer Bands: Effects of Soil Type, Phosphorus Form, and Coapplied Potassium. J. Agric. Food Chem. 2020, 29, 7571–7580. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, N.N.; Lin, J.Q.; Zhang, Y.; Ma, X.Q.; Wu, P.F. Root system-rhizosphere soil-bulk soil interactions in different Chinese fir clones based on fungi community diversity change. Front. Ecol. Evol. 2022, 10, 1028686. [Google Scholar] [CrossRef]
- Peng, Y.; Duan, Y.S.; Huo, W.G.; Zhang, Z.J.; Huang, D.; Xu, M.G.; Wang, X.H.; Yang, X.Y.; Wang, B.R.; Kuzyakov, Y.; et al. C:P stoichiometric imbalance between soil and microorganisms drives microbial phosphorus turnover in the rhizosphere. Biol. Fert. Soils 2022, 58, 421–433. [Google Scholar] [CrossRef]
- Muhammad, H.U.R.; Guo, H.L.; Zheng, S.S.; Li, L.X.; Ma, X.Q.; Taimoor, H.F.; Muhammad, F.N.; Narayan, P.G.; Wu, P.F. Effects of low phosphorus availability on root cambial activity, biomass production and root morphological pattern in two clones of Chinese fir. Forestry 2022, cpac030. [Google Scholar] [CrossRef]
- Zou, X.H.; Wu, P.F.; Chen, N.L.; Wang, P.; Ma, X.Q. Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil. Can. J. For. Res. 2015, 45, 402–410. [Google Scholar] [CrossRef]
- Zou, X.H.; Hu, Y.N.; Wei, D.; Chen, S.T.; Wu, P.F.; Ma, X.Q. Correlation between endogenous hormone and the adaptability of Chinese fir with high phosphorus-use efficiency to low phosphorus stress. Chin. J. Plant Ecol. 2019, 43, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.F.; Wang, G.Y.; El-Kassaby, Y.A.; Wang, P.; Zou, X.H.; Ma, X.Q. Solubilization of aluminum-bound phosphorus by root cell walls: Evidence from Chinese fir, Cunninghamia lanceolata. Can. J. For. Res. 2017, 47, 419–423. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P Fractions | PUE for Dry Mass (g μg−1) | |||||
|---|---|---|---|---|---|---|
| Leaf | Stem | Root | ||||
| L-P | H-P | L-P | H-P | L-P | H-P | |
| Inorganic P | −0.990 ** | −0.734 ns | −0.798 * | −0.701 ns | −0.018 ns | 0.661 ns |
| Ester P | −0.980 ** | −0.858 * | −0.816 * | −0.607 ns | 0.056 ns | −0.796 * |
| Lipid P | −0.720 ns | −0.772 ns | −0.642 ns | −0.723 ns | 0.440 ns | −0.623 ns |
| Nucleic P | −0.940 ** | −0.767 ns | −0.813 * | −0.785 ns | −0.878 ** | −0.549 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Liu, Q.; Huang, Z.; Chen, S.; Wu, P.; Ma, X.; Cai, L. Allocation of Phosphorus Fractions in Chinese Fir in Response to Low Phosphorus Availability Using 32P Tracer. Forests 2022, 13, 1769. https://doi.org/10.3390/f13111769
Zou X, Liu Q, Huang Z, Chen S, Wu P, Ma X, Cai L. Allocation of Phosphorus Fractions in Chinese Fir in Response to Low Phosphorus Availability Using 32P Tracer. Forests. 2022; 13(11):1769. https://doi.org/10.3390/f13111769
Chicago/Turabian StyleZou, Xianhua, Qingqing Liu, Zhijun Huang, Sitong Chen, Pengfei Wu, Xiangqing Ma, and Liping Cai. 2022. "Allocation of Phosphorus Fractions in Chinese Fir in Response to Low Phosphorus Availability Using 32P Tracer" Forests 13, no. 11: 1769. https://doi.org/10.3390/f13111769
APA StyleZou, X., Liu, Q., Huang, Z., Chen, S., Wu, P., Ma, X., & Cai, L. (2022). Allocation of Phosphorus Fractions in Chinese Fir in Response to Low Phosphorus Availability Using 32P Tracer. Forests, 13(11), 1769. https://doi.org/10.3390/f13111769