
Effects of Forest Gap on Soil Microbial Communities in an Evergreen Broad-Leaved Secondary Forest

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
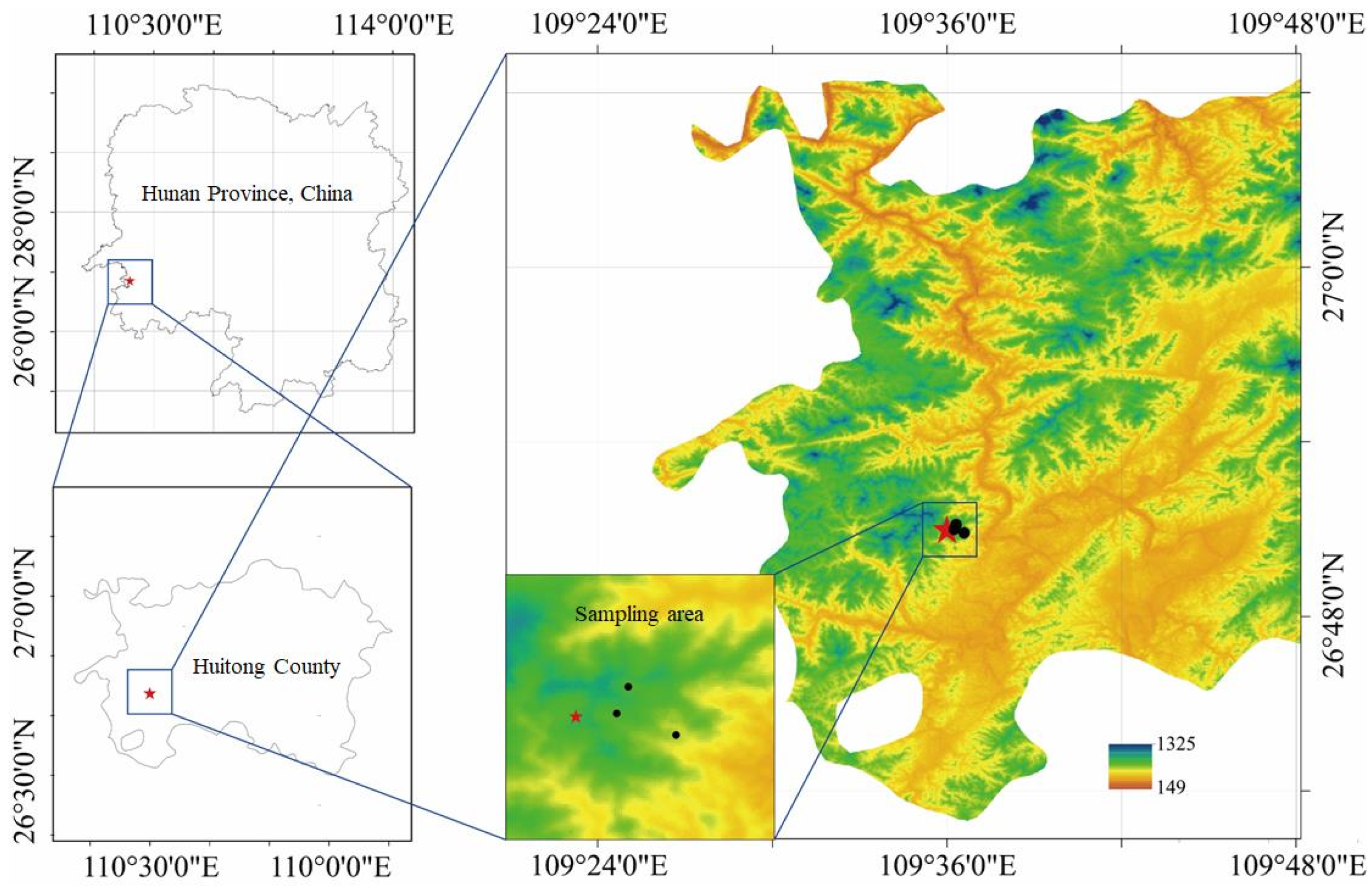
2.1. Site Description
2.2. Experimental Design
2.3. Analysis Methods
2.3.1. Basic Chemical Properties of Soil
2.3.2. Soil Microbial Community Structure
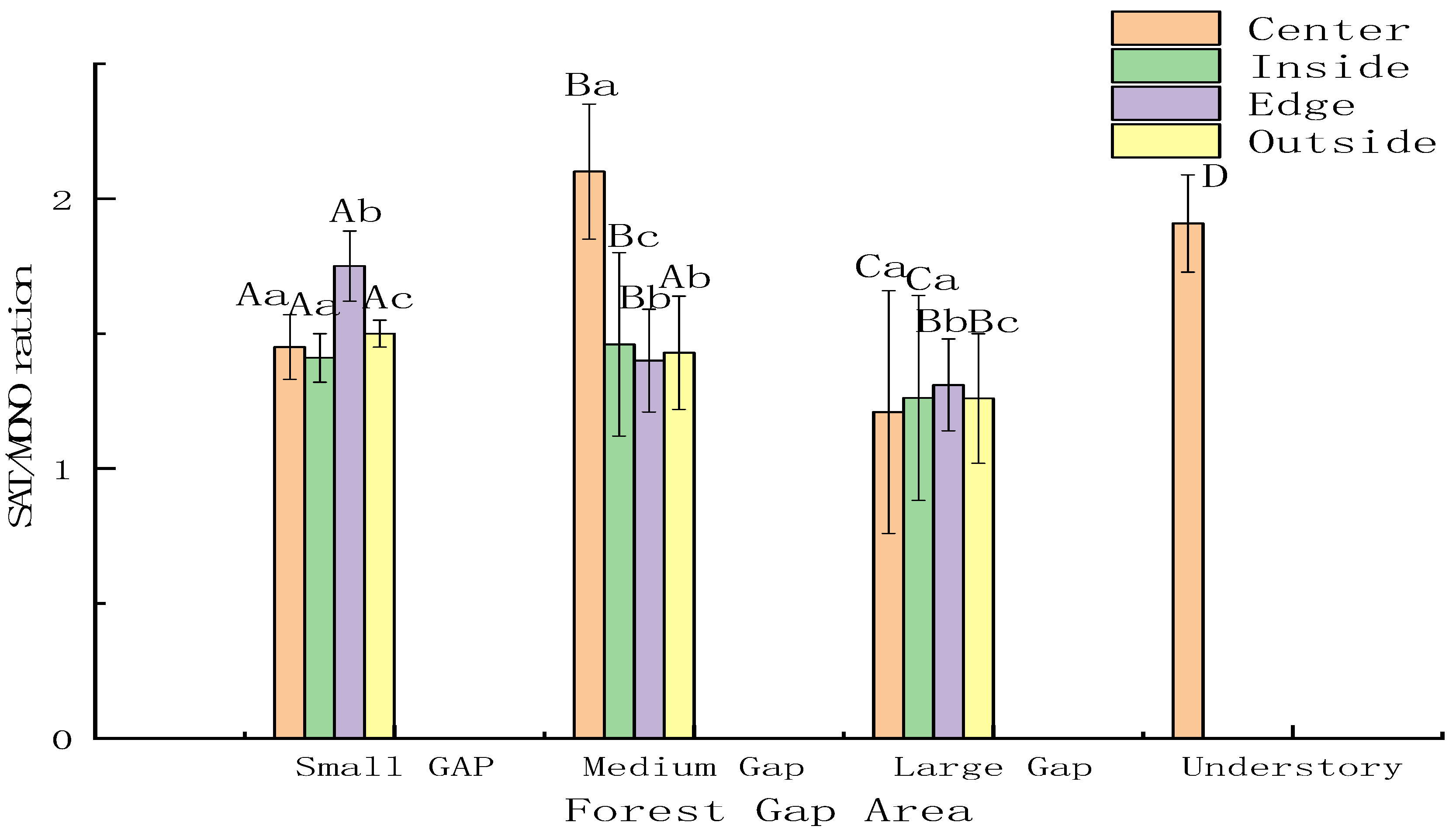
2.3.3. Saturated Fatty Acids/Monounsaturated Characteristics
2.4. Statistical Analysis
3. Results
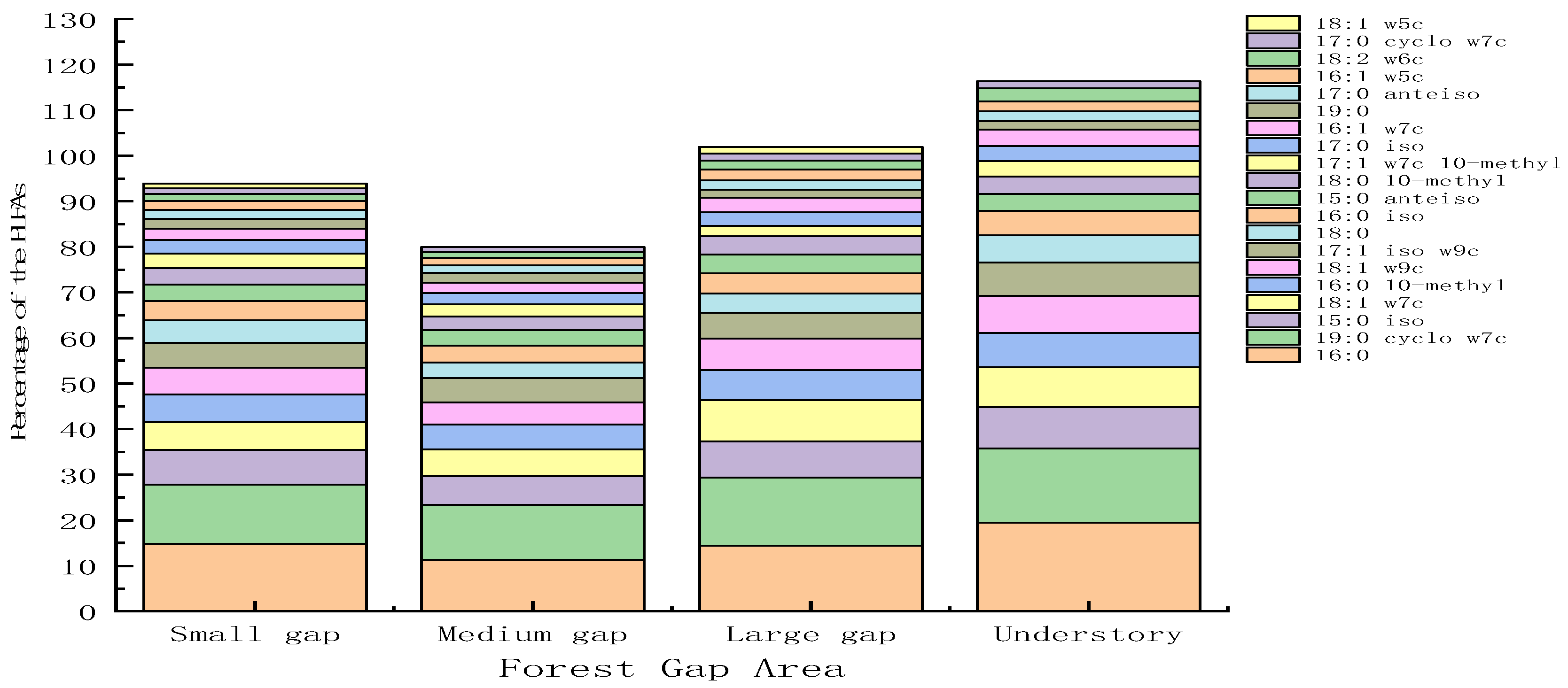
3.1. Response of Soil Microbial Community Structure to Forest Gap
3.2. Response of Soil Microbial Community Composition to Forest Gap
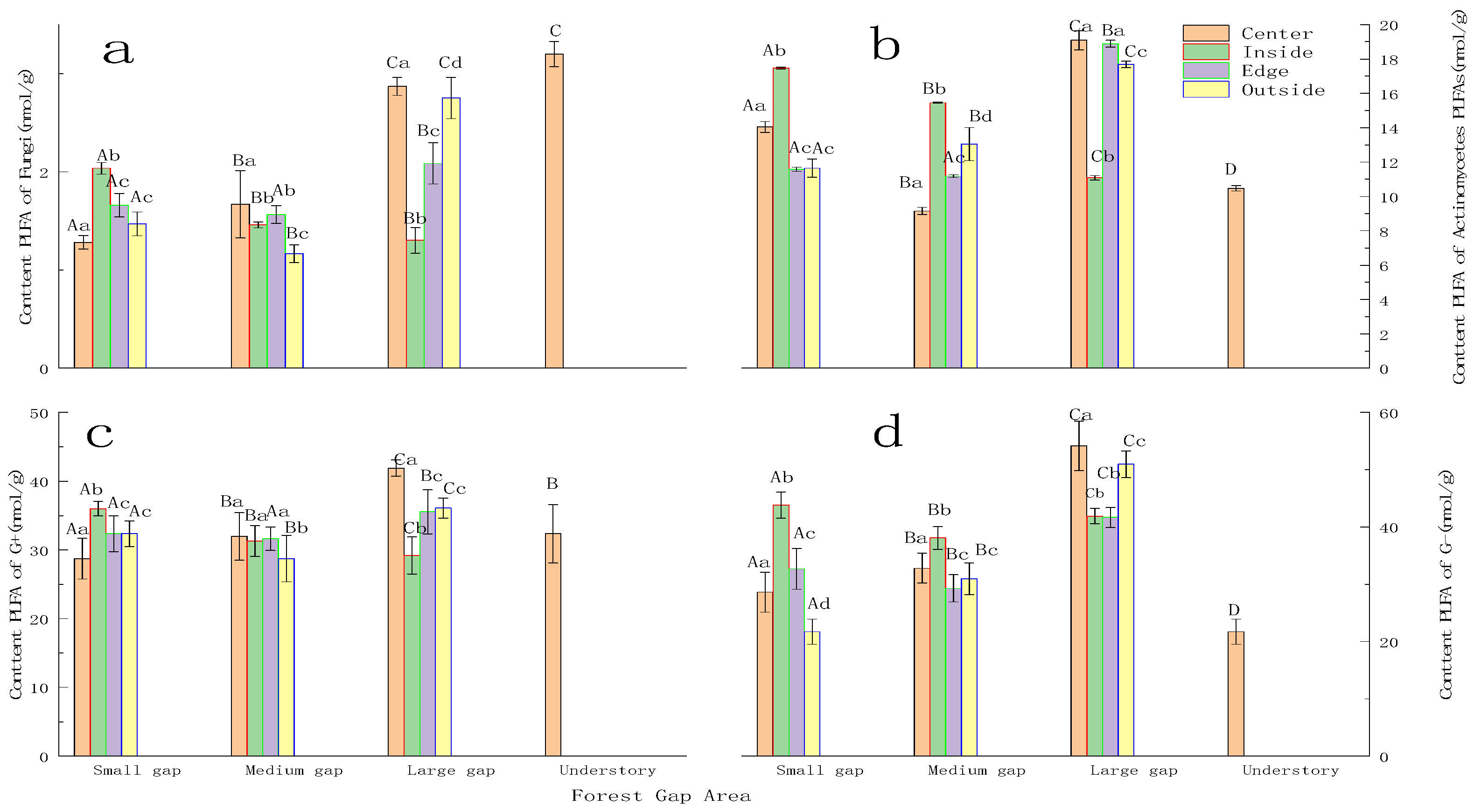
3.2.1. Soil Fungi, Actinomycetes, G+ Bacteria, and G− Bacteria Response to Gap Size
3.2.2. Soil AM Fungi and Anaerobic Response to Forest Gap
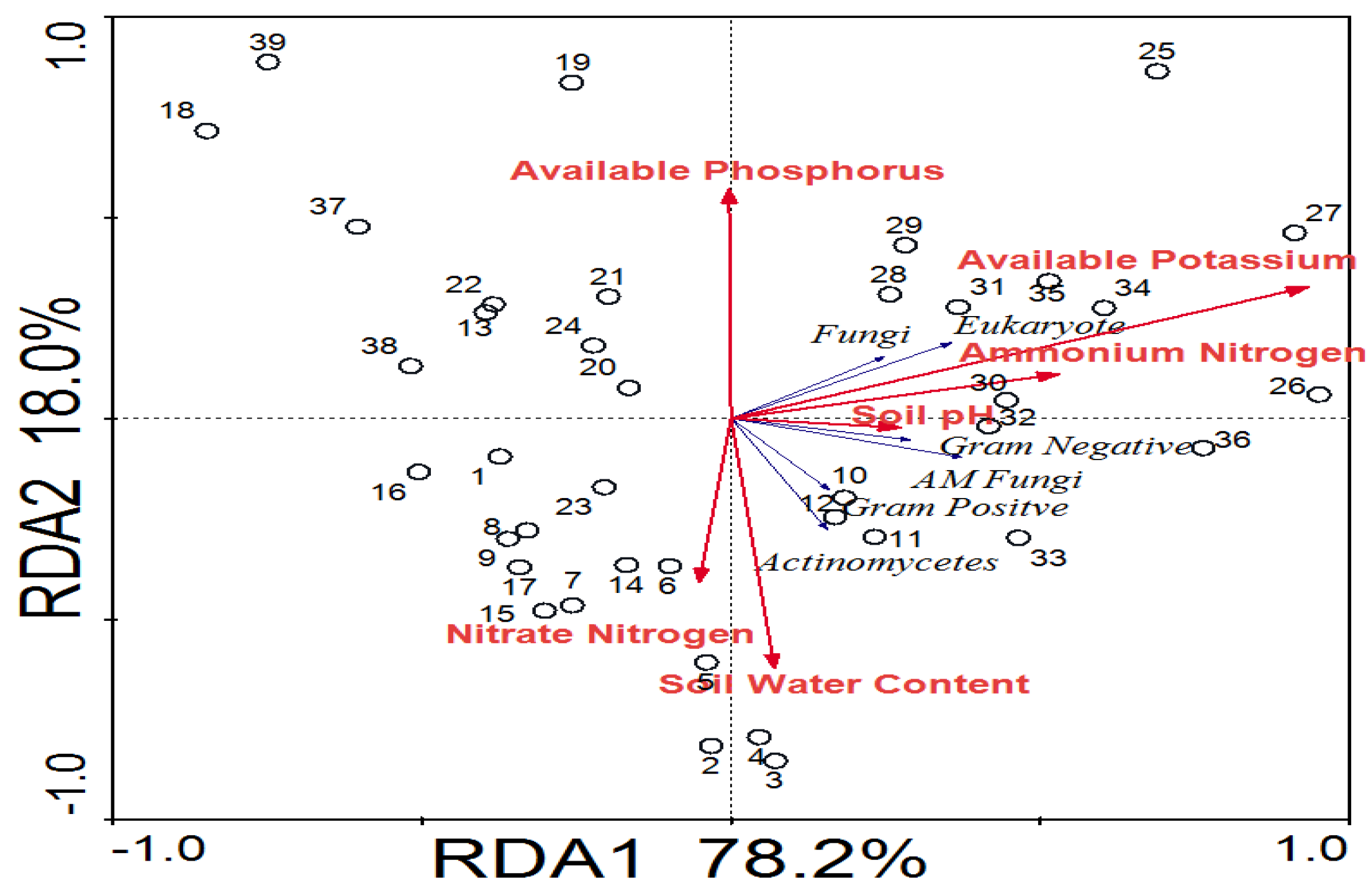
3.3. Soil Factors Driving Soil Microbial Community Shift
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mantovani, M.T.W. Gap-phase regeneration in a tropical montane forest the effects of gap structure and bamboo species. Plant Ecol. 2000, 148, 149–155. [Google Scholar]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Thompson, J.; Proctor, J.; Scott, D.A.; Fraser, P.J.; Marrs, R.H.; Miller, R.P.; Viana, V. Rain forest on Maracá Island Roraima Brazil artificial gaps and plant response to them. For. Ecol. Manag. 1998, 102, 305–321. [Google Scholar] [CrossRef]
- Orman, O.; Dobrowolska, D. Gap dynamics in the Western Carpathian mixed beech old-growth forests affected by spruce bark beetle outbreak. Eur. J. For. Res. 2017, 136, 571–581. [Google Scholar] [CrossRef]
- Forbes, A.S.; Norton, D.A.; Carswell, F.E. Artificial canopy gaps accelerate restoration within an exotic Pinus radiata plantation. Restor. Ecol. 2016, 24, 336–345. [Google Scholar] [CrossRef]
- Zhu, J.; Matsuzaki, T.; Lee, F.-Q.; Gonda, Y. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. For. Ecol. Manag. 2003, 182, 339–354. [Google Scholar] [CrossRef]
- Keram, A.; Halik, Ü.; Keyimu, M.; Aishan, T.; Mamat, Z.; Rouzi, A. Gap dynamics of natural Populus euphratica floodplain forests affected by hydrological alteration along the Tarim River: Implications for restoration of the riparian forests. For. Ecol. Manag. 2019, 438, 103–113. [Google Scholar] [CrossRef]
- Gálhidy, L.; Mihók, B.; Hagyó, A.; Rajkai, K.; Standovár, T. Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest. Plant Ecol. 2005, 183, 133–145. [Google Scholar] [CrossRef]
- Zhao, Q.; Pang, X.; Bao, W.; He, Q. Effects of gap-model thinning intensity on the radial growth of gap-edge trees with distinct crown classes in a spruce plantation. Trees 2015, 29, 1861–1870. [Google Scholar] [CrossRef]
- Schliemann, S.A.B.; James, G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Vajari, K.A. The influece of forest gaps on some properties of humus in a managed beech forest, northern Iran. Eurasian Soil Sci. 2015, 48, 1131–1135. [Google Scholar] [CrossRef]
- Ritter, E.; Dalsgaard, L.; Einhorn, K.S. Light, temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark. For. Ecol. Manag. 2005, 206, 15–33. [Google Scholar] [CrossRef]
- Barreiro, A.; Martín, A.; Carballas, T.; Díaz-Raviña, M. Response of soil microbial communities to fire and fire-fighting chemicals. Sci. Total Environ. 2010, 408, 6172–6178. [Google Scholar] [CrossRef] [PubMed]
- Baena, C.W.; Andrés-Abellán, M.; Lucas-Borja, M.E.; Martínez-García, E.; García-Morote, F.A.; Rubio, E.; López-Serrano, F.R. Thinning and recovery effects on soil properties in two sites of a Mediterranean forest, in Cuenca Mountain (South-eastern of Spain). For. Ecol. Manag. 2013, 308, 223–230. [Google Scholar] [CrossRef]
- Hu, B.; Yang, B.; Pang, X.; Bao, W.; Tian, G. Responses of soil phosphorus fractions to gap size in a reforested spruce forest. Geoderma 2016, 279, 61–69. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests. Can. J. For. Res. 2002, 32, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Ni, X.; Yang, W.; Tan, B.; Li, H.; He, J.; Xu, L.; Wu, F. Forest gaps slow the sequestration of soil organic matter: A humification experiment with six foliar litters in an alpine forest. Sci. Rep. 2016, 6, 19744. [Google Scholar] [CrossRef] [Green Version]
- Bauhus, J.; Bartsch, N. Mechanisms for carbon and nutrient release and retention in beech forest gaps. Plant Soil 1995, 168, 579–584. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, X. Soil properties in forest gaps and under canopy in broad-leaved Pinus koraiensis forests in Changbai Mountainous Region, China. Front. For. China 2007, 2, 60–65. [Google Scholar] [CrossRef]
- Achat, D.L.; Augusto, L.; Bakker, M.R.; Gallet-Budynek, A.; Morel, C. Microbial processes controlling P availability in forest spodosols as affected by soil depth and soil properties. Soil Biol. Biochem. 2012, 44, 39–48. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, F.; Tan, B.; Yang, W.; Ni, X.; Yang, Y. Carbon, nitrogen and phosphorus stocks in soil organic layer as affected by forest gaps in the alpine forest of the eastern Tibet Plateau. Russ. J. Ecol. 2015, 46, 246–251. [Google Scholar] [CrossRef]
- Han, M.; Tang, M.; Shi, B.; Jin, G. Effect of canopy gap size on soil respiration in a mixed broadleaved-Korean pine forest: Evidence from biotic and abiotic factors. Eur. J. Soil Biol. 2020, 99, 103194. [Google Scholar] [CrossRef]
- Bååth, E.; Díaz-Raviña, M.; Bakken, L.R. Microbial Biomass, Community Structure and Metal Tolerance of a Naturally Pb-Enriched Forest Soil. Microb. Ecol. 2005, 50, 496–505. [Google Scholar] [CrossRef]
- Banning, N.C.; Murphy, D.V. Effect of heat-induced disturbance on microbial biomass and activity in forest soil and the relationship between disturbance effects and microbial community structure. Appl. Soil Ecol. 2008, 40, 109–119. [Google Scholar] [CrossRef]
- Bhardwaj, Y.; Sharma, M.P.; Pandey, J.; Dubey, S.K. Variations in microbial community in a tropical dry deciduous forest across the season and topographical gradient assessed through signature fatty acid biomarkers. Ecol. Res. 2020, 35, 139–153. [Google Scholar] [CrossRef]
- Berg, E.C.; Zarnoch, S.J.; McNab, W.H. Twenty-year survivorship of tree seedlings in wind-created gaps in an upland hardwood forest in the eastern US. New For. 2019, 50, 323–344. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, W.; Zhang, J.; Xu, Z.; Liu, Y.; Li, H.; You, C.; Tan, B. Forest Gap Size Alters the Functional Diversity of Soil Nematode Communities in Alpine Forest Ecosystems. Forests 2019, 10, 806. [Google Scholar] [CrossRef] [Green Version]
- Mogilewsky, M.; Vasey, N.; Andriamahaihavana, M.A.; Rakotomalala, Z. Contribution of tree-fall canopy gaps to variation in nitrogen content of fruits and leaves from Varecia and Eulemur food trees in northeastern Madagascar. Am. J. Phys. Anthropol. 2019, 168, 167–168. [Google Scholar]
- Dietz, L.; Collet, C.; Dupouey, J.; Lacombe, E.; Laurent, L.; Gégout, J. Windstorm-induced canopy openings accelerate temperate forest adaptation to global warming. Glob. Ecol. Biogeogr. 2020, 29, 2067–2077. [Google Scholar] [CrossRef]
- Herault, B.; Ouallet, J.; Blanc, L.; Wagner, F.; Baraloto, C. Growth responses of neotropical trees to logging gaps. J. Appl. Ecol. 2010, 47, 821–831. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Bockheim, J.G. Impacts of forest gaps on soil properties and processes in old growth northern hardwood-hemlock forests. Plant Soil 2007, 294, 219–233. [Google Scholar] [CrossRef]
- Kushwaha, P.; Neilson, J.W.; Barberán, A.; Chen, Y.; Fontana, C.G.; Butterfield, B.J.; Maier, R.M. Arid Ecosystem Vegetation Canopy-Gap Dichotomy: Influence on Soil Microbial Composition and Nutrient Cycling Functional Potential. Appl. Environ. Microbiol. 2021, 87, e02780-20. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps in the forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Yang, B.; Pang, X.; Hu, B.; Bao, W.; Tian, G. Does thinning-induced gap size result in altered soil microbial community in pine plantation in eastern Tibetan Plateau? Ecol. Evol. 2017, 7, 2986–2993. [Google Scholar] [CrossRef]
- Han, W.; Wang, G.; Liu, J.; Ni, J. Effects of vegetation type, season, and soil properties on soil microbial community in subtropical forests. Appl. Soil Ecol. 2021, 158, 103813. [Google Scholar] [CrossRef]
- Li, D.; Li, X.; Su, Y.; Li, X.; Yin, H.; Li, X.; Guo, M.; He, Y. Forest gaps influence fungal community assembly in a weeping cypress forest. Appl. Microbiol. Biotechnol. 2019, 103, 3215–3224. [Google Scholar] [CrossRef] [PubMed]
- Kisanuki, H.; Nakasu, M.; Nakai, A.; Yurugi, Y. Predicting the population dynamics of three understory broad-leaved evergreen species under the influence of Sika deer in primary and secondary forests of mid-western Japan. J. For. Res. 2008, 13, 52–58. [Google Scholar] [CrossRef]
- Li, X.; Wilson, S.; Song, Y. Secondary succession in two subtropical forests. Plant Ecol. 1999, 143, 13–21. [Google Scholar] [CrossRef]
- Pang, X.; Bao, W.; Zhu, B.; Cheng, W. Responses of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agric. For. Meteorol. 2013, 171–172, 57–64. [Google Scholar] [CrossRef]
- Bolat, I. The effect of thinning on microbial biomass C, N and basal respiration in black pine forest soils in Mudurnu, Turkey. Eur. J. For. Res. 2013, 133, 131–139. [Google Scholar] [CrossRef]
- Xiao, F. Mensuration of respiration amount in the community of secondary evergreen broadleaved forests in Huitong County, Hu’nan Province. J. Beijing For. Univ. 2006, 28, 40–44. [Google Scholar]
- Yan, S.; Wang, S.; Hu, Y.; Gao, H.; Zhang, X. A comparative study on soil fauna in native secondary evergreen broad-leaved forest and Chinese fir plantation forests in subtropics. Chin. J. Appl. Ecol. 2004, 15, 1792–1796. [Google Scholar]
- Wang, J.; Zhang, Y.; Ma, Y.; Liu, Y.; Li, Y.; Duan, W. Linear character of the sunshine of the gap in the artifical forest in Xishuangbanna. J. Plant Resour. Environ. 2000, 9, 27–30. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Runkle, J.R. Patterns of Disturbance in Some Old-Growth Mesic Forests of Eastern North America. Ecology 1982, 63, 1533–1546. [Google Scholar] [CrossRef] [Green Version]
- Amir, A.A.D.; Norman, C. Distinct characteristics of canopy gaps in the subtropical mangroves of Moreton Bay, Australia. Estuar. Coast. Shelf Sci. 2019, 222, 66–80. [Google Scholar] [CrossRef]
- Jiménez, J.J.; Igual, J.M.; Villar, L.; Benito-Alonso, J.L.; Abadias-Ullod, J. Hierarchical drivers of soil microbial community structure variability in “Monte Perdido” Massif (Central Pyrenees). Sci. Rep. 2019, 9, 8768. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Luan, Y.; Wang, B.; Dai, W.; Zhao, M. Analysis of spatiotemporal variations in the characteristics of soil microbial communities in Castanopsis fargesii forests. J. For. Res. 2019, 31, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Pollierer, M.M.; Ferlian, O.; Scheu, S. Temporal dynamics and variation with forest type of phospholipid fatty acids in litter and soil of temperate forests across regions. Soil Biol. Biochem. 2015, 91, 248–257. [Google Scholar] [CrossRef]
- Ou, J.; Liu, Y.; Zhang, J.; Zhang, J.; Cui, N.; Deng, C.; Ji, T.; He, R. Early influence of forest gap harvesting on soil phosphorus in Pinus massoniana plantation in a hilly area of the reaches of Yangtze River. Chin. J. Ecol. 2014, 33, 592–601. [Google Scholar]
- Wei, Q. Infuence of forest gaps on the available nitrogen of soil in a pinus massoniaan plantation. J. For. Environ. 2021, 41, 124–131. [Google Scholar]
- Zhu, J.-J.; Tan, H.; Li, F.-Q.; Chen, M.; Zhang, J.-X. Microclimate regimes following gap formation in a montane secondary forest of eastern Liaoning Province, China. J. For. Res. 2007, 18, 167–173. [Google Scholar] [CrossRef]
- Wang, Q.; Pieristè, M.; Liu, C.; Kenta, T.; Robson, T.M.; Kurokawa, H. The contribution of photodegradation to litter decomposition in a temperate forest gap and understorey. New Phytol. 2021, 229, 2625–2636. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-G.; Hao, K.-J.; Li, X.-W.; Fan, C.; Chen, Y.-L.; Liu, Y.-K.; Wang, X. Effects of forest gap on seasonal dynamics of soil organic carbon and microbial biomass car- bon in Picea asperata forest in Miyaluo of Western Sichuan, Southwest China. Chin. J. Appl. Ecol. 2014, 25, 2469–2476. [Google Scholar]
- Wang, X.; Liu, J.; He, Z.; Xing, C.; Zhu, D. Forest gaps mediate the structure and function of the soil microbial community in a castanopsis kawakamii forest. Ecol. Indic. 2021, 122, 107288. [Google Scholar] [CrossRef]
- Chen, L.; Han, W.; Liu, D.; Liu, G. How forest gaps shaped plant diversity along an elevational gradient in Wolong National Nature Reserve? J. Geogr. Sci. 2019, 29, 1081–1097. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.Q.; Han, P.-B.; Li, S.; Shen, L.-Q.; Huang, D.-S.; Li, Y.-F.; Peng, M.-C.; Wang, C.-Y.; Li, X.-S.; Li, W.; et al. Species richness, forest types and regeneration of Schima in the subtropical forest ecosystem of Yunnan, southwestern China. For. Ecosyst. 2020, 7, 35. [Google Scholar] [CrossRef]
- Modrow, T.; Kuehne, C.; Saha, S.; Bauhus, J.; Pyttel, P.L. Photosynthetic performance, height growth, and dominance of naturally regenerated sessile oak (Quercus petraea Mattuschka Liebl.) seedlings in small-scale canopy openings of varying sizes. Eur. J. For. Res. 2020, 139, 41–52. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
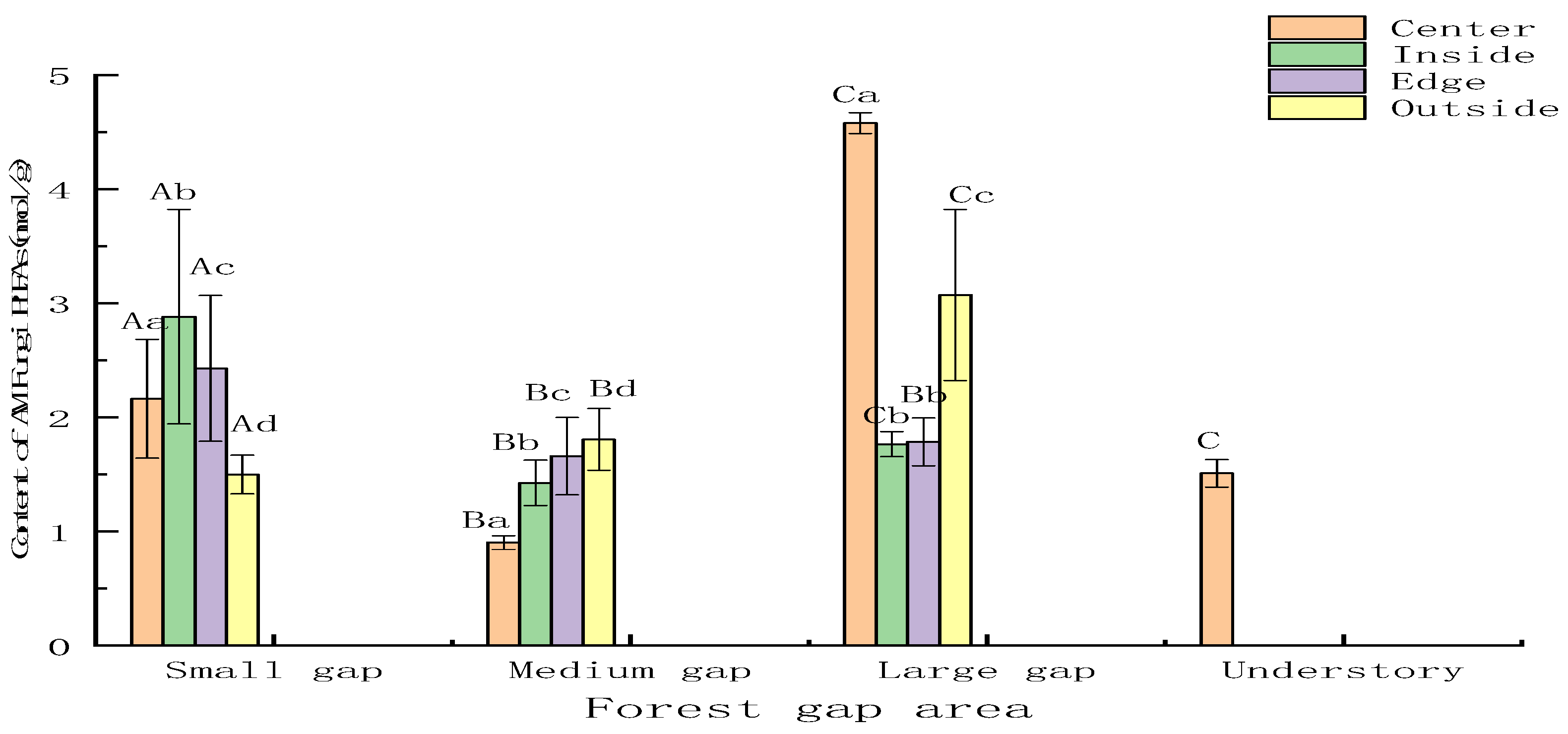{kind=link}
{kind=link}
| Forest Gap | D/H | Area/m2 | Slope Direction | Geographic Position | Formation |
|---|---|---|---|---|---|
| G1 | 0.3 | 60 | East | 109°36′6″ E 26°51′11″ N | Cutting |
| G2 | 1.3 | 130 | Northeast | 109°36′8″ E 26°51′13″ N | Cutting |
| G3 | 2.3 | 270 | East | 109°36′7″ E 26°51′09″ N | Cutting |
| G4 | 0.3 | 73 | Southeast | 109°36′9″ E 26°51′12″ N | Cutting |
| G5 | 1.3 | 149 | Southeast | 109°36′9″ E 26°51′07″ N | Cutting |
| G6 | 2.3 | 286 | Northeast | 109°36′10″ E 26°47′10″ N | Cutting |
| G7 | 0.3 | 80 | East | 109°36′10″ E 26°50′15″ N | Cutting |
| G8 | 1.3 | 160 | Northeast | 109°36′11″ E 26°52′14″ N | Cutting |
| G9 | 2.3 | 300 | East | 109°36′15″ E 26°51′14″ N | Cutting |
| Understory | — | 100 | Southeast | 109°36′16″ E 26°49′14″ N | Natural |
| Understory | — | 100 | Northeast | 109°36′18″ E 26°46′14″ N | Natural |
| Understory | — | 100 | East | 109°36′17″ E 26°50′14″ N | Natural |
| Microbial Group | Phospholipids Fatty Acids Signatures |
|---|---|
| Actinomycetes | 16:0 10-methyl; 17:0 10-methyl; 17:1 w7c 10-methyl; 18:0 10-methyl; 18:1 w7c 10-methyl etc. |
| G+ bacteria | 11:0 anteiso; 11:0 iso; 12:0 anteiso; 12:0 iso; 13:0 anteiso; 13:0 iso; 14:0 anteiso; 14:0 iso; 14:1 iso w7c; 15:0 anteiso; 15:0 iso; 15:1 anteiso w9c; 15:1 iso w6c; 15:1 iso w9c; 16:0 anteiso; 16:0 iso; 17:0 anteiso; 17:0 iso; 17:1 iso w9c; 18:0 iso; 19:0 anteiso; 19:0 iso; 20:0 iso; 22:0 iso |
| G− bacteria | 13:1 w5c; 14:0 2OH; 14:1 w8c; 14:1 w9c; 15:1 w7c; 15:1 w8c; 5:1 w9c; 16:0 2OH; 17:0 cyclo w7c; 17:1 w3c; 21:1 w3c; 21:1 w4c; 21:1 w5c; 21:1 w6c; 21:1 w8c; 22:1 w6c; 22:1 w8c; 22:1 w9c; 24:1 w7c |
| Eukaryote | 15:3 w3c; 15:4 w3c; 16:3 w6c; 16:4 w3c; 18:3 w6c; 19:3 w3c; 19:3 w6c; 19:4 w6c; 20:2 w6c; 20:3 w6c; 20:4 w6c; 20:5 w3c; 21:3 w3c; 21:3 w6c; 22:2 w6c; 22:4 w6c 22:5 w3c; 22:5 w6c; 22:6 w3c; 23:1 w4c; 23:1 w5c; 23:3 w3c; 23:3 w6c; 23:4 w6c; 24:1 w3c; 24:3 w3c; 24:3 w6c; 24:4 w6c |
| AM Fungi | 18:2 w6c |
| Fungi | 16:1 w5c |
| Gap Type | Gap Position | ||||
|---|---|---|---|---|---|
| Center | Inside | Edge | Outside | Total | |
| Small gap | 100.82 ± 2.21 Aa | 95.71.41 ± 1.19 Bb | 94.25 ± 2.4 Cb | 90.08 ± 2.58 Dd | 94.35 ± 3.12 a |
| Medium gap | 108.93 ± 3.81 Ba | 105.00 ± 3.34 Bb | 98.48 ± 4.45 Bc | 95.00 ± 3.81 Dc | 101.73 ± 1.10 b |
| Large gap | 86.75 ± 2.63 Ca | 95.27 ± 1.63 Bb | 116.73 ± 2.83 Cc | 86.08 ± 1.41 Ba | 95.75 ± 2.06 c |
| Understory | 90.43 ± 3.62 C | 90.43 ± 3.62 a | |||
| Gap Size | Available Phosphorus | Available Potassium | Nitrate Nitrogen | Ammonium Nitrogen | Soil Water Content | Soil pH |
|---|---|---|---|---|---|---|
| Small gap | 1.83 ± 0.07 A | 44.10 ± 5.23 A | 2.1 ± 1.09 A | 8.57 ± 1.18 A | 0.38 ± 0.03 A | 4.28 ± 0.02 A |
| Medium gap | 2.09 ± 0.09 B | 43.88 ± 3.91 A | 2.48 ± 0.71 B | 8.93 ± 0.88 B | 0.37 ± 0.02 A | 4.37 ± 0.11 A |
| Large gap | 2.07 ± 0.09 C | 69.50 ± 7.09 B | 1.58 ± 0.62 C | 9.85 ± 4.43 C | 0.34 ± 0.01 B | 4.44 ± 0.08 A |
| Understory | 1.98 ± 0.06 A | 36.59 ± 0.3.7 C | 0.37 ± 0.01 D | 8.27 ± 0.04 A | 0.32 ± 0.05 C | 4.31 ± 0.04 A |
| Factor | df | Available Phosphorus | Available Potassium | Nitrate Nitrogen | Ammonium Nitrogen | Soil Water Content | Soil pH |
|---|---|---|---|---|---|---|---|
| Gap area | 3 | <0.01 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Gap position | 3 | <0.05 | <0.05 | <0.01 | <0.01 | <0.01 | 0.679 |
| Gap area × gap position | 9 | <0.05 | 0.269 | <0.01 | <0.01 | <0.05 | 0.963 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Jiang, C.; Bai, Y.; Wang, H.; Jiang, C.; Huang, K.; Guo, L.; Zeng, S.; Wang, S. Effects of Forest Gap on Soil Microbial Communities in an Evergreen Broad-Leaved Secondary Forest. Forests 2022, 13, 2015. https://doi.org/10.3390/f13122015
Chen S, Jiang C, Bai Y, Wang H, Jiang C, Huang K, Guo L, Zeng S, Wang S. Effects of Forest Gap on Soil Microbial Communities in an Evergreen Broad-Leaved Secondary Forest. Forests. 2022; 13(12):2015. https://doi.org/10.3390/f13122015
Chicago/Turabian StyleChen, Shiyou, Chunqian Jiang, Yanfeng Bai, Hui Wang, Chunwu Jiang, Ke Huang, Lina Guo, Suping Zeng, and Shuren Wang. 2022. "Effects of Forest Gap on Soil Microbial Communities in an Evergreen Broad-Leaved Secondary Forest" Forests 13, no. 12: 2015. https://doi.org/10.3390/f13122015
APA StyleChen, S., Jiang, C., Bai, Y., Wang, H., Jiang, C., Huang, K., Guo, L., Zeng, S., & Wang, S. (2022). Effects of Forest Gap on Soil Microbial Communities in an Evergreen Broad-Leaved Secondary Forest. Forests, 13(12), 2015. https://doi.org/10.3390/f13122015