

Alpine Shrubification: Juniper Encroachment into Tundra in the Ural Mountains

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
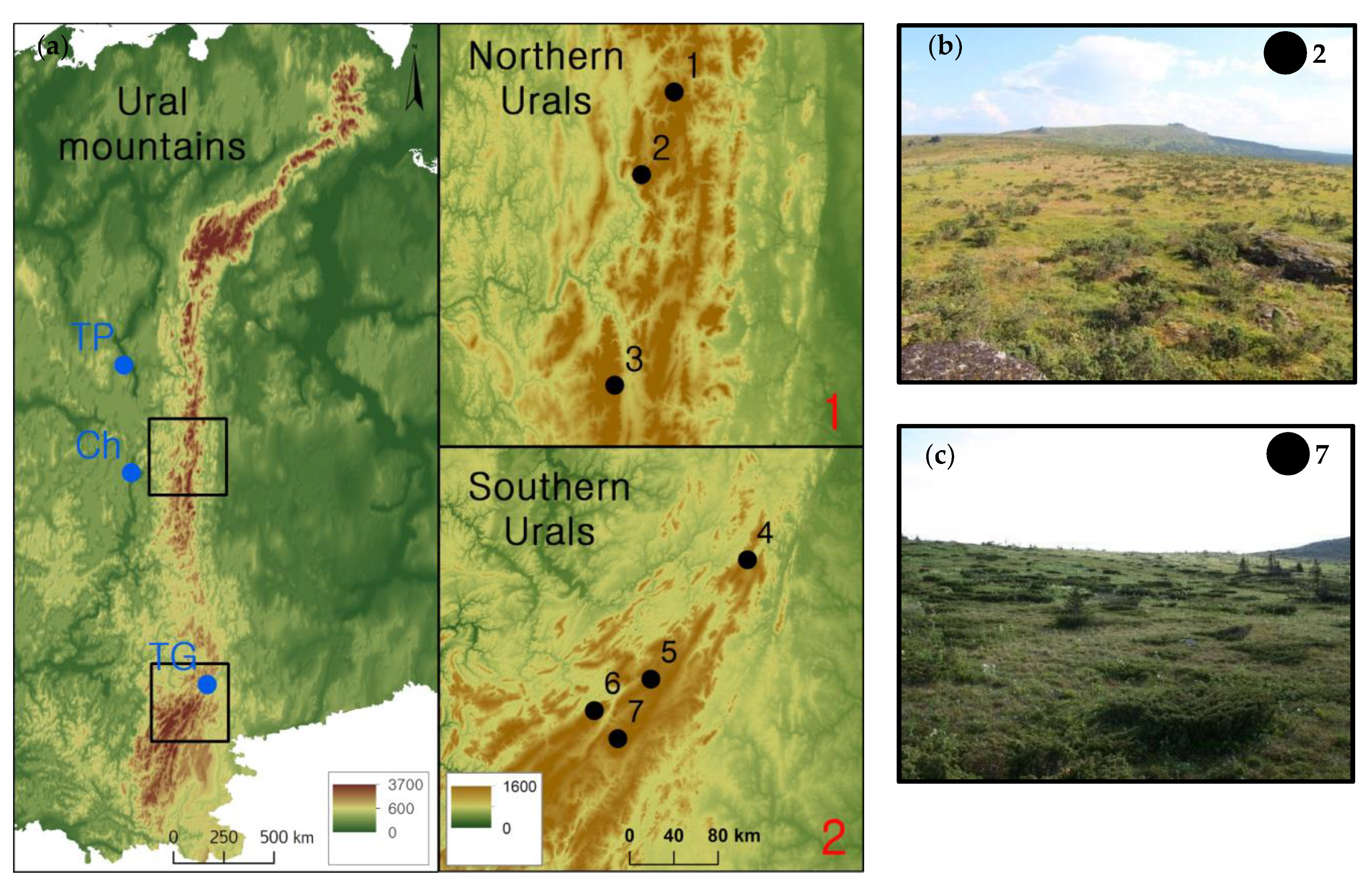
2.1. Study Sites
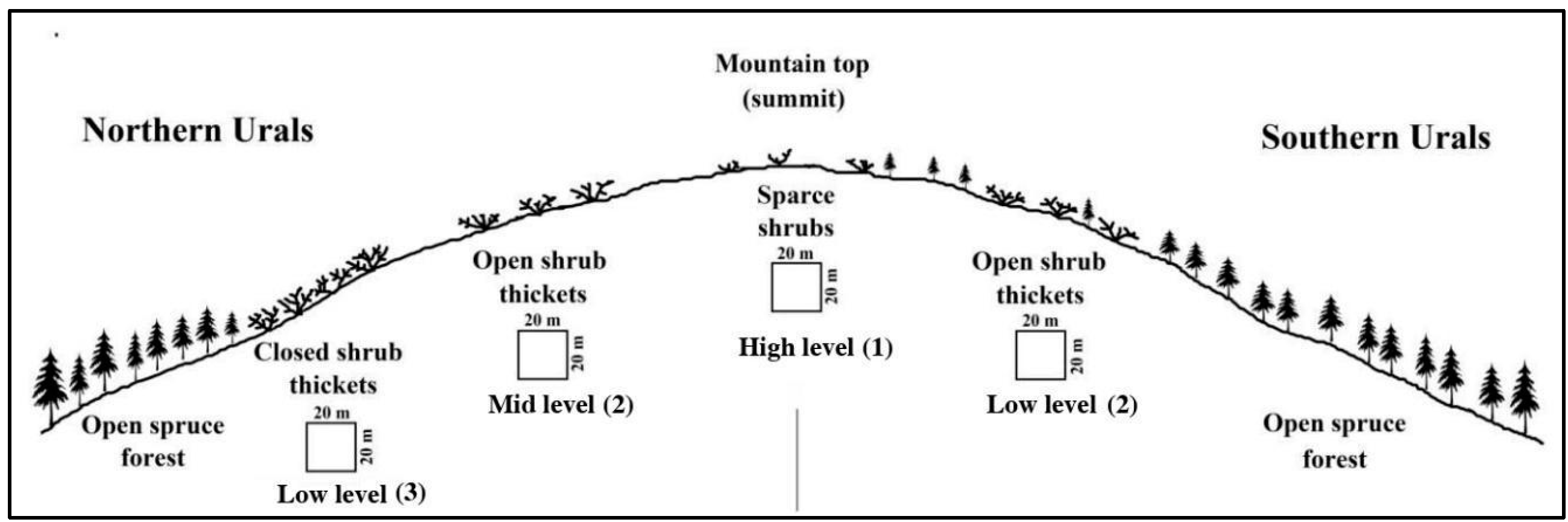
2.2. Design of Field Sampling
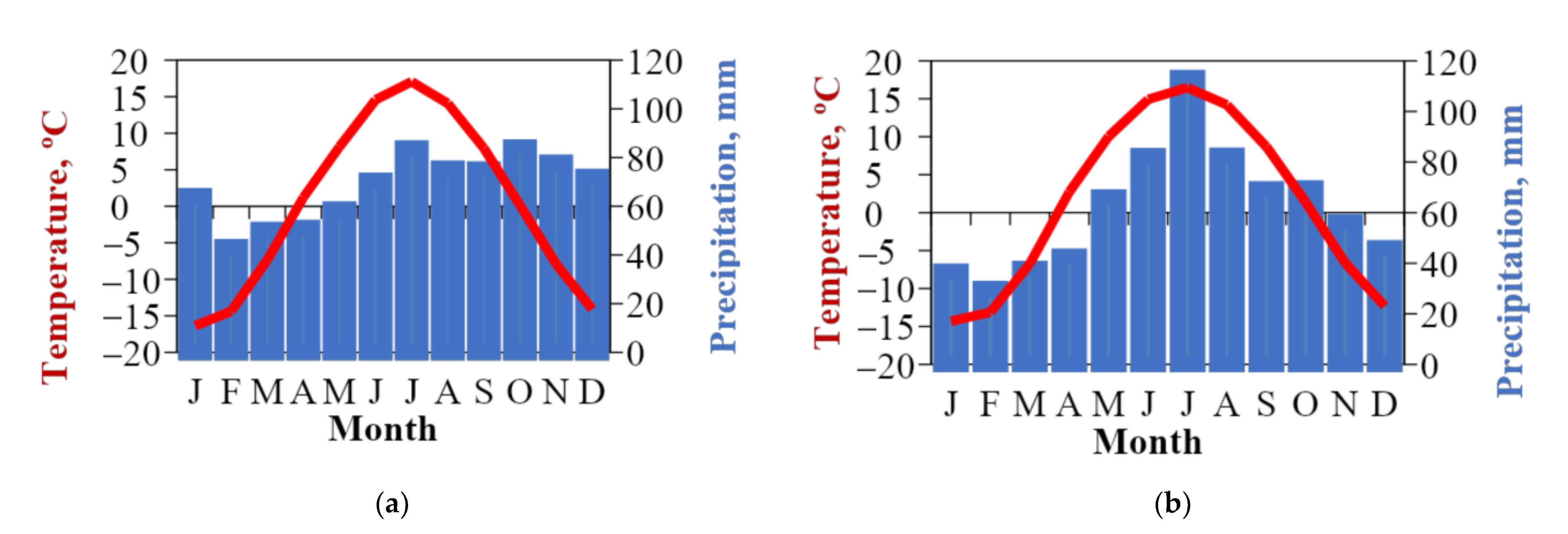
2.3. Climate Data
2.4. Associations between Climate and Juniper Recruitment
3. Results
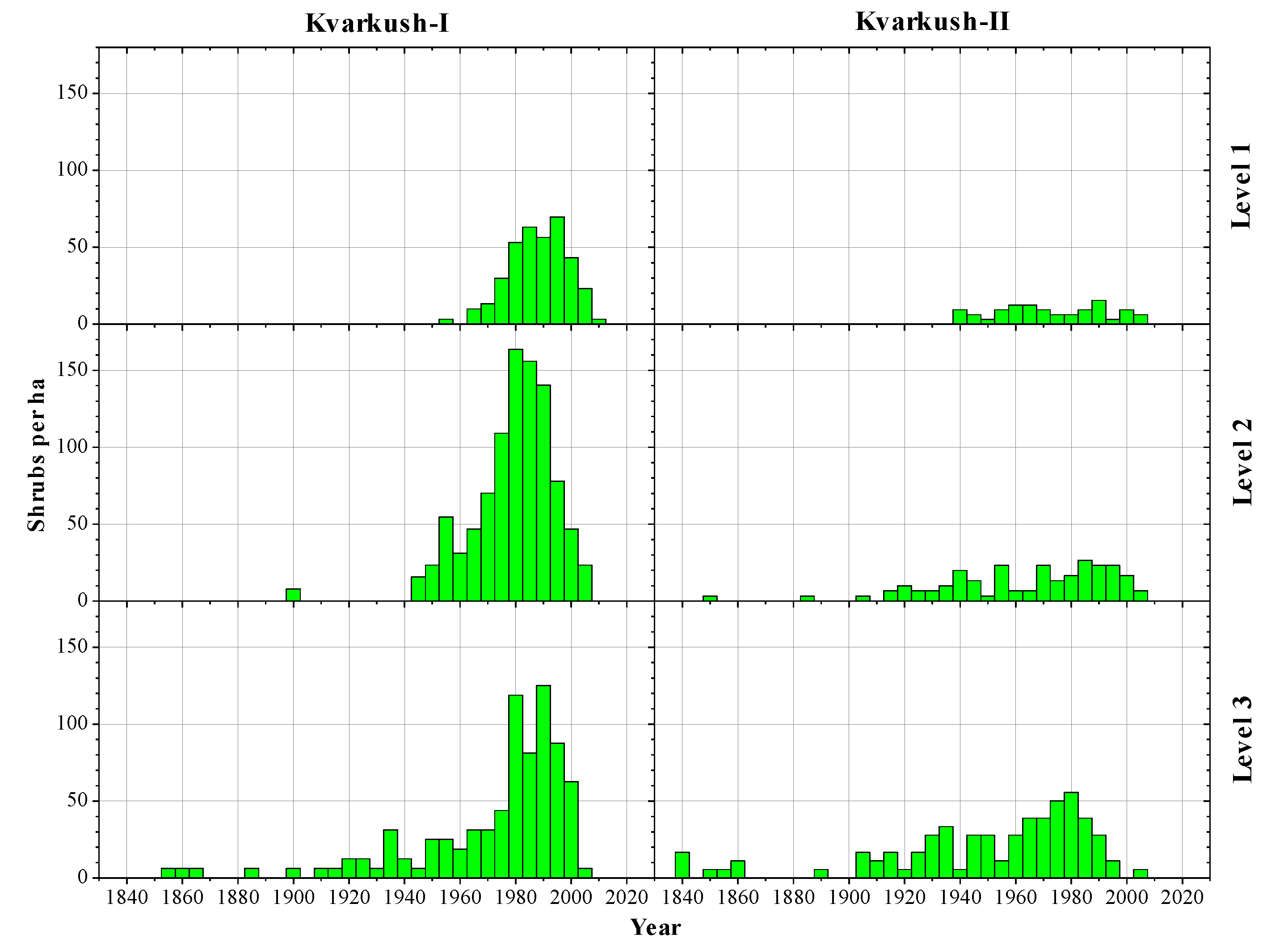
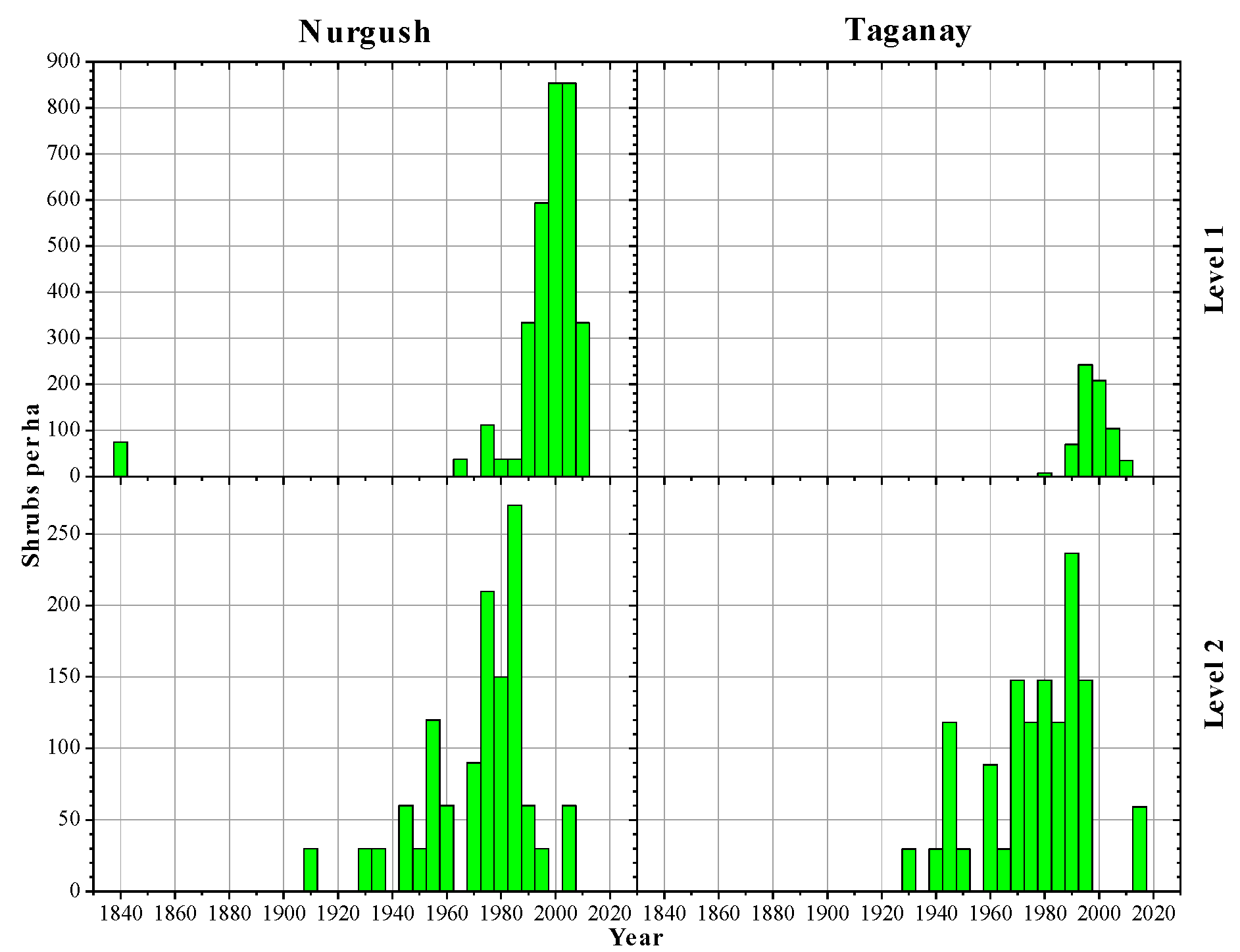
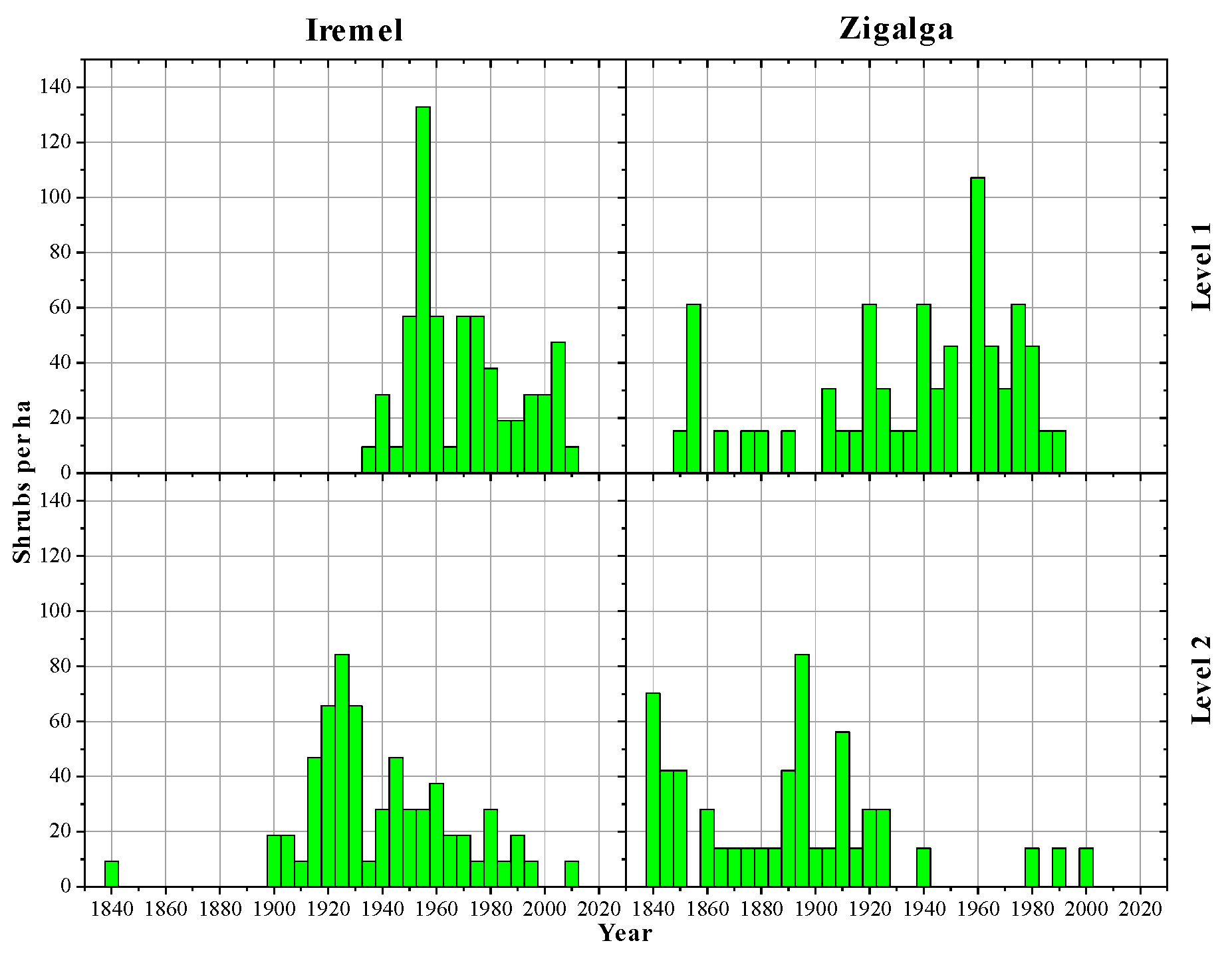
3.1. Altitudinal Changes in Shrub Size and Density
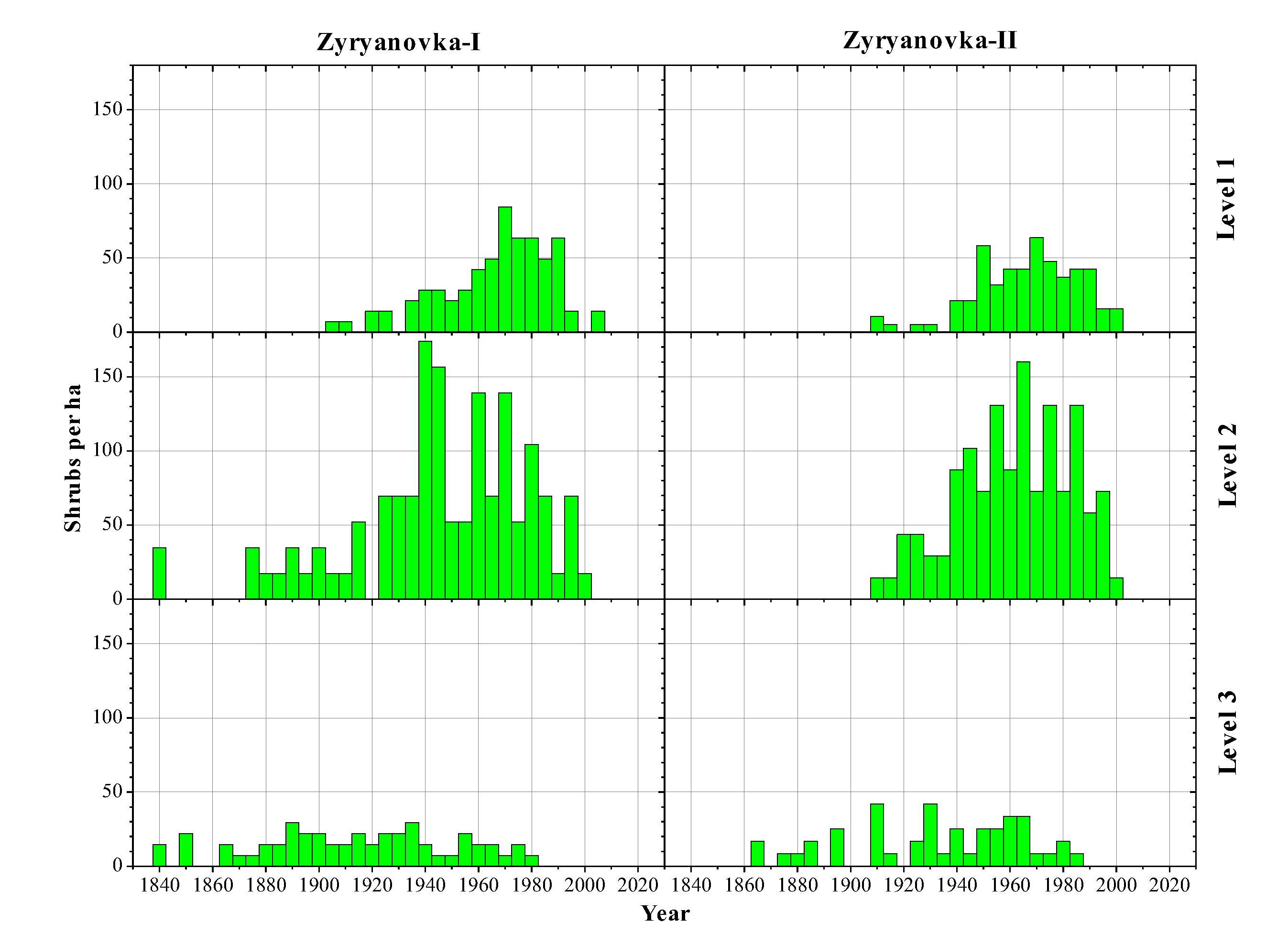
3.2. Age Structures of Shrubs and Trees
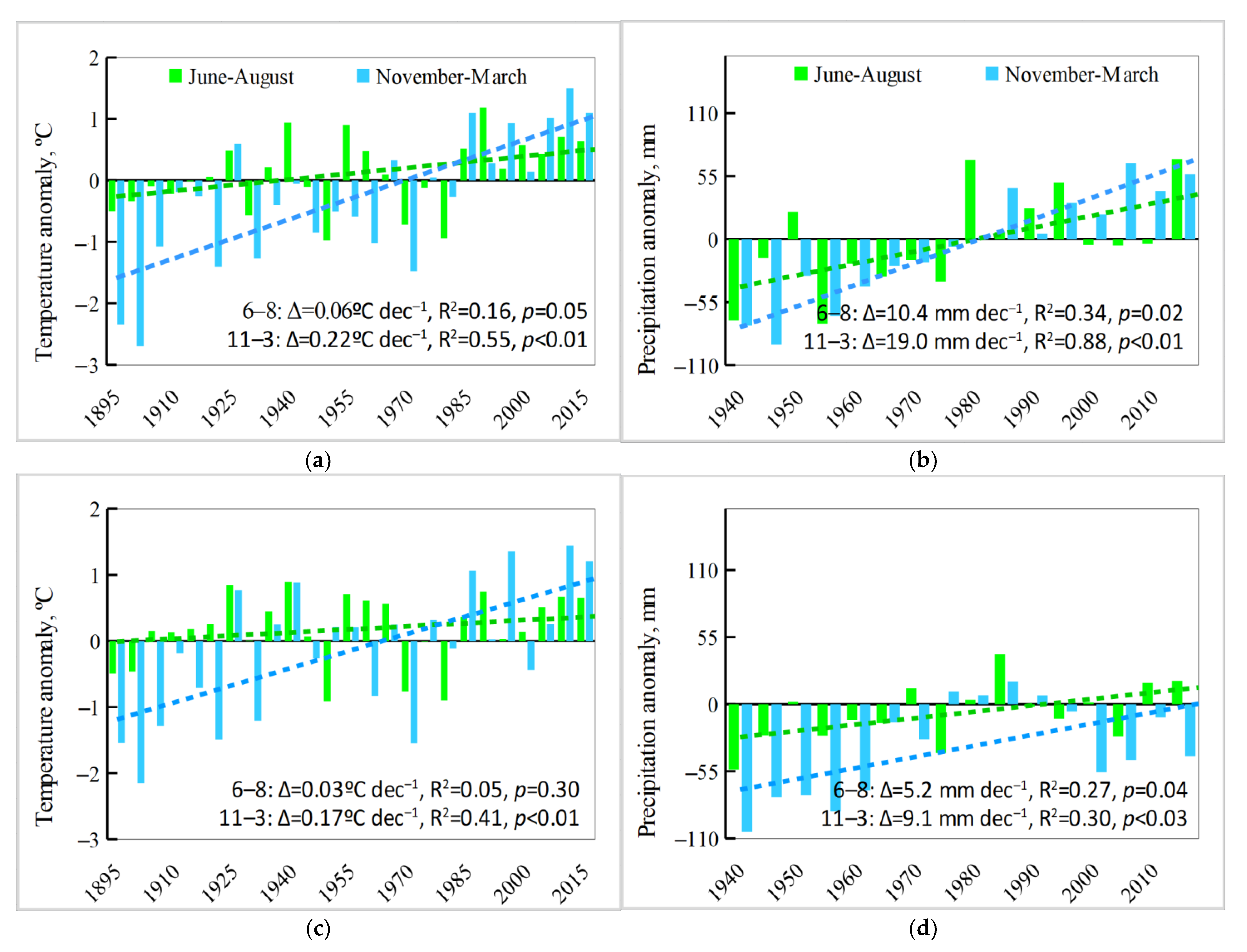
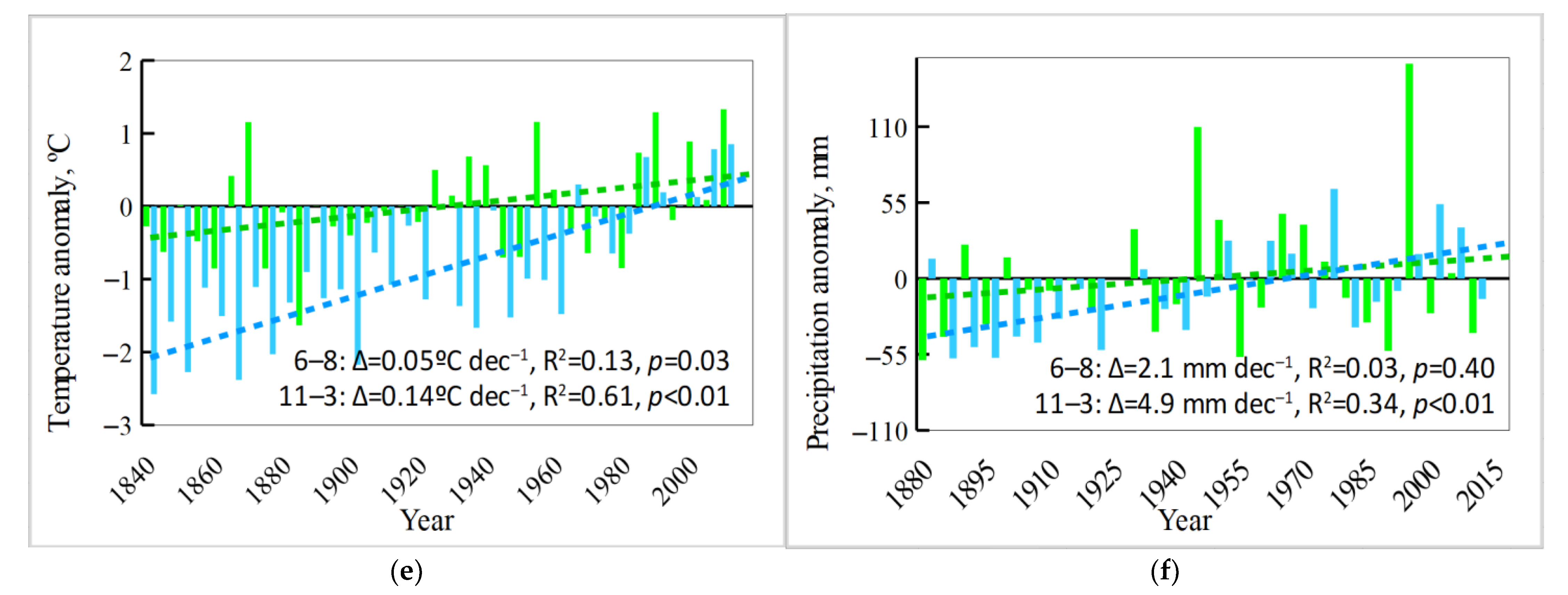
3.3. Analysis of Climate Trends
3.4. Relationships between Climate Variability and Shrub Recruitment Depend on Site and Altitude
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. 2021: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Chapin, F.S.; Sturm, M.; Serreze, M.C.; McFadden, J.P.; Key, J.R.; Lloyd, A.H.; McGuire, A.D.; Rupp, T.S.; Lynch, A.H.; Schimel, J.P.; et al. Role of land-surface changes in arctic summer warming. Science 2005, 310, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Gorchakovskiy, P.L.; Shiyatov, S.G. Phytoindication of Environmental Conditions and Natural Processes in High Mountain Regions; Nauka: Moscow, Russia, 1985. [Google Scholar]
- Kullman, L.; Öberg, L. Post-little ice age tree line rise and climate warming in the Swedish Scandes: A landscape ecological perspective. J. Ecol. 2009, 97, 415–429. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartysh, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’ev, M.; Bugman, H.; et al. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar]
- Wilmking, M.; Sanders, T.G.M.; Zhang, Y.; Kenter, S.; Holzkämper, S.; Crittenden, P.D. Effects of Climate, Site Conditions, and Seed Quality on Recent Treeline Dynamics in NW Russia: Permafrost and Lack of Reproductive Success Hamper Treeline Advance? Ecosystems 2012, 15, 1053–1064. [Google Scholar] [CrossRef]
- Tape, K.; Sturm, M.; Racine, C. The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Chang. Biol. 2006, 12, 686–702. [Google Scholar] [CrossRef]
- Forbes, B.C.; Fauria, M.M.; Zetterberg, P. Russian Arctic warming and “greening” are closely tracked by tundra shrub willows. Glob. Chang. Biol. 2010, 16, 1542–1554. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Blok, D.; Sass-Klaassen, U.; Schaepman-Strub, G.; Heijmans, M.M.P.D.; Sauren, P.; Berendse, F. What are the main climate drivers for shrub growth in Northeastern Siberian tundra? Biogeosciences 2011, 8, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Hellmann, L.; Tegel, W.; Normand, S.; Myers-Smith, I.; Kirdyanov, A.V.; Nievergelt, D.; Schweingruber, F.H. Temperature-induced recruitment pulses of Arctic dwarf shrub communities. J. Ecol. 2015, 103, 489–501. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; MacIas-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Elmendorf, S.C.; Beck, P.S.A.; Wilmking, M.; Hallinger, M.; Blok, D.; Tape, K.D.; Rayback, S.A.; Macias-Fauria, M.; Forbes, B.C.; et al. Climate sensitivity of shrub growth across the tundra biome. Nat. Clim. Chang. 2015, 5, 887–891. [Google Scholar] [CrossRef] [Green Version]
- Naito, A.T.; Cairns, D.M. Patterns and processes of global shrub expansion. Prog. Phys. Geogr. 2011, 35, 423–442. [Google Scholar] [CrossRef]
- Rundqvist, S.; Hedenås, H.; Sandström, A.; Emanuelsson, U.; Eriksson, H.; Jonasson, C.; Callaghan, T.V. Tree and shrub expansion over the past 34 years at the tree-line near Abisko, Sweden. Ambio 2011, 40, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dial, R.J.; Berg, E.E.; Timm, K.; McMahon, A.; Geek, J. Changes in the alpine forest-tundra ecotone commensurate with recent warming in southcentral Alaska: Evidence from Orthophotos and field plots. J. Geophys. Res. Biogeosciences 2007, 112, 1–15. [Google Scholar] [CrossRef]
- Dial, R.J.; Smeltz, S.T.; Sullivan, P.F.; Rinas, C.L.; Timm, K.; Geck, J.E.; Tobin, C.S.; Golden, T.S.; Berg, E.C. Shrubline but not treeline advance matches climate velocity in montane ecosystems of south-central Alaska. Glob. Chang. Biol. 2016, 22, 1841–1856. [Google Scholar] [CrossRef]
- Duchesne, R.R.; Chopping, M.J.; Tape, K.D.; Wang, Z.; Schaaf, C.L.B. Changes in tall shrub abundance on the North Slope of Alaska, 2000–2010. Remote Sens. Environ. 2018, 219, 221–232. [Google Scholar] [CrossRef]
- Terskaia, A.; Dial, R.J.; Sullivan, P.F. Pathways of tundra encroachment by trees and tall shrubs in the western Brooks Range of Alaska. Ecography 2020, 43, 1–10. [Google Scholar] [CrossRef]
- Ropars, P.; Lévesque, E.; Boudreau, S. How do climate and topography influence the greening of the forest-tundra ecotone in northern Québec? A dendrochronological analysis of Betula glandulosa. J. Ecol. 2015, 103, 679–690. [Google Scholar]
- Formica, A.; Farrer, E.C.; Ashton, I.W.; Suding, K.N. Shrub expansion over the past 62 years in Rocky Mountain alpine tundra: Possible causes and consequences. Arct. Antarct. Alp. Res. 2014, 46, 616–631. [Google Scholar] [CrossRef] [Green Version]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 2007, 5, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Wipf, S.; Stoeckli, V.; Bebi, P. Winter climate change in alpine tundra: Plant responses to changes in snow depth and snowmelt timing. Clim. Chang. 2009, 94, 105–121. [Google Scholar] [CrossRef]
- Grigoriev, A.A.; Shalaumova, Y.V.; Erokhina, O.V.; Sokovnina, S.Y.; Vatolina, E.I.; Wilmking, M. Expansion of Juniperus sibirica Burgsd. as a response to climate change and associated effect on mountain tundra vegetation in the Northern Urals. J. Mt. Sci. 2020, 17, 2339–2353. [Google Scholar]
- Shiyatov, S.G.; Moiseev, P.A.; Grigoriev, A.A. Photomonitoring of Tree and Shrub Vegetation in the Highlands of the Southern Urals over the Past 100 Years; USFEU: Ekaterinburg, Russia, 2020. [Google Scholar]
- Kullman, L. A richer, greener and smaller alpine world: Review and projection of warming-induced plant cover change in the Swedish Scandes. Ambio 2010, 39, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, R.G.; Phillips, S.J.; Loranty, M.M.; Beck, P.S.A.; Damoulas, T.; Knight, S.J.; Goetz, S.J. Shifts in Arctic vegetation and associated feedbacks under climate change. Nat. Clim. Chang. 2013, 3, 673–677. [Google Scholar] [CrossRef]
- Liston, G.E.; Mcfadden, J.P.; Sturm, M.; Pielke, R.A. Modelled changes in arctic tundra snow, energy and moisture fluxes due to increased shrubs. Glob. Chang. Biol. 2002, 8, 17–32. [Google Scholar] [CrossRef]
- Sturm, M.; Douglas, T.; Racine, C.; Liston, G. Changing snow and shrub conditions affect albedo with global implications. J. Geophys. Res. 2005, 110, 1–13. [Google Scholar] [CrossRef]
- Sturm, M.; Schimel, J.; Michaelson, G.; Welker, J.; Oberbauer, S.; Liston, G.; Fahnestock, J.; Romanovsky, V.E. Winter Biological Processes Could Help Convert Arctic Tundra to Shrubland. Bioscience 2005, 55, 17–26. [Google Scholar] [CrossRef]
- Cable, J.M.; Barron-Gafford, G.A.; Ogle, K.; Pavao-Zuckerman, M.; Scott, R.L.; Williams, D.G.; Huxman, T.E. Shrub encroachment alters sensitivity of soil respiration to temperature and moisture. J. Geophys. Res. Biogeosciences 2012, 117, 1–11. [Google Scholar] [CrossRef]
- Blok, D.; Heijmans, M.; Schaepman-Strub, G.; Kononov, A.V.; Maximov, T.C.; Berendse, F. Shrub expansion may reduce summer permafrost thaw in Siberian tundra. Glob. Chang. Biol. 2010, 16, 1296–1305. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Hik, D.S. Climate warming as a driver of tundra shrubline advance. J. Ecol. 2018, 106, 547–560. [Google Scholar] [CrossRef] [Green Version]
- Mod, H.K.; Luoto, M. Arctic shrubification mediates the impacts of warming climate on changes to tundra vegetation. Environ. Res. Lett. 2016, 11, 124028. [Google Scholar] [CrossRef] [Green Version]
- Voronchikhina, E.A. Ecological Monitoring in the Vishera State Nature Reserve. In Coordination of Environmental Monitoring in the Specially Protected Natural Areas of the Urals; Ekaterinburg: Ekaterinburg, Russia, 2000; pp. 90–95. [Google Scholar]
- Chuvalsky Kamen Ridge (Stratotype of the Ordovician Chuval Formation). Available online: http://perm-kray.ru/pam010-1.htm (accessed on 8 January 2012).
- Borisevich, D.V. Relief and geological structure. In Ural and Cis-Urals. Series: Natural Conditions and Natural Resources of the USSR; Nauka: Moscow, Russia, 1968; pp. 19–81. [Google Scholar]
- Kuvshinova, K.V. Climate. In Ural and Cis-Urals. Series: Natural Conditions and Natural Resources of the USSR; Nauka: Moscow, Russia, 1968; pp. 82–118. [Google Scholar]
- Korchagin, P.L. Field Geobotany: Methodological Manual; Russian Academy of Sciences Publisher: Moscow, Russia, 1960; Volume 2, pp. 241–248. [Google Scholar]
- Serebryakov, I.G. Ecological Morphology of Plants. Life Forms of Angiosperms and Conifers; Vysshaya Shkola: Moscow, Russia, 1962. [Google Scholar]
- Fritts, H.C. Tree-Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Shiyatov, S.G.; Vaganov, E.A.; Kirdyanov, A.V.; Kruglov, V.B.; Mazepa, V.S.; Naurzbaev, M.M.; Khantemirov, R.M. Dendrochronological Methods. Part I: Fundamentals of Dendrochronology. Collection and Obtaining of Tree Ring Information; Krasnoyarskij Gosudarstvennyj Universitet: Krasnoyarsk, Russia, 2000. [Google Scholar]
- Hantemirov, R.M.; Shiyatov, S.G.; Gorlanova, L.A. Dendroclimatic study of Siberian juniper (Juniperus sibirica Burgsd.). Dendrochronologia 2011, 29, 119–122. [Google Scholar] [CrossRef]
- Surso, M.V.; Barzut, O.S. Features of the growth and development of conifers in the Bolshezemelskaya tundra. Juniper in the Pym-Va-Shor tract. Bull. Moscow State Univ. For. Lesn. Bull. 2010, 5, 18–21. [Google Scholar]
- Zyryanova, Y.V.; Ayoshina, E.N.; Velichko, N.A. Overcoming the deep physiological dormancy of the embryos of the Siberian juniper in vitro. Conifers Boreal Area 2016, 1, 38–43. [Google Scholar]
- Ural UGMS. Monthly Meteorological Tables. Annual, 13vols.
- Bogdanova, E.G.; Golubev, V.S.; Ilyin, B.M.; Dragomilova, I.V. A new model for correcting measured precipitation and its application in the polar regions of the Russian Federation. Meteorol. Hydrol. 2002, 10, 68–93. [Google Scholar]
- Bogdanova, E.G.; Gavrilova, S.Y. Elimination of the inhomogeneity of precipitation time series caused by the replacement of the rain gauge with Nifer protection by the Tretyakov rain gauge. Meteorol. Hydrol. 2008, 8, 87–102. [Google Scholar]
- Gamache, I.; Payette, S. Latitudinal response of subarctic tree lines to recent climate change in eastern Canada. J. Biogeogr. 2005, 32, 849–862. [Google Scholar] [CrossRef]
- Pellizzari, E.; Pividori, M.; Carrer, M. Winter precipitation effect in a mid-latitude temperature-limited environment: The case of common juniper at high elevation in the Alps. Environ. Res. Lett. 2014, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Carrer, M.; Pellizzari, E.; Prendin, A.L.; Pividori, M.; Brunetti, M. Winter precipitation—Not summer temperature—Is still the main driver for Alpine shrub growth. Sci. Total Environ. 2019, 682, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Holtmeier, F.-K. Mountain Timberlines: Ecology, Patchiness, and Dynamics. Advanced Global Change Resource; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Wipf, S.; Rixen, C. A review of snow manipulation experiments in Arctic and alpine tundra ecosystems. Polar Res. 2010, 29, 95–109. [Google Scholar] [CrossRef]
- Wahren, C.H.A.; Walker, M.D.; Bret-Harte, M.S. Vegetation responses in Alaskan arctic tundra after 8 years of a summer warming and winter snow manipulation experiment. Glob. Chang. Biol. 2005, 11, 537–552. [Google Scholar] [CrossRef]
- Bokhorst, S.F.; Bjerke, J.W.; Tømmervik, H.; Callaghan, T.V.; Phoenix, G.K. Winter warming events damage sub-Arctic vegetation: Consistent evidence from an experimental manipulation and a natural event. J. Ecol. 2009, 97, 1408–1415. [Google Scholar] [CrossRef]
- Schmidt, N.M.; Baittinger, C.; Kollmann, J.; Forchhammer, M.C. Consistent dendrochronological response of the dioecious Salix arctica to variation in local snow precipitation across gender and vegetation types. Arctic Antarct. Alp. Res. 2010, 42, 471–475. [Google Scholar] [CrossRef] [Green Version]
- Myers-Smith, I.H.; Hik, D.S. Shrub canopies influence soil temperatures but not nutrient dynamics: An experimental test of tundra snow–shrub interactions. Ecol. Evol. 2013, 3, 3683–3700. [Google Scholar] [CrossRef]
- Baptist, F.; Yoccoz, N.G.; Choler, P. Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient. Plant Soil 2010, 328, 397–410. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bilbrough, C.; Welker, J.M. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biol. Biochem. 2004, 36, 217–227. [Google Scholar] [CrossRef]
- Sturm, M.; McFadden, J.P.; Liston, G.E.; Stuart Chapin, F.; Racine, C.H.; Holmgren, J. Snow-shrub interactions in Arctic Tundra: A hypothesis with climatic implications. J. Clim. 2001, 14, 336–344. [Google Scholar] [CrossRef]
- Sveinbjornsson, B.; Kauhanen, H.; Nordell, O. Treeline ecology of mountain birch in the Tornetrask area. Ecol. Bull. 1996, 45, 65–70. [Google Scholar]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder soils in a warmer world: A snow manipulation study in a northern hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Hollesen, J.; Buchwal, A.; Rachlewicz, G.; Hansen, B.U.; Hansen, M.O.; Stecher, O.; Elberling, B. Winter warming as an important co-driver for Betula nana growth in western Greenland during the past century. Glob. Chang. Biol. 2015, 21, 2410–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantanen, M.; Karpechko, A.Y.; Lipponen, A.; Nordling, K.; Hyvärinen, O.; Ruosteenoja, K.; Vihma, T.; Laaksonen, A. The Arctic has warmed nearly four times faster than the globe since 1979. Comm. Earth Env. 2022, 3, 168. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Variables | Northern Urals | Southern Urals | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Molebnyi Kamen’ | Zyryanovka I | Zyryanovka II | Kvarkush I | Kvarkush II | Taganay | Nurgush | Zigalga | Iremel | ||
| Coordinates | 61°18′52″ N59°13′51″ E | 60°59′24″ N58°58′05″ E | 60°58′16″ N58°58′23″ E | 60°08′49″ N58°44′56″ E | 60°08′47″ N58°41′51″ E | 55°22′03″ N59°54′30″ E | 54°48′35″ N59°07′18″ E | 54°39′46″ N58°39′47″ E | 54°31′49″ N58°51′16″ E | |
| Altitude (m a.s.l.) | High | 890 | 835 | 840 | 907 | 921 | 1090 | 1240 | 1286 | 1301 |
| Mid | - | 820 | 820 | 881 | 907 | 1080 | 1235 | 1260 | 1297 | |
| Low | - | 790 | 790 | 863 | 886 | - | - | - | - | |
| Exposition | High | summit | summit | summit | summit | NE | summit | summit | summit | summit |
| Mid | - | SW | SW | NW | NE | S | SE | SE | SW | |
| Low | - | SW | SW | NW | NE | - | - | - | - | |
| Snow depth (cm) | High | no date | 73.8 ± 8.9 | no date | no date | no date | 15.1 ± 7.2 | 25.0 ± 9.6 | 21.6 ± 11.8 | 13.2 ± 7.3 |
| Mid | - | 87.0 ± 17.1 | no date | no date | no date | 39.6 ± 23.0 | 111.2 ± 30.1 | 70.9 ± 33.0 | 40.9 ± 16.4 | |
| Low | - | 122.5 ± 14.2 | no date | no date | no date | - | - | - | - | |
| Variables | Altitudinal Level | Northern Urals | Southern Urals | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Molebnyi Kamen’ | Zyryanovka I | Zyryanovka II | Kvarkush I | Kvarkush II | Taganay | Nurgush | Zigalga | Iremel | ||
| Height (m) | High | 0.49 ± 0.15 | 0.36 ± 0.02 | 0.39 ± 0.02 | 0.44 ± 0.02 | 0.44 ± 0.02 | 0.13 ± 0.01 | 0.15 ± 0.01 | 0.20 ± 0.02 | 0.22 ± 0.02 |
| Mid | - | 0.56 ± 0.02 | 0.40 ± 0.02 | 0.48 ± 0.03 | 0.76 ± 0.04 | 0.31 ± 0.02 | 0.24 ± 0.01 | 0.71 ± 0.03 | 0.27 ± 0.01 | |
| Low | - | 0.88 ± 0.03 | 0.82 ± 0.02 | 0.66 ± 0.03 | 0.95 ± 0.04 | - | - | - | - | |
| Crown diameter (m) | High | 1.26 ± 0.49 | 0.87 ± 0.05 | 1.18 ± 0.06 | 0.66 ± 0.03 | 1.19 ± 0.10 | 0.41 ± 0.02 | 0.36 ± 0.02 | 1.79 ± 0.12 | 1.24 ± 0.07 |
| Mid | - | 1.55 ± 0.08 | 1.17 ± 0.05 | 0.76 ± 0.04 | 1.47 ± 0.11 | 0.75 ± 0.07 | 1.21 ± 0.09 | 3.53 ± 0.21 | 2.07 ± 0.12 | |
| Low | - | 3.84 ± 0.23 | 3.63 ± 0.17 | 1.26 ± 0.09 | 1.73 ± 0.14 | - | - | - | - | |
| Density (ind ha−1) | High | 417 | 614 | 510 | 738 | 234 | 665 | 3270 | 720 | 625 |
| Mid | - | 1600 | 1366 | 967 | 488 | 1300 | 1230 | 590 | 619 | |
| Low | - | 420 | 476 | 782 | 540 | - | - | - | - | |
| Region | Site | Altitudinal Level | Precipitation | Temperature | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Corresponding Period | Shifted Period | Corresponding Period | Shifted Period | |||||||
| Cold | Warm | Cold | Warm | Cold | Warm | Cold | Warm | |||
| Northern Urals | Molebnyi Kamen’ | High | 0.12 | −0.46 | 0.27 | 0.16 | 0.06 | −0.07 | −0.07 | −0.18 |
| Zyryanovka I | High | 0.59 | 0.21 | 0.71 | 0.42 | 0.07 | 0.01 | −0.35 | −0.46 | |
| Mid | −0.25 | −0.15 | −0.16 | −0.10 | 0.46 | 0.09 | 0.17 | −0.12 | ||
| Low | −0.41 | −0.68 | −0.18 | −0.24 | 0.10 | 0.16 | −0.06 | 0.02 | ||
| All | −0.11 | −0.20 | 0.23 | 0.19 | 0.39 | 0.14 | 0.07 | −0.30 | ||
| Zyryanovka II | High | 0.21 | 0.31 | 0.70 | 0.44 | −0.08 | −0.25 | −0.18 | −0.28 | |
| Mid | 0.03 | −0.45 | 0.46 | −0.04 | 0.39 | 0.11 | 0.02 | −0.35 | ||
| Low | −0.36 | −0.30 | −0.08 | −0.29 | 0.21 | 0.05 | −0.15 | 0.02 | ||
| All | −0.01 | −0.27 | 0.59 | 0.09 | 0.47 | 0.15 | 0.09 | −0.20 | ||
| Kvarkush I | High | 0.79 | 0.81 | 0.86 * | 0.57 * | 0.21 | 0.04 | −0.05 * | 0.06 * | |
| Mid | 0.59 | 0.34 | 0.63 * | 0.30 * | 0.16 | 0.11 | 0.04 * | −0.46 * | ||
| Low | 0.62 | 0.60 | 0.72 * | 0.42 * | 0.25 | 0.25 | 0.30 * | −0.02 * | ||
| All | 0.78 | 0.57 | 0.80 * | 0.40 * | 0.30 | 0.24 | 0.31 * | 0.09 * | ||
| Kvarkush II | High | −0.07 | −0.34 | 0.24 * | −0.04 * | −0.25 | 0.61 | 0.69 * | 0.00 * | |
| Mid | 0.26 | 0.13 | 0.64 * | 0.53 * | 0.20 | 0.43 | 0.40 * | −0.02 * | ||
| Low | 0.18 | 0.12 | −0.11 * | −0.45 * | −0.08 | −0.43 | −0.02 * | −0.24 * | ||
| All | 0.23 | 0.13 | 0.17 * | −0.27 * | 0.02 | −0.01 | 0.28 * | −0.06 * | ||
| Southern Urals | Taganay | High | 0.77 | 0.60 | 0.60 | 0.49 | −0.09 | −0.03 | 0.09 | −0.09 |
| Low | −0.36 | 0.27 | 0.38 | 0.23 | −0.08 | −0.32 | 0.00 | −0.14 | ||
| All | 0.03 | 0.18 | 0.48 | 0.14 | 0.16 | −0.11 | 0.38 | 0.05 | ||
| Nurgush | High | 0.66 | −0.15 | −0.03 | −0.12 | 0.08 | 0.65 | 0.55 | 0.39 | |
| Low | −0.20 | −0.24 | 0.08 | −0.24 | 0.04 | −0.09 | −0.01 | 0.13 | ||
| All | 0.43 | −0.13 | 0.24 | −0.13 | 0.42 | 0.19 | 0.50 | 0.29 | ||
| Zigalga | High | 0.03 | 0.27 | 0.11 | 0.24 | 0.01 | −0.16 | 0.23 | −0.24 | |
| Low | −0.50 | −0.35 | −0.31 | 0.14 | −0.41 | −0.11 | −0.28 | −0.13 | ||
| All | −0.21 | 0.08 | −0.04 | 0.36 | −0.24 | −0.20 | 0.05 | −0.38 | ||
| Iremel | High | 0.16 | −0.07 | 0.33 | −0.26 | 0.01 | −0.07 | 0.06 | 0.07 | |
| Low | −0.38 | 0.15 | 0.20 | 0.25 | −0.11 | −0.16 | −0.38 | −0.17 | ||
| All | 0.14 | 0.05 | 0.45 | −0.05 | 0.13 | −0.05 | −0.03 | 0.02 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigoriev, A.A.; Shalaumova, Y.V.; Balakin, D.S.; Erokhina, O.V.; Abdulmanova, S.Y.; Moiseev, P.A.; Camarero, J.J. Alpine Shrubification: Juniper Encroachment into Tundra in the Ural Mountains. Forests 2022, 13, 2106. https://doi.org/10.3390/f13122106
Grigoriev AA, Shalaumova YV, Balakin DS, Erokhina OV, Abdulmanova SY, Moiseev PA, Camarero JJ. Alpine Shrubification: Juniper Encroachment into Tundra in the Ural Mountains. Forests. 2022; 13(12):2106. https://doi.org/10.3390/f13122106
Chicago/Turabian StyleGrigoriev, Andrey A., Yulia V. Shalaumova, Dmitriy S. Balakin, Olga V. Erokhina, Svetlana Yu. Abdulmanova, Pavel A. Moiseev, and Jesús Julio Camarero. 2022. "Alpine Shrubification: Juniper Encroachment into Tundra in the Ural Mountains" Forests 13, no. 12: 2106. https://doi.org/10.3390/f13122106
APA StyleGrigoriev, A. A., Shalaumova, Y. V., Balakin, D. S., Erokhina, O. V., Abdulmanova, S. Y., Moiseev, P. A., & Camarero, J. J. (2022). Alpine Shrubification: Juniper Encroachment into Tundra in the Ural Mountains. Forests, 13(12), 2106. https://doi.org/10.3390/f13122106