
Effects of Larix olgensis Henry Stumps and Coarse Roots on Phosphorus Fractions and Availability in Plantation Microsite Soils


Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Study Site
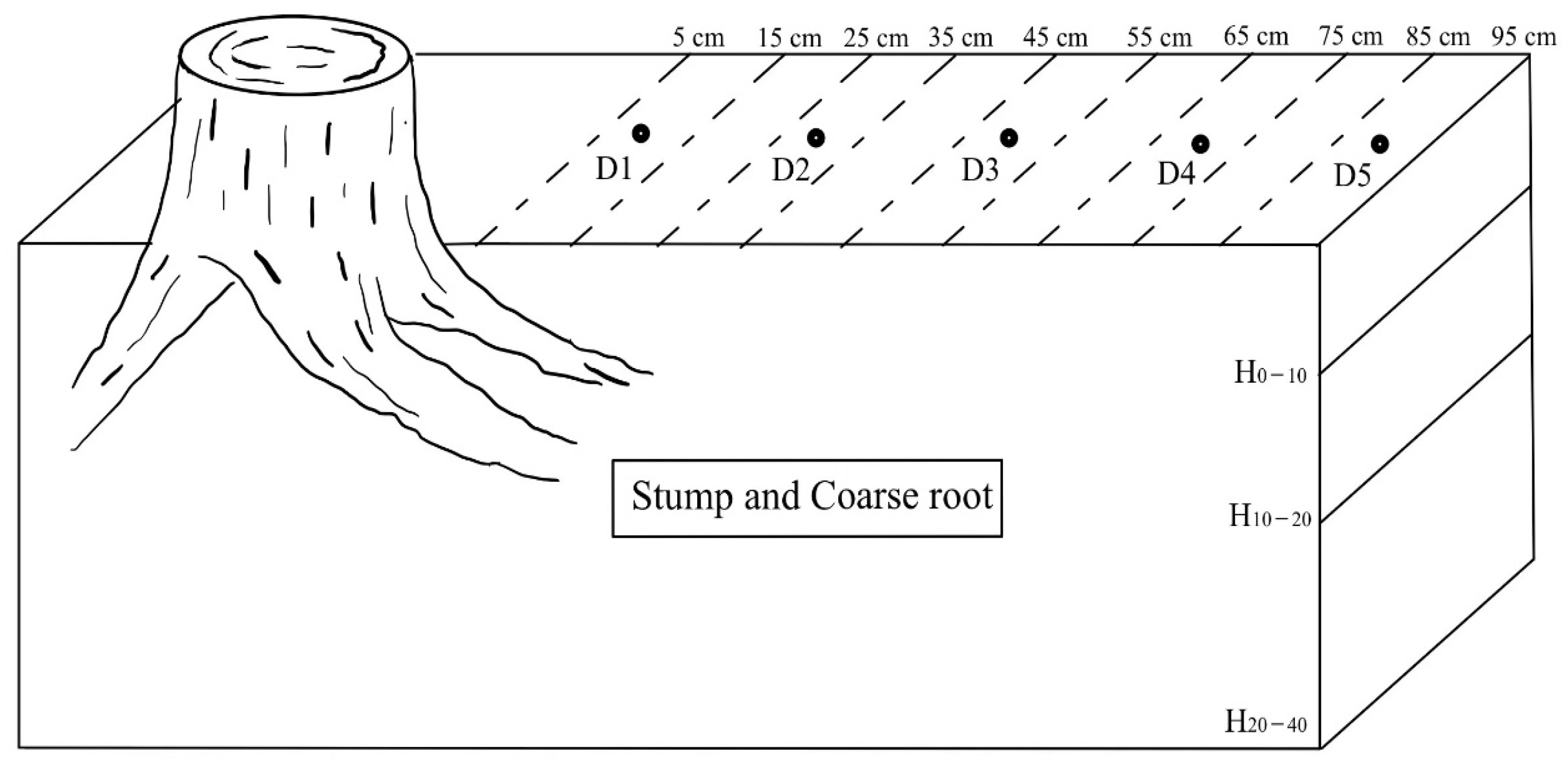
2.2. Soil Sampling
2.3. Laboratory Analysis
2.4. Statistical Analysis
3. Results
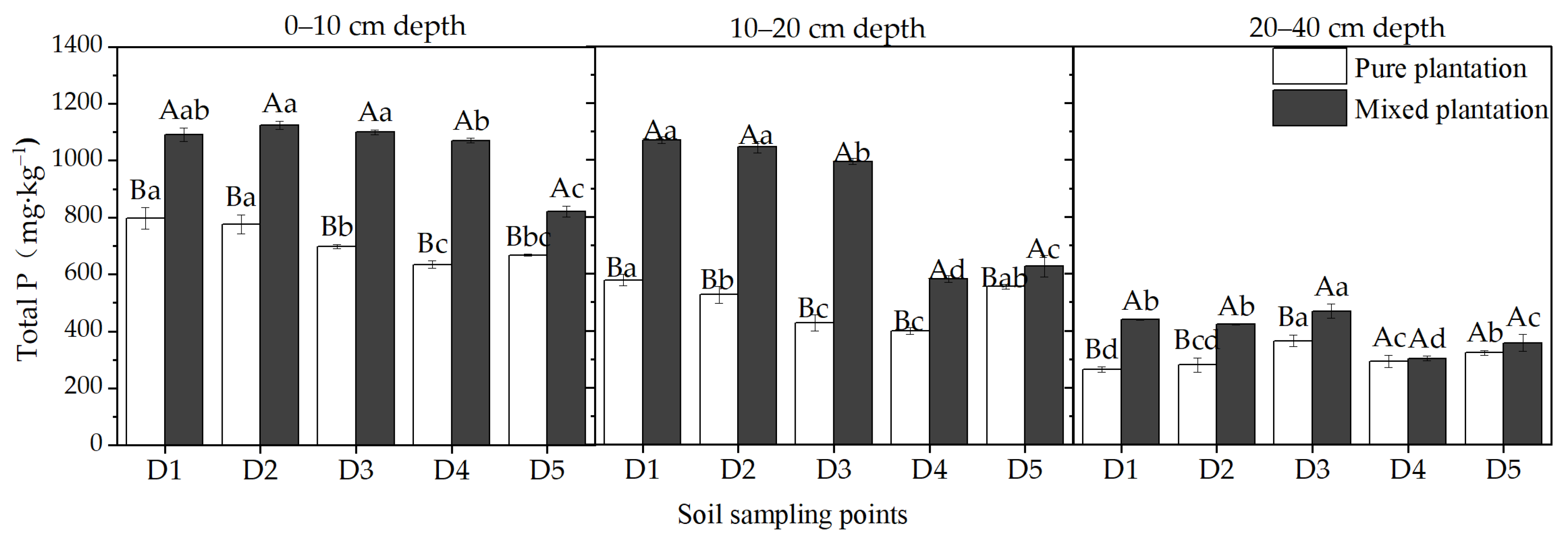
3.1. Soil Total P
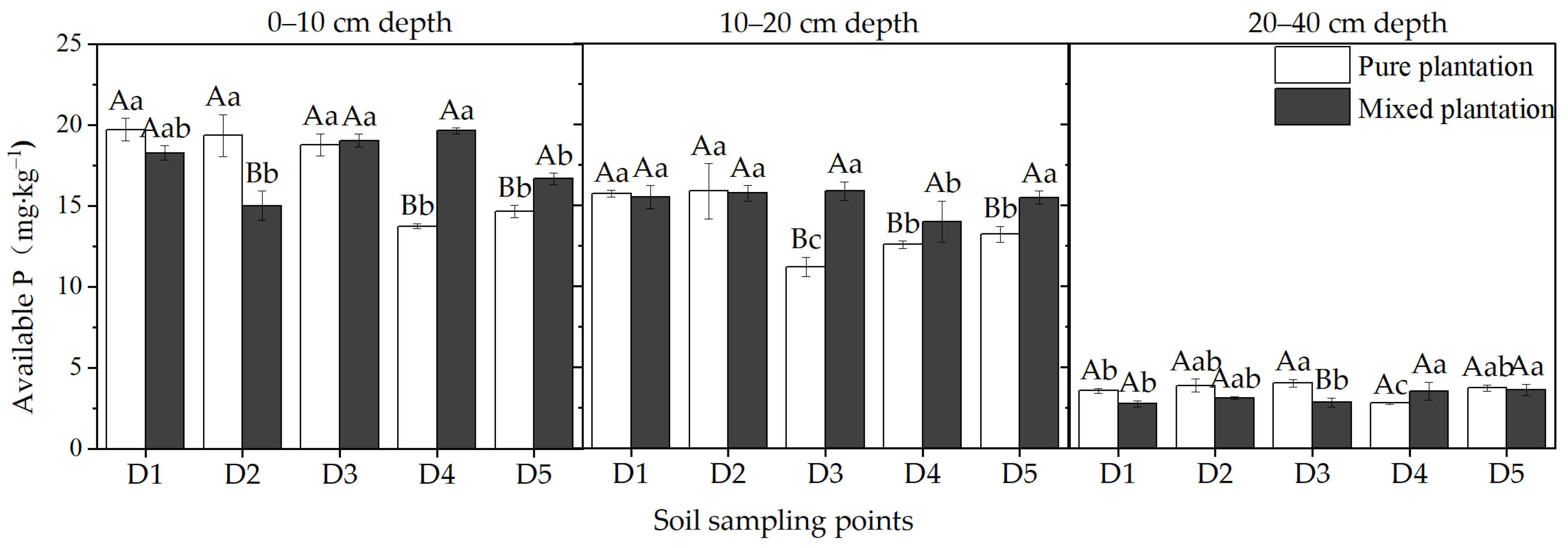
3.2. Soil Available P
3.3. Soil P Fractions
4. Discussion
4.1. Effect of Stumps and Coarse Roots on Microsite Soil P and Availability
4.2. Effects of Different Plantation Established Species on Microsite Soil P
4.3. Limitations of Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oburger, E.; Jones, D.L.; Wenzel, W.W. Phosphorus saturation and pH differentially regulate the efficiency of organic acid anion-mediated P solubilization mechanisms in soil. Plant Soil 2011, 341, 363–382. [Google Scholar] [CrossRef]
- Blake, L.; Johnston, A.E.; Poulton, P.R.; Goulding, K.W.T. Changes in soil phosphorus fractions following positive and negative phosphorus balances for long periods. Plant Soil 2003, 254, 245–261. [Google Scholar] [CrossRef]
- Crews, T.E.; Kitayama, K.; Fownes, J.H.; Riley, R.H.; Herbert, D.A.; Muellerdombois, D.; Vitousek, P.M. Changes in Soil-Phosphorus Fractions and Ecosystem Dynamics across a Long Chronosequence in Hawaii. Ecology 1995, 76, 1407–1424. [Google Scholar] [CrossRef]
- Guppy, C.N.; Menzies, N.W.; Moody, P.W.; Compton, B.L.; Blamey, F.P.C. Analytical methods and quality assuranceA simplified, sequential, phosphorus fractionation method. Commun. Soil Sci. Plan. 2008, 31, 1981–1991. [Google Scholar] [CrossRef]
- Han, Y.; Chen, X.; Choi, B. Effect of freeze–thaw cycles on phosphorus fractions and their availability in biochar-amended Mollisols of northeast China (laboratory experiment). Sustainability 2019, 11, 1006. [Google Scholar] [CrossRef] [Green Version]
- Sui, Y.B.; Thompson, M.L.; Shang, C. Fractionation of phosphorus in a mollisol amended with biosolids. Soil Sci. Soc. Am. J. 1999, 63, 1174–1180. [Google Scholar] [CrossRef]
- Zhang, X.; Gu, H.Y.; Chen, X.W. Effects of selective cutting on soil phosphorus forms and availability in Korean pine broad-leaved forest in Xiaoxing’an Mountains of China. Chin. J. Appl. Ecol. 2018, 29, 441–448. [Google Scholar] [CrossRef]
- Yang, X.; Post, W.M. Phosphorus transformations as a function of pedogenesis: A synthesis of soil phosphorus data using Hedley fractionation method. Biogeosciences 2011, 8, 2907–2916. [Google Scholar] [CrossRef] [Green Version]
- Doolette, A.; Smernik, R. Soil organic phosphorus speciation using spectroscopic techniques. In Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–36. [Google Scholar]
- Redel, Y.; Rubio, R.; Godoy, R.; Borie, F. Phosphorus fractions and phosphatase activity in an Andisol under different forest ecosystems. Geoderma 2008, 145, 216–221. [Google Scholar] [CrossRef]
- Yang, X.Y.; Fan, R.Y.; Wang, E.H.; Xia, X.Y.; Chen, X.W. Topsoil phosphorus forms and availability of different soil and water conservation plantations in typical black soil region, northeast China. Chin. J. Appl. Ecol. 2014, 25, 1555–1560. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Fang, X.; Wang, L.F.; Xiang, W.H.; Alharbi, H.A.; Lei, P.F.; Kuzyakov, Y. Regulation of soil phosphorus availability and composition during forest succession in subtropics. Forest Ecol. Manag. 2021, 502, 119706. [Google Scholar] [CrossRef]
- McGrath, D.A.; Duryea, M.L.; Cropper, W.P. Soil phosphorus availability and fine root proliferation in Amazonian agroforests 6 years following forest conversion. Agr. Ecosyst. Environ. 2001, 83, 271–284. [Google Scholar] [CrossRef]
- Schaap, K.J.; Fuchslueger, L.; Hoosbeek, M.R.; Hofhansl, F.; Martins, N.P.; Valverde-Barrantes, O.J.; Hartley, I.P.; Lugli, L.F.; Quesada, C.A. Litter inputs and phosphatase activity affect the temporal variability of organic phosphorus in a tropical forest soil in the Central Amazon. Plant Soil 2021, 469, 423–441. [Google Scholar] [CrossRef]
- Brandtberg, P.O.; Davis, M.R.; Clinton, P.W.; Condron, L.M.; Allen, R.B. Forms of soil phosphorus affected by stand development of mountain beech (Nothofagus) forests in New Zealand. Geoderma 2010, 157, 228–234. [Google Scholar] [CrossRef]
- Rabinowitsch-Jokinen, R.; Vanha-Majamaa, I. Immediate Effects of Logging, Mounding and Removal of Logging Residues and Stumps on Coarse Woody Debris in Managed Boreal Norway Spruce Stands. Silva. Fenn. 2010, 44, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Błońska, E.; Kacprzyk, M.; Spólnik, A. Effect of deadwood of different tree species in various stages of decomposition on biochemical soil properties and carbon storage. Ecol. Res. 2017, 32, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Hope, G.D. Changes in soil properties, tree growth, and nutrition over a period of 10 years after stump removal and scarification on moderately coarse soils in interior British Columbia. Forest Ecol. Manag. 2007, 242, 625–635. [Google Scholar] [CrossRef]
- Sucre, E.B.; Fox, T.R. Decomposing stumps influence carbon and nitrogen pools and fine-root distribution in soils. Forest Ecol. Manag. 2009, 258, 2242–2248. [Google Scholar] [CrossRef]
- Polglase, P.J.; Attiwill, P.M.; Adams, M.A. Nitrogen and phosphorus cycling in relation to stand age of Eucalyptus regnans F. Muell. III. Labile inorganic and organic P phosphatase activity and P availability. Plant Soil 1992, 142, 177–185. [Google Scholar] [CrossRef]
- Peng, W.; Li, F.R.; Dong, L.H. Individual tree diameter growth model for Larix olgensis plantation in Heilongjiang Province, China. J. Nanjing Forest Univ. 2018, 42, 19–27. [Google Scholar]
- Burger, D.; Shi, D.Z. An introductory comparison of forest ecological conditions in northeast China and Ontario, Canada. For. Chron. 1988, 64, 105–115. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Chen, X. Effects of thinning intensity on carbon storage of Larix olgensis plantation ecosystem. J. Beijing Forest Univ. 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Hunter, J.M.L. Wildlife, Forests, and Forestry. Principles of Managing Forests for Biological Diversity; Prentice Hall: Hoboken, NJ, USA, 1990; p. 370. [Google Scholar]
- Petrillo, M.; Cherubini, P.; Fravolini, G.; Marchetti, M.; Ascher-Jenull, J.; Schärer, M.; Synal, H.; Bertoldi, D.; Camin, F.; Larcher, R.; et al. Time since death and decay rate constants of Norway spruce and European larch deadwood in subalpine forests determined using dendrochronology and radiocarbon dating. Biogeosciences 2016, 13, 1537–1552. [Google Scholar] [CrossRef] [Green Version]
- Kajimoto, T.; Matsuura, Y.; Osawa, A.; Prokushkin, A.S.; Sofronov, M.A.; Abaimov, A.P. Root system development of Larix gmelinii trees affected by micro-scale conditions of permafrost soils in central Siberia. Plant Soil 2003, 255, 281–292. [Google Scholar] [CrossRef]
- Van, E.R.; Verloo, M.; Demeyer, A.; Pauwels, J.M. Manual for the Soil Chemistry and Fertility Laboratory; International Training Centre for Post-Graduate Soil Scientists, Ghent University: Gent, Belgium, 1999; p. 243. [Google Scholar]
- Hedley, M.J.; Stewart, J.W.B.; Chauhan, B.S. Changes in Inorganic and Organic Soil-Phosphorus Fractions Induced by Cultivation Practices and by Laboratory Incubations. Soil Sci. Soc. Am. J. 1982, 46, 970–976. [Google Scholar] [CrossRef]
- Rose, T.J.; Hardiputra, B.; Rengel, Z. Wheat, canola and grain legume access to soil phosphorus fractions differs in soils with contrasting phosphorus dynamics. Plant Soil 2010, 326, 159–170. [Google Scholar] [CrossRef]
- Qin, S.J.; Liu, J.S.; Wang, G.P.; Wang, J.D. Seasonal changes of soil phosphorus fractions under Calamagrostis angustifolia wetlands in Sanjiang Plain, China. Acta Ecol. Sin. 2007, 27, 3844–3851. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta. 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, S.; Zhu, P.; Huang, S.; Wang, B.; Zhao, L.; Xu, M. Characterizing differences in the phosphorus activation coefficient of three typical cropland soils and the influencing factors under long-term fertilization. PLoS ONE 2017, 12, e0176437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Zhang, M.; Zhang, Y.; Li, Y.; Zhang, Y.; Chi, Q.; Pang, H.; Sun, G. The carbon metabolism characteristics and diversity of soil microbial communities from pure or coniferous and broad-leaved mixed forests in the Maoer Mountain region. J. Nanjing Forest Univ. 2017, 60, 81. [Google Scholar]
- Palviainen, M.; Finer, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Phosphorus and base cation accumulation and release patterns in decomposing Scots pine, Norway spruce and silver birch stumps. Forest Ecol. Manag. 2010, 260, 1478–1489. [Google Scholar] [CrossRef]
- Spears, J.D.H.; Lajtha, K. The imprint of coarse woody debris on soil chemistry in the western Oregon Cascades. Biogeochemistry 2004, 71, 163–175. [Google Scholar] [CrossRef]
- Cromack, J.K.; Sollins, P.; Graustein, W.C.; Speidel, K.; Todd, A.W.; Spycher, G.; Li, C.Y.; Todd, R.L. Calcium oxalate accumulation and soil weathering in mats of the hypogeous fungus Hysterangium crassum. Soil Biol. Biochem. 1979, 11, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L. Organic acids in the rhizosphere—A critical review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Pietri, J.C.A.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Thomas, S.J.; Pierzynski, G.M. Chemistry of phosphorus in soils. In Chemical Processes in Soils; Soil Science Society of America: Madison, WI, USA, 2005; pp. 151–192. [Google Scholar]
- Sanyal, S.K.; Dedatta, S.K. Chemistry of Phosphorus Transformations in Soil. In Advances in Soil Science; Springer: New York, NY, USA, 1991; pp. 1–120. [Google Scholar]
- Zhang, H.W.; Huang, Y.; Ye, X.S.; Shi, L.; Xu, F.S. Genotypic differences in phosphorus acquisition and the rhizosphere properties of Brassica napus in response to low phosphorus stress. Plant Soil 2009, 320, 91–102. [Google Scholar] [CrossRef]
- Lustenhouwer, N.; Maynard, D.S.; Bradford, M.A.; Lindner, D.L.; Oberle, B.; Zanne, A.E.; Crowther, T.W. A trait-based understanding of wood decomposition by fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 11551–11558. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E.; Mcclaugherty, C.A.; Rayburn, L.; Repert, D.; Weiland, T. Wood Decomposition: Nitrogen and Phosphorus Dynamics in Relation to Extracellular Enzyme Activity. Ecology 1993, 74, 1586–1593. [Google Scholar] [CrossRef]
- Song, C.; Han, X.Z.; Tang, C. Changes in phosphorus fractions, sorption and release in Udic Mollisols under different ecosystems. Biol. Fert. Soils 2007, 44, 37–47. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Types | Planting Time | Stand Initial Density (Trees∙ha−1) | Stand Density (Trees∙ha−1) | Canopy Coverage (%) | Average Height (m) | Average DBH (cm) | Stand Volume (m3∙ha−1) |
|---|---|---|---|---|---|---|---|
| Pure plantation | 1969 | 4440 | 920 | 79 | 23.5 | 24.1 | 251 |
| Mixed plantation | 1969 | 4440 | 1190 | 84 | 20.2 | 20.6 | 258 |
| Soil Depth | Types of Plantations | D1 | D2 | D3 | D4 | D5 |
|---|---|---|---|---|---|---|
| 0–10 cm | Pure | 2.57 ± 0.12 Aa | 2.62 ± 0.18 Aa | 2.71 ± 0.25 Aa | 2.20 ± 0.11 Ab | 2.23 ± 0.08 Ab |
| Mixed | 1.60 ± 0.12 Bcd | 1.46 ± 0.14 Bd | 1.76 ± 0.08 Bbc | 1.85 ± 0.15 Bab | 2.02 ± 0.09 Aa | |
| 10–20 cm | Pure | 2.70 ± 0.12 Abc | 2.99 ± 0.41 Aab | 2.62 ± 0.20 Ac | 3.16 ± 0.12 Aa | 2.38 ± 0.10 Ac |
| Mixed | 1.45 ± 0.07 Bb | 1.51 ± 0.05 Bb | 1.60 ± 0.07 Bb | 2.41 ± 0.19 Ba | 2.47 ± 0.12 Aa | |
| 20–40 cm | Pure | 1.35 ± 0.06 Aa | 1.40 ± 0.09 Aa | 1.11 ± 0.12 Ab | 0.96 ± 0.06 Ac | 1.16 ± 0.08 Ab |
| Mixed | 0.62 ± 0.04 Bb | 0.74 ± 0.03 Bb | 0.61 ± 0.09 Bb | 1.17 ± 0.15 Aa | 1.04 ± 0.13 Aa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, Y.; Men, X.; Sun, Z.; Chen, X. Effects of Larix olgensis Henry Stumps and Coarse Roots on Phosphorus Fractions and Availability in Plantation Microsite Soils. Forests 2022, 13, 2166. https://doi.org/10.3390/f13122166
Yue Y, Men X, Sun Z, Chen X. Effects of Larix olgensis Henry Stumps and Coarse Roots on Phosphorus Fractions and Availability in Plantation Microsite Soils. Forests. 2022; 13(12):2166. https://doi.org/10.3390/f13122166
Chicago/Turabian StyleYue, Yang, Xiuli Men, Zhihu Sun, and Xiangwei Chen. 2022. "Effects of Larix olgensis Henry Stumps and Coarse Roots on Phosphorus Fractions and Availability in Plantation Microsite Soils" Forests 13, no. 12: 2166. https://doi.org/10.3390/f13122166
APA StyleYue, Y., Men, X., Sun, Z., & Chen, X. (2022). Effects of Larix olgensis Henry Stumps and Coarse Roots on Phosphorus Fractions and Availability in Plantation Microsite Soils. Forests, 13(12), 2166. https://doi.org/10.3390/f13122166