
Plant Functional Trait Responses to Dolomite and Limestone Karst Forests in Southwest China
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Plots Survey and Soil Samples
2.3. Functional Trait Selection
2.4. Data Analysis and Statistics
3. Results
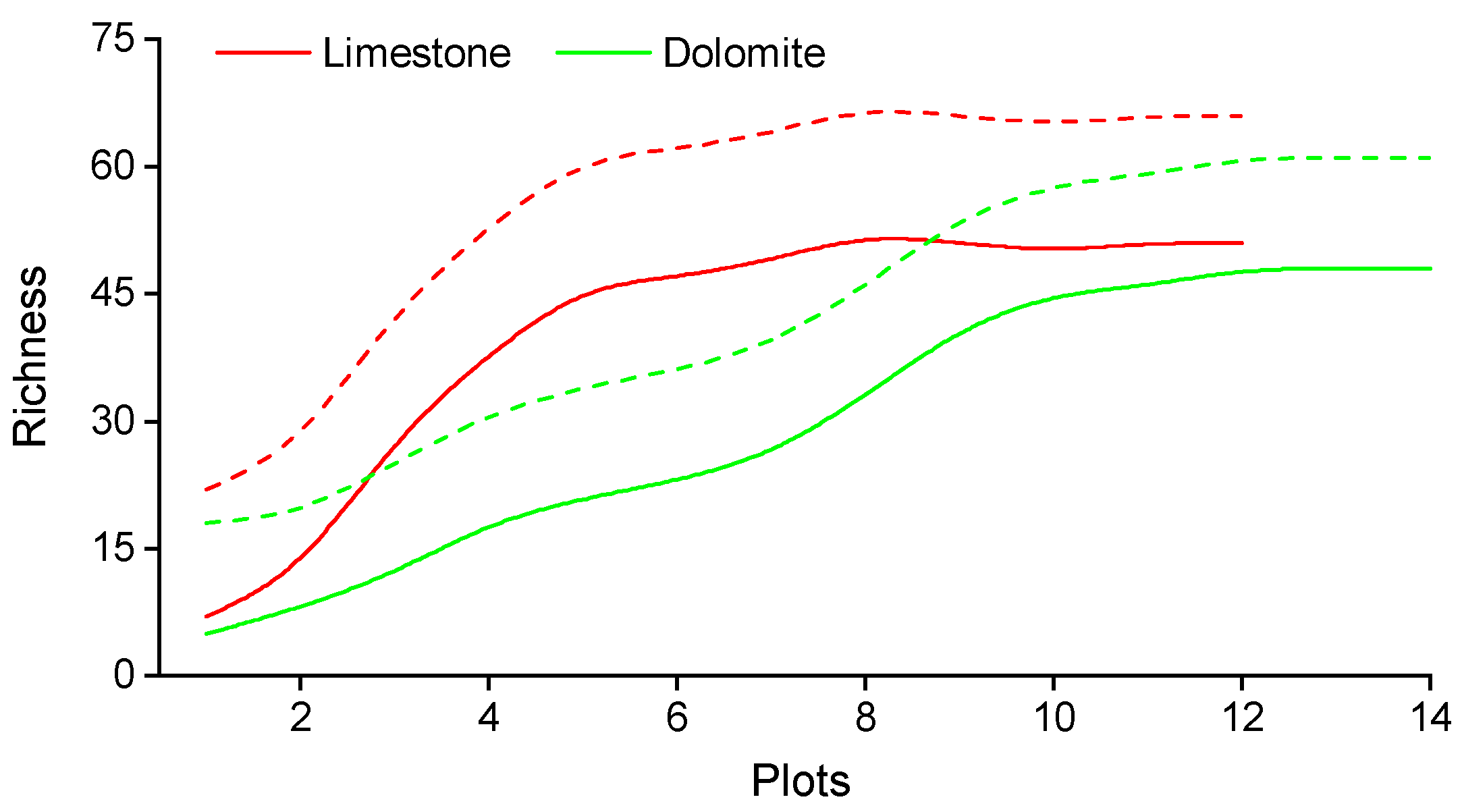
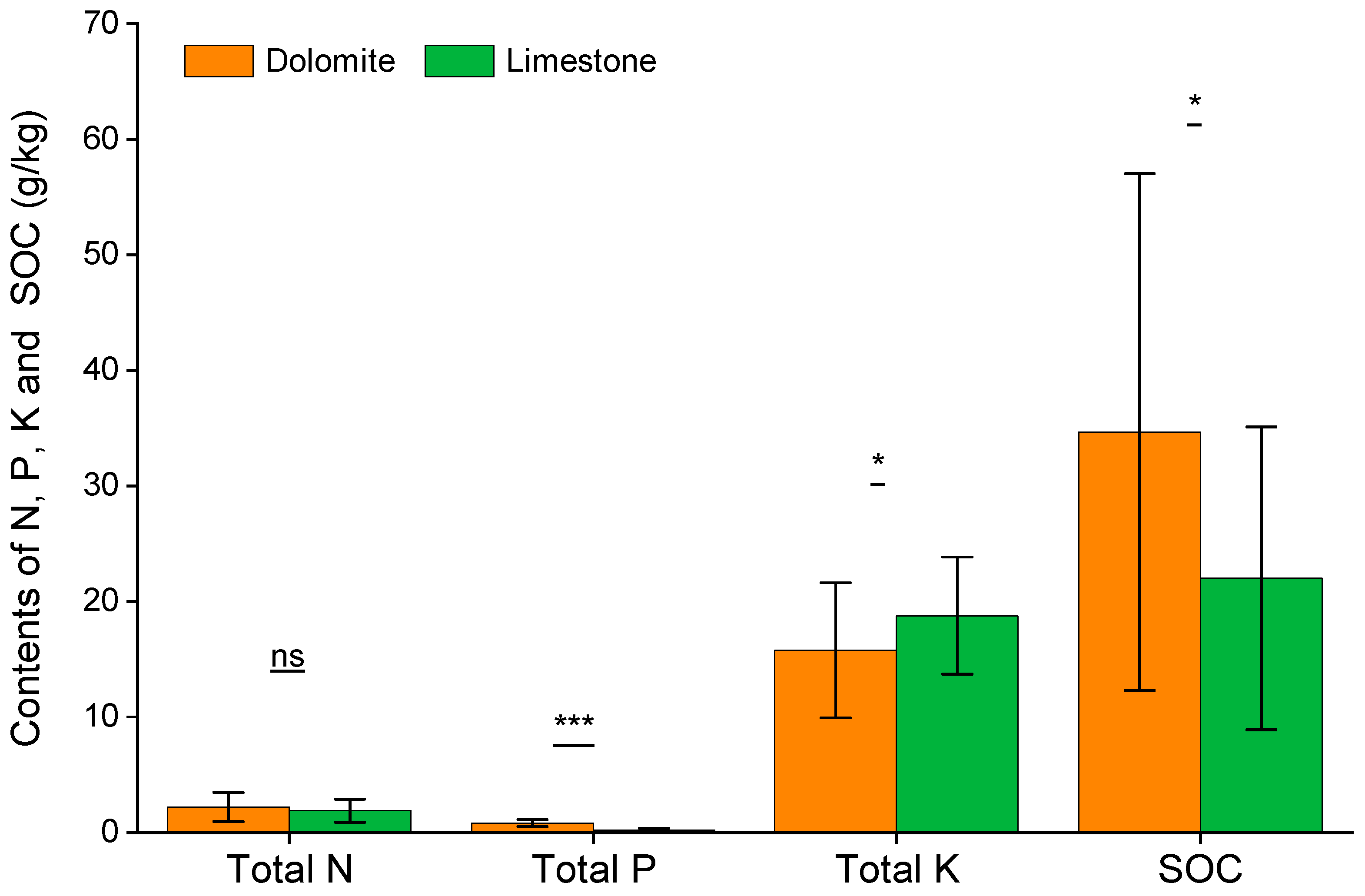
3.1. Species Diversity and Soil Characteristics
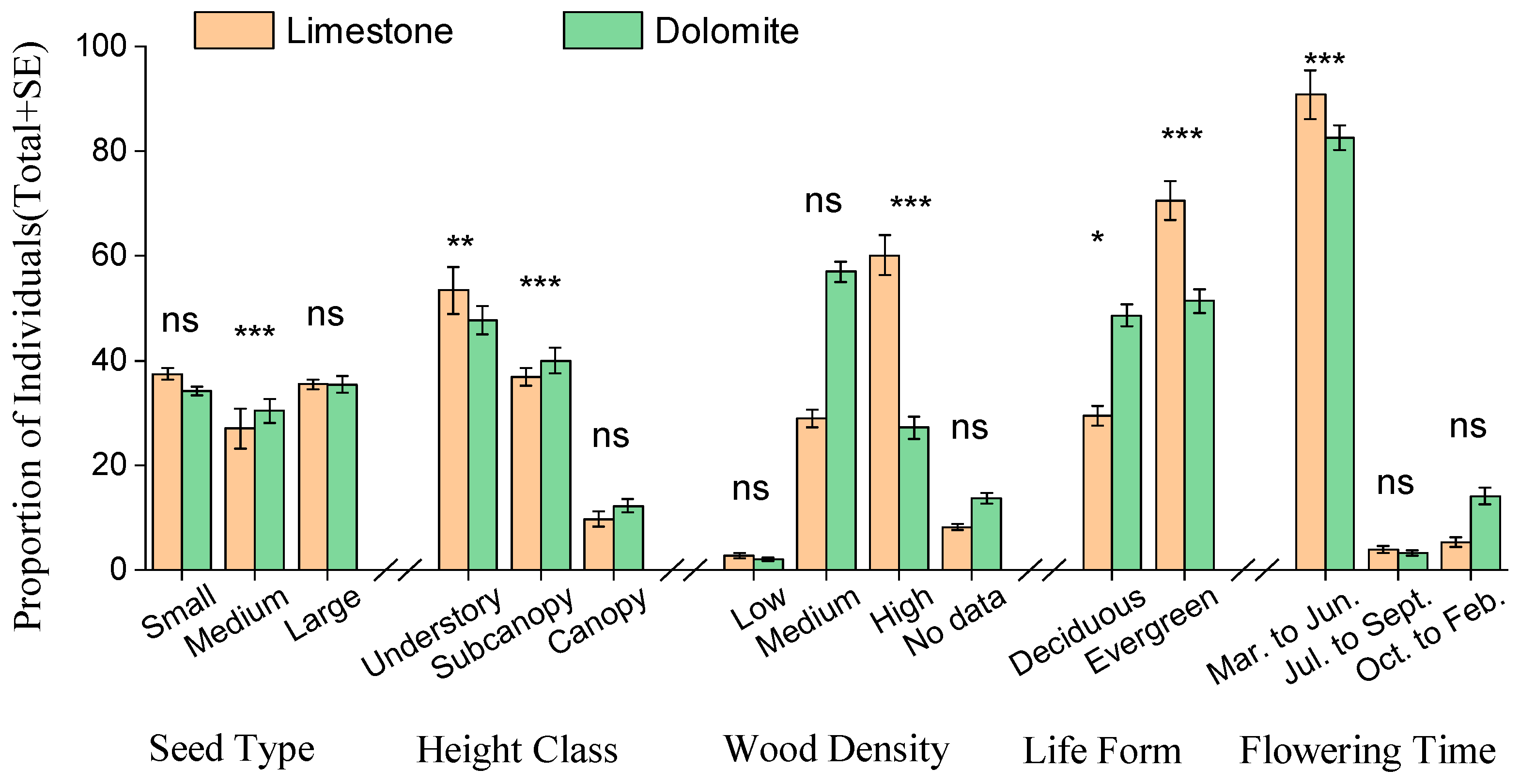
3.2. Functional Trait Representation
3.3. Functional Trait Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Sheng, M.; Xiong, K.; Wang, L.; Li, X.; Li, R. Response of soil physical and chemical properties to Rocky desertification succession in South China Karst. Carbonates Evaporites 2018, 33, 15–28. [Google Scholar] [CrossRef]
- Geekiyanage, N.; Goodale, U.M.; Cao, K.; Kitajima, K. Plant ecology of tropical and subtropical karst ecosystems. Biotropica 2019, 51, 626–640. [Google Scholar] [CrossRef]
- Huang, Y.; Ai, X.; Yao, L.; Zang, R.; Ding, Y.; Huang, J.; Feng, G.; Liu, J. Changes in the diversity of evergreen and deciduous species during natural recovery following clear-cutting in a subtropical evergreen-deciduous broadleaved mixed forest of central China. Trop. Conserv. Sci. 2015, 8, 1033–1052. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chen, X.; Ma, J.; Liang, S.; Huang, J.; Liu, R.; Pan, Y. Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broadleaved mixed forests in karst topography, Guilin, Southwest China. Trop. Conserv. Sci. 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, B.; Mallik, A.U.; Huang, F.; Xiang, W.; Ding, T.; Wen, S.; Lu, S.; Li, D.; He, Y. Topographic species–habitat associations of tree species in a heterogeneous tropical karst seasonal rain forest, China. J. Plant. Ecol. 2017, 3, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ouyang, Z.; Xu, W.; Zheng, H.; Meng, X. Sampling adequacy estimation for plant species composition by accumulation curves—A case study of urban vegetation in Beijing, China. Landsc. Urban Plan. 2010, 95, 113–121. [Google Scholar] [CrossRef]
- Hartmann, J.; Moosdorf, N. The new global lithological map database GLiM: A representation of rock properties at the Earth surface. Geochem. Geophys. Geosystems 2012, 13. [Google Scholar] [CrossRef]
- Li, D.; Wen, L.; Yang, L.; Luo, P.; Xiao, K.; Chen, H.; Zhang, W.; He, X.; Chen, H.; Wang, K. Dynamics of soil organic carbon and nitrogen following agricultural abandonment in a karst region. J. Geophys. Res. Biogeosci. 2017, 122, 230–242. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Hu, G.; Ni, J. Effects of topographical and edaphic factors on the distribution of plant communities in two subtropical karst forests, southwestern China. J. Mt. Sci.-Engl. 2013, 10, 95–104. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Hu, G.; Zhu, J.D.; Luo, D.H.; Ni, J. Spatial patterns and interspecific associations of dominant tree species in two old-growth karst forests, SW China. Ecol. Res. 2010, 25, 1151–1160. [Google Scholar] [CrossRef]
- Guo, Y.; Li, D.; Wang, B.; Bai, K.; Xiang, W.; Li, X. C, N and P stoichiometric characteristics of soil and litter fall for six common tree species in a northern tropical karst seasonal rainforest in Nonggang, Guangxi, southern China. Biodivers. Sci. 2017, 25, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- László, E.; Gy Rgy, K.D.; Zoltán, B.; Bence, K.; Csaba, N.; János, K.P.; Csaba, T. Habitat heterogeneity as a key to high conservation value in forest-grassland mosaics. Biol. Conserv. 2018, 226, 72–80. [Google Scholar]
- Yuan, D. On the karst environmental system. Carsological Sin. 1988, 3, 179–186. [Google Scholar]
- Cao, J.; Yuan, D.; Pan, G. Some soil features in karst ecosystem. Adv. Earth Sci. 2003, 18, 37–44. [Google Scholar]
- Dong, Q.; You, Y.; Luo, W.; Liu, S.; Wang, G.; Liu, Y.; Zhou, J. Species composition and niche differences of dominant populations of plant communities from different parent rocks in karst area. Carsological Sin. 2021, 40, 849–859. [Google Scholar]
- Xu, M.; Ke, X.; Zhang, Y.; Wang, Y.; Su, Z. Wood densities of six hardwood tree species in Eastern Guangdong and Influencing factors. J. South China Agric. Univ. 2016, 37, 100–106. [Google Scholar]
- Zhu, J.; Shi, Y.; Fang, L.; Liu, X.; Ji, C. Patterns and determinants of wood physical properties across major tree species in China. Sci. Sin. Vitae 2015, 45, 56–67. [Google Scholar] [CrossRef]
- Zimmermann, M.H. Xylem Structure and the Ascent of Sap; Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- Cao, K.; Fu, P.; Chen, Y.; Jiang, Y.; Zhu, S. Implications of the Ecophysiological Adaptation of Plants on Tropical Karst Habitats for the Ecological Restoration of Desertified Rocky Lands in Southern China. Sci. Sin. Vitae 2014, 44, 238–247. [Google Scholar]
- Geekiyanage, N.; Goodale, U.M.; Cao, K.; Kitajima, K. Leaf trait variations associated with habitat affinity of tropical karst tree species. Ecol. Evol. 2018, 8, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Shipley, B.; Vile, D.; Garnier, E. From plant traits to plant communities: A statistical mechanistic approach to biodiversity. Science 2006, 314, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Sargent, R.D.; Ackerly, D.D. Plant-pollinator interactions and the assembly of plant communities. Trends. Ecol. Evol. 2008, 23, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, S.; Massai, R.; Iacona, C.; Guerriero, R.; Viti, R. Forty-year investigations on apricot blooming: Evidences of climate change effects. Sci. Hortic-Amst. 2019, 244, 399–405. [Google Scholar] [CrossRef]
- Tao, Z.; Zhong, S.; Ge, Q.; Dai, J.; Xu, Y.; Wang, H. Spatiotemporal variations in flowering duration of woody plants in China from 1963 to 2012. Acta Geogr. Sin. 2017, 72, 53–63. [Google Scholar]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef]
- Fitter, A.H.; Fitter, R.S.R. Rapid Changes in Flowering Time in British Plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef]
- Menzel, A. Phenology: Its Importance to the Global Change Community. Clim. Chang. 2002, 54, 379–385. [Google Scholar] [CrossRef]
- Stephenson, N.L. Climatic Control of Vegetation Distribution: The Role of the Water Balance. Am. Nat. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- Zhong, F.; Xu, X.; Li, Z.; Zeng, X.; Yi, R.; Luo, W.; Zhang, Y.; Xu, C. Relationships between lithology, topography, soil, and vegetation, and their implications for karst vegetation restoration. Catena 2022, 209, 105831. [Google Scholar] [CrossRef]
- Zhang, S.; Bai, X.; Zhao, C.; Tan, Q.; Luo, G.; Cao, Y.; Deng, Y.; Li, Q.; Li, C.; Wu, L.; et al. Limitations of soil moisture and formation rate on vegetation growth in karst areas. Sci. Total Environ. 2022, 810, 151209. [Google Scholar] [CrossRef] [PubMed]
- Furey, G.N.; Tilman, D. Plant biodiversity and the regeneration of soil fertility. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; He, Q.; Liu, Z.; Wu, Z.; Xie, S. Soil nutrient variation impacted by ecological restoration in the different lithological karst area, Shibing, China. Glob. Ecol. Conserv. 2021, 25, e1399. [Google Scholar] [CrossRef]
- Hou, M.; Jiang, Z. Species diversity of karst original forest in different geochemical environments in Maolan. Ecol. Environ. 2006, 3, 572–576. [Google Scholar]
- Brancalion, P.H.; Bello, C.; Chazdon, R.L.; Galetti, M.; Jordano, P.; Lima, R.A.; Medina, A.; Pizo, M.A.; Reid, J.L. Maximizing biodiversity conservation and carbon stocking in restored tropical forests. Conserv. Lett. 2018, 11, e12454. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, X.; Li, Z.; Liu, M.; Xu, C.; Zhang, R.; Luo, W. Effects of vegetation restoration on soil quality in degraded karst landscapes of southwest China. Sci. Total Environ. 2019, 650, 2657–2665. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Soil nutrients and stoichiometric ratios as affected by land use and lithology at county scale in a karst area, southwest China. Sci. Total Environ. 2018, 619–620, 1299–1307. [Google Scholar] [CrossRef]
- Engert, J.E.; Vogado, N.O.; Freebody, K.; Byrne, B.; Murphy, J.; Sheather, G.; Snodgrass, P.; Nugent, L.; Lloyd, D.; Laurance, S.G.W. Functional trait representation differs between restoration plantings and mature tropical rainforest. For. Ecol. Manag. 2020, 473, 118304. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Woodall, C.W.; Perry, C.H.; Miles, P.D. The relative density of forests in the United States. For. Ecol. Manag. 2006, 226, 368–372. [Google Scholar] [CrossRef]
- Bátori, Z.; Erdős, L.; Gajdács, M.; Barta, K.; Tobak, Z.; Frei, K.; Tölgyesi, C. Managing climate change microrefugia for vascular plants in forested karst landscapes. For. Ecol. Manag. 2021, 496, 119446. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, B.; Li, D.; Xiang, W.; Ding, T.; Huang, F.; Li, X. Multivariate relationships between litter productivity and its drivers in a tropical karst seasonal rainforest. Flora 2020, 273, 151728. [Google Scholar] [CrossRef]
- Lu, M.; Du, H.; Song, T.; Peng, W.; Su, L.; Zhang, H.; Zeng, Z.; Wang, K.; Zeng, F. Effects of density dependence in an evergreen-deciduous broadleaf karst forest in southwest China. For. Ecol. Manag. 2021, 490, 119142. [Google Scholar] [CrossRef]
- Ding, Y.; Nie, Y.; Chen, H.; Wang, K.; Querejeta, J.I. Water uptake depth is coordinated with leaf water potential, water-use efficiency and drought vulnerability in karst vegetation. New Phytol. 2021, 229, 1339–1353. [Google Scholar] [CrossRef]
- Tan, F.; Song, H.; Fu, P.; Chen, Y.; Siddiq, Z.; Cao, K.; Zhu, S. Hydraulic safety margins of co-occurring woody plants in a tropical karst forest experiencing frequent extreme droughts. Agr. For. Meteorol. 2020, 292–293, 108107. [Google Scholar] [CrossRef]
- Zhu, S.D.; Chen, Y.J.; Fu, P.L.; Cao, K.F. Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability. Tree Physiol. 2017, 37, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.M.; McCulloh, K.A.; Woodruff, D.R.; Meinzer, F.C. Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Sci. 2012, 195, 48–53. [Google Scholar] [CrossRef]
- Mota, J.F.; Medina-Cazorla, J.M.; Navarro, F.B.; Pérez-García, F.J.; Pérez-Latorre, A.; Sánchez-Gómez, P.; Torres, J.A.; Benavente, A.; Blanca, G.; Gil, C.; et al. Dolomite flora of the Baetic Ranges glades (South Spain). Flora-Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 359–375. [Google Scholar] [CrossRef]
- Allison, J.R.; Stevens, T.E. Vascular flora of Ketona dolomite outcrops in Bibb County, Alabama. Castanea 2001, 66, 154–205. [Google Scholar]
- Mota, J.F.; Valle, F.; Cabello, J. Dolomitic vegetation of South Spain. Plant Ecol. 1993, 109, 29–45. [Google Scholar] [CrossRef]
- Lu, M.; Du, H.; Song, T.; Peng, W.; Su, L.; Zhang, H.; Zeng, Z.; Wang, K.; Tang, W.; Wei, L.; et al. Drivers of tree survival in an evergreen-deciduous broadleaf karst forest in southwest China. For. Ecol. Manag. 2021, 499, 119598. [Google Scholar] [CrossRef]
- Batalha, M.A.; Pipenbaher, N.; Bakan, B.; Kaligaric, M.; Skornik, S. Assessing community assembly along a successional gradient in the North Adriatic Karst with functional and phylogenetic distances. Oecologia 2015, 178, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zou, X.; Lu, E.; Deng, Y.; Luo, Y.; Chen, H.; Liu, W. Litterfall biomass and nutrient cycling in karst and nearby non-karst forests in tropical China: A 10-year comparison. Sci. Total Environ. 2021, 758, 143619. [Google Scholar] [CrossRef]
- Yi, R.; Xu, X.; Zhu, S.; Zhang, Y.; Zhong, F.; Zeng, X.; Xu, C. Difference in hydraulic resistance between planted forest and naturally regenerated forest and its implications for ecosystem restoration in subtropical karst landscapes. J. Hydrol. 2021, 596, 126093. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Xiong, K.; Yu, Y.; Min, X. Changes of leaf functional traits in karst rocky desertification ecological environment and the driving factors. Glob. Ecol. Conserv. 2020, 24, e1381. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.; Wang, K.; Yang, H.; Yue, Y.; Zeng, Z.; Peng, W.; Zeng, F. Influences of stand characteristics and environmental factors on forest biomass and root–shoot allocation in southwest China. Ecol. Eng. 2016, 91, 7–15. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Xu, X.; Jiang, X.; Ding, B.; Wu, P.; Ding, F. Plant Functional Trait Responses to Dolomite and Limestone Karst Forests in Southwest China. Forests 2022, 13, 2187. https://doi.org/10.3390/f13122187
Zhou H, Xu X, Jiang X, Ding B, Wu P, Ding F. Plant Functional Trait Responses to Dolomite and Limestone Karst Forests in Southwest China. Forests. 2022; 13(12):2187. https://doi.org/10.3390/f13122187
Chicago/Turabian StyleZhou, Hua, Xuelei Xu, Xia Jiang, Bo Ding, Peng Wu, and Fangjun Ding. 2022. "Plant Functional Trait Responses to Dolomite and Limestone Karst Forests in Southwest China" Forests 13, no. 12: 2187. https://doi.org/10.3390/f13122187
APA StyleZhou, H., Xu, X., Jiang, X., Ding, B., Wu, P., & Ding, F. (2022). Plant Functional Trait Responses to Dolomite and Limestone Karst Forests in Southwest China. Forests, 13(12), 2187. https://doi.org/10.3390/f13122187