
Low Frequency of Plants Associated with Symbiotic Nitrogen-Fixers Exhibits High Frequency of Free-Living Nitrogen Fixing Bacteria: A Study in Karst Shrub Ecosystems of Southwest China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Survey of Plant and Collecting of Soil Sample
2.3. Extraction and Purification of Soil DNA
2.4. Amplification of Polymerase Chain Reaction (PCR) and Analyses of Terminal Restriction Fragment Length Polymorphism (T-RFLP)
2.5. Abundance of nifH Genes
2.6. Determination of Soil Physicochemical Parameters
2.7. Analyses of Statistics
3. Results
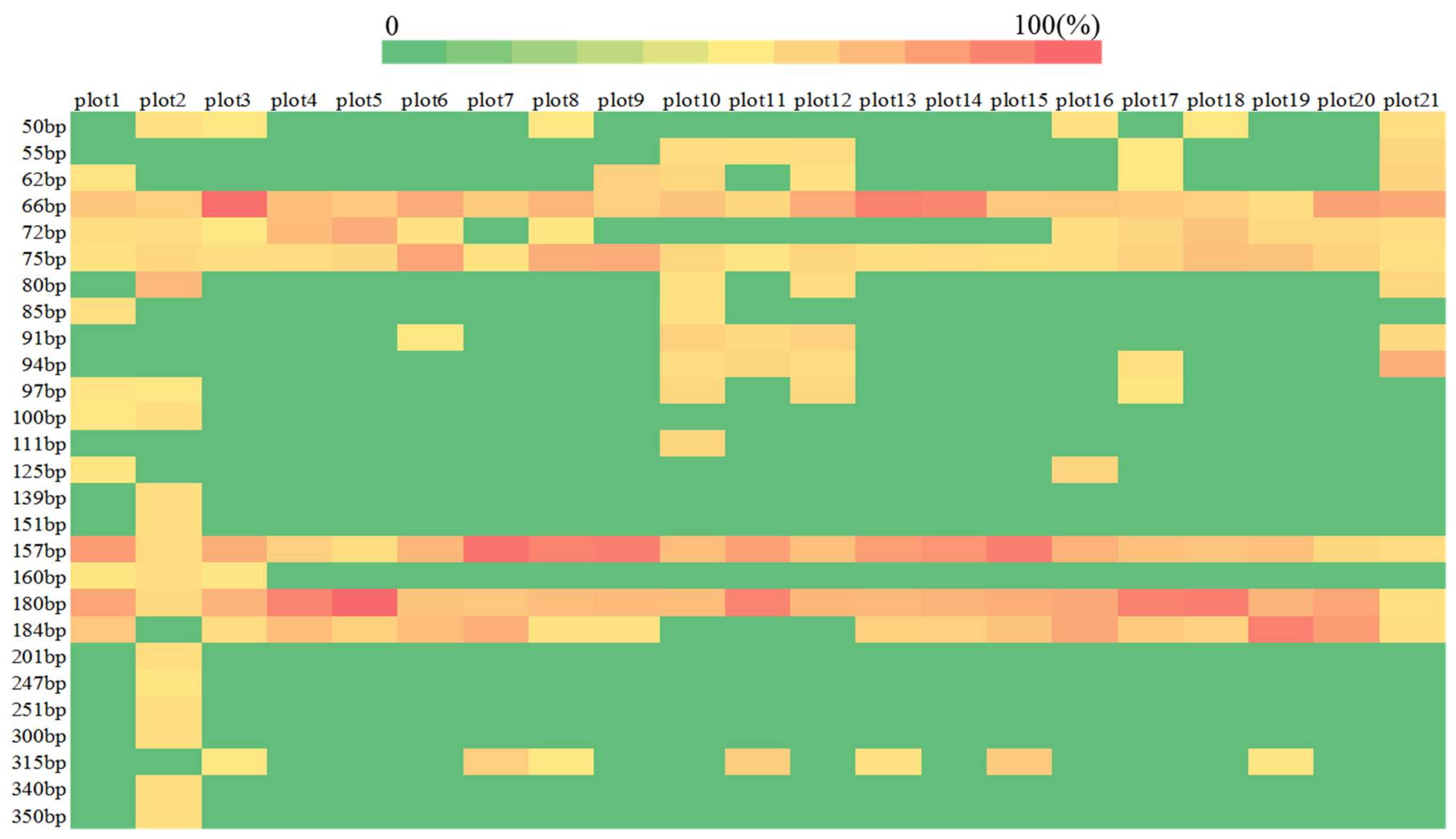
3.1. Composition Community of Soil Free-Living Nitrogen-Fixing Bacteria from Different Plot Soils in Karst Shrub Ecosystems
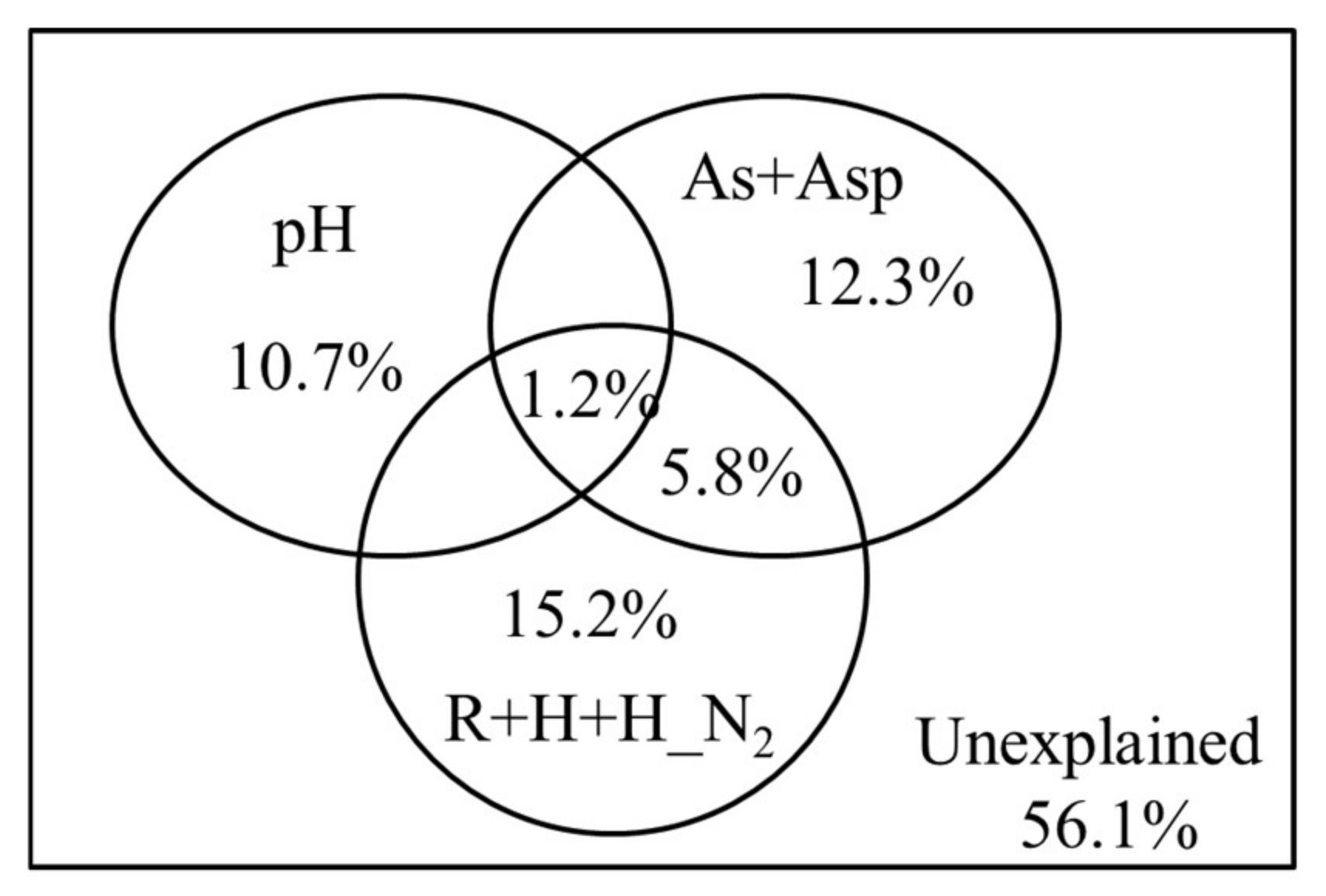
3.2. Factors Influencing Soil Free-Living Nitrogen-Fixing Bacterial Community Structure in Karst Shrub Ecosystems
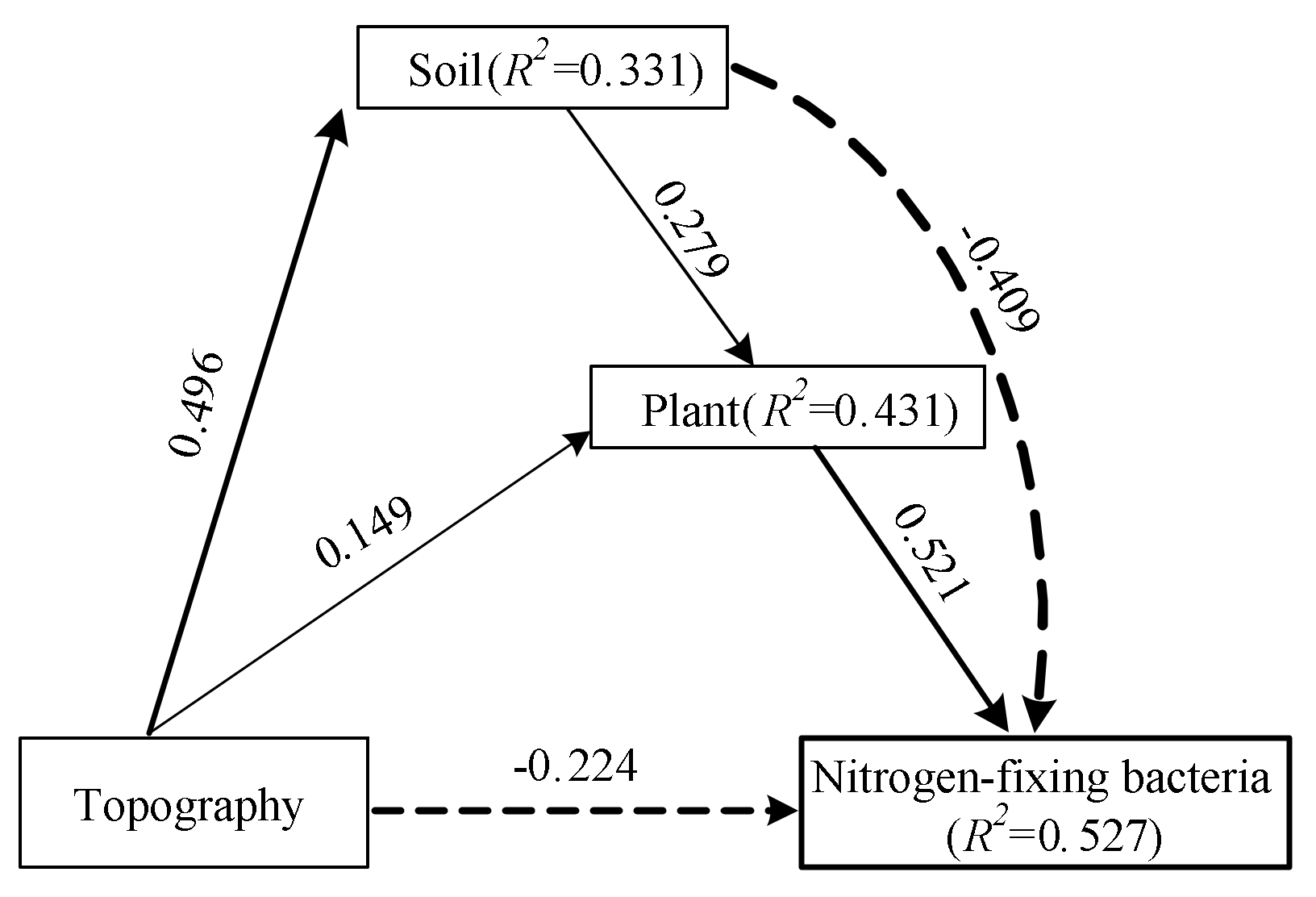
3.3. Factors Influencing the Community Abundance of Free-Living Nitrogen-Fixing Bacteria in the Karst Shrub Ecosystems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liu, S.J.; Li, D.J.; Wang, K.L. Leaf N: P stoichiometry across plant functional groups in the karst region of southwestern China. Trees Struct. Funct. 2015, 29, 883–892. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.J.; Li, D.J.; Chen, H.S.; Wang, K.L. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Ladha, J.K. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil 1995, 174, 3–28. [Google Scholar] [CrossRef]
- Shridhar, B.S. Review: Nitrogen fixing microorganisms. Int. J. Microbiol. Res. 2012, 3, 46–52. [Google Scholar]
- Burgmann, H.; Widmer, F.; Von Sigler, W.; Zeyer, J. New molecular screening tools for analysis of free-living diazotrophs in soil. Appl. Environ. Microb. 2004, 70, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Kahindi, J.H.P.; Woomer, P.; George, T.; de Souza Moreira, F.M.; Karanja, N.K.; Giller, K.E. Agricultural intensification, soil biodiversity and ecosystem function in the tropics: The role of nitrogen-fixing bacteria. Appl. Soil Ecol. 1997, 6, 55–76. [Google Scholar] [CrossRef]
- Lovell, C.R.; Piceno, Y.M.; Quattro, J.M.; Bagwell, C.E. Molecular analysis of diazotroph diversity in the rhizosphere of the smooth cordgrass, Spartina alterniflora. Appl. Environ. Microbiol. 2000, 66, 3814–3822. [Google Scholar] [CrossRef] [Green Version]
- Poly, F.; Ranjard, L.; Nazaret, S.; Goirbiere, F.; Monrozier, L.J. Comparison of nifH gene pools in soils and soil microenvironments with contrasting properties. Appl. Environ. Microbiol. 2001, 67, 2255–2262. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; He, X.Y.; Wang, K.L.; Xie, Y.J.; Xie, Q.; O’Donnell, A.G.; Chen, C.Y. The Bradyrhizobium-legume symbiosis is dominant in the shrubby ecosystem of the Karst region, Southwest China. Eur. J. Soil Biol. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Lan, A.J.; Zhang, B.P.; Xiong, K.N.; An, Y.L. Spatial pattern of the fragile karst environment in southwest Guizhou province. Geogr. Res. 2003, 22, 733–741. (In Chinese) [Google Scholar]
- Tu, Y.L. A analysis of flora and ecological characteristics of karst scrubs in Guizhou province. J. Guizhou Norm. Univ. (Nat. Sci.) 1995, 13, 1–8. (In Chinese) [Google Scholar]
- Pan, F.J.; Liang, Y.M.; Zhang, W.; Zhao, J.; Wang, K.L. Enhanced nitrogen availability in karst ecosystems by oxalic acid release in the rhizosphere. Front. Plant Sci. 2016, 7, 687. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.M.; Pan, F.J.; He, X.Y.; Chen, X.B.; Su, Y.R. Effect of vegetation types on soil arbuscular mycorrhizal fungi and nitrogen-fixing bacterial communities in a karst region. Environ. Sci. Pollut. R. 2016, 23, 18482–18491. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Yannarell, A.C.; Mackie, R.I.; Davis, S.C.; Mackie, R.I. Changes in N-transforming archaea and bacteria in soil during the establishment of bioenergy crops. PLoS ONE 2011, 6, e24750. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.R.R.; Marriel, I.; Jenkins, S.N.; Lanyon, C.; Seldin, L.; O’Donnell, A.G. Molecular detection and quantification of nifH gene sequences in the rhizosphere of sorghum (Sorghum bicolor) sown with two levels of nitrogen fertilizer. Appl. Soil Ecol. 2009, 42, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Tai, X.S.; Mao, W.L.; Liu, G.X.; Chen, T.; Zhang, W.; Wu, X.K.; Long, H.Z.; Zhang, B.G.; Zhang, Y. High diversity of nitrogen-fixing bacteria in the upper reaches of the Heihe River, northwestern China. Biogeosciences 2013, 10, 5589–5600. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.G.; Li, D.Q.; Wang, H.M.; Xiao, Q.M.; Liu, X.D. Molecular diversity of nitrogen-fixing bacteria from the Tibetan Plateau, China. FEMS Microbiol. Lett. 2006, 260, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelso, D.R.; Mele, P.M. The impact of crop residue amendments and limes on microbial community structure and nitrogen-fixing bacteria in the wheat rhizosphere. Aust. J. Soil Res. 2006, 44, 319–329. [Google Scholar] [CrossRef]
- Collavino, M.M.; Tripp, H.J.; Frank, I.E.; Vidoz, M.L.; Calderoli, P.A.; Donato, M.; Zehr, J.P.; Aguilar, O.M. NifH pyrosequencing reveals the potential for location-specific soil chemistry to influence N2-fixing community dynamics. Environ. Microb. 2014, 16, 3211–3223. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, C.; Kou, Y.; Wang, J.; Tu, B.; Li, H.; Li, X.; Wang, C.; Yao, M. Soil pH is a major driver of soil diazotrophic community assembly in Qinghai-Tibet alpine meadows. Soil Biol. Biochem. 2017, 115, 547–555. [Google Scholar] [CrossRef]
- Wang, Y.S.; Li, C.N.; Shen, Z.H.; Rui, J.P.; Jin, D.C.; Li, J.B.; Li, X.Z. Community assemblage of free-living diazotrophs along the elevational gradient of Mount Gongga. Soil Ecol. Lett. 2019, 1, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Chen, H.S.; Wang, K.L.; Hou, Y.; Zhang, J.G. Spatial variability of soil organic carbon and available phosphorus in a typical Karst depression, northwest of Guangxi. Acta Ecol. Sin. 2007, 27, 5168–5175. (In Chinese) [Google Scholar]
- He, X.Y.; Wang, K.L.; Zhang, W.; Chen, Z.H.; Zhu, Y.G.; Chen, H.S. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst. Plant Soil 2008, 307, 123–134. [Google Scholar] [CrossRef]
- Ma, K.P.; Huang, J.H.; Yu, S.L.; Chen, L.Z. Plant community diversity in Dongling Mountain Beijing, China, species richness, evenness and species diversity. Acta Ecol. Sin. 1995, 15, 268–277. (In Chinese) [Google Scholar]
- Hsu, S.F.; Buckley, D.H. Evidence for the functional significance of diazotroph community structure in soil. ISME J. 2008, 3, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Lukow, T.; Dunfield, P.F.; Liesack, W. Use of the T-RFLP technique to assess spatial and temporal changes in the bacterial community structure with in an agricultural soil planted with transgenic and no transgenic potato plants. FEMS Microbial. Ecol. 2000, 32, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Aldrich-Wolfe, L. Distinct mycorrhizal communities on new and established hosts in a transitional tropical plant community. Ecology 2007, 88, 559. [Google Scholar] [CrossRef] [PubMed]
- Colwell, J.D. The estimation of phosphorus fertilizer requirements of wheat in southern New South Wales by soil analysis. Aust. J. Exp. Agric. Anim. Husb. 1963, 3, 190–197. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Ed.; American Society of Agronomy, Inc.: Madison, WI, USA, 1965; Volume 2, pp. 1149–1178. [Google Scholar]
- Xiao, D.; Tan, Y.J.; Liu, X.; Yang, R.; Zhang, W.; He, X.Y.; Xu, Z.H.; Wang, K.L. Responses of soil diazotrophs to legume species and density in a karst grassland, southwest China. Agric. Ecosyst. Environ. 2020, 288, 106707. [Google Scholar] [CrossRef]
- Koponen, P.; Nygren, P.; Domenach, A.M.; Le Roux, C.; Saur, E.; Roggy, J.C. Nodulation and dinitrogen fixation of legume trees in a tropical freshwater swamp forest in French Guiana. J. Trop. Ecol. 2003, 19, 655–666. [Google Scholar] [CrossRef]
- Pereira e Silva, M.C.; Schloter-Hai, B.; Schloter, M.; van Elsas, J.D.; Salles, J.F. Temporal Dynamics of Abundance and Composition of Nitrogen-Fixing Communities across Agricultural Soils. PLoS ONE 2013, 8, e74500. [Google Scholar] [CrossRef]
- Legay, N.; Baxendale, C.; Grigulis, K.; Krainer, U.; Kastl, E.; Schloter, M.; Bardgett, R.; Arnoldi, C.; Bahn, M.; Dumont, M. Contribution of above-and below-ground plant traits to the structure and function of grassland soil microbial communities. Ann. Bot. 2014, 114, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, M.G.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization. Soil Biol. Biochem. 2016, 98, 109–126. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y.; Biryukova, O.; Kuznetzova, T.; Molter, K. Carbon partitioning in plant soil, carbon dioxide fluxes and enzyme activities as affected by cutting ryegrass. Biol. Fertil. Soils 2002, 35, 348–358. [Google Scholar]
- Atere, C.T.; Ge, T.D.; Zhu, Z.K.; Tong, C.L.; Jones, D.L.; Shibistova, O.; Guggenberger, G.; Wu, J.S. Rice rhizodeposition and carbon stabilization in paddy soil are regulated via dry-rewetting cycles and nitrogen fertilization. Biol. Fert. Soils 2017, 53, 407–417. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Domanski, G. Carbon input by plant into the soil. Review. Z. Pflanzenernahr. Bodenkd. 2000, 163, 421–431. [Google Scholar] [CrossRef]
- Wang, M.; Shi, S.; Lin, F.; Hao, Z.Q.; Jiang, P.; Dai, G.H. Effects of soil water and nitrogen on growth and photosynthetic response of Manchurian Ash (Fraxinus mandshurica) seedlings in Northeastern China. PLoS ONE 2012, 7, e30754. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Hu, Y.G.; Ren, C.Z.; Guo, L.C.; Wang, C.L.; Jiang, Y.; Wang, X.J.; Phendukani, H.; Zeng, Z.H. Effects of nitrogen application on chlorophyll fluorescence parameter and leaf gas exchange in naked oat. J. Integ. Agric. 2013, 12, 2164–2171. [Google Scholar] [CrossRef] [Green Version]
- Reardon, C.L.; Gollany, H.T.; Wuest, S.B. Diazotroph community structure and abundance in wheat–fallow and wheat–pea crop rotations. Soil Biol. Biochem. 2014, 69, 406–412. [Google Scholar] [CrossRef]
- Osler, G.H.R.; Sommerkorn, M. Toward a complete soil C and N cycle, incorporating the soil fauna. Ecology 2007, 88, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, S.; Kravchenko, A.N.; Robertson, G.P. Topography influences management system effects on total soil carbon and nitrogen. Soil Sci. Soc. Am. J. 2009, 73, 2059–2067. [Google Scholar] [CrossRef]
- Wiaux, F.; Cornelis, J.T.; Cao, W.; Vanclooster, M.; Van Oost, K. Combined effect of geomorphic and pedogenic processes on the distribution of soil organic carbon quality along an eroding hillslope on loess soil. Geoderma 2014, 216, 36–47. [Google Scholar] [CrossRef]
- Feng, S.Z.; Su, Y.R.; Zhang, W.; Chen, X.B.; He, X.Y. Effects of slope position and soil horizon on soil microbial biomass and abundance in karst primary forest of Southwest China. Environ. Sci. 2015, 36, 3832–3838. (In Chinese) [Google Scholar]
- Gotsch, S. Land cover and slope position affect water use and microclimate in the tropical montane cloud forests of Central Veracruz, Mexico. In Proceedings of the New Frontiers in Tropical Biology: The Next 50 Years (A Joint Meeting of ATBC and OTS), San Jose, Costa Rica, 23–27 June 2013. [Google Scholar]
- Brewer, S.W.; Rejmanek, M.; Webb, M.A.H.; Fine, P.V.A. Relationships of phytogeography and diversity of tropical tree species with limestone topography in Southern Belize. J. Biogeogr. 2003, 30, 1669–1688. [Google Scholar] [CrossRef]
- Burke, A. Classification and ordination of plant communities of the Naukluft Mountain, Namibia. J. Veg. Sci. 2001, 12, 53–60. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Hu, G.; Ni, J. Effects of topographical and edaphic factors on the distribution of plant communities in two subtropical karst forests, southwestern China. J. Mt. Sci. 2013, 10, 95–104. [Google Scholar] [CrossRef]
- Göransson, H.; Edwards, P.J.; Perreijn, K.; Smittenberg, R.H.; Venterink, H.O. Rocks create nitrogen hotspots and N: P heterogeneity by funnelling rain. Biogeochemistry 2014, 121, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Jean, M.E.; Phalyvong, K.; Forest-Drolet, J.; Belleng, J.P. Molybdenum and phosphorus limitation of asymbiotic nitrogen fixation in forests of Eastern Canada: Influence of vegetative cover and seasonal variability. Soil Biol. Biochem. 2013, 67, 140–146. [Google Scholar] [CrossRef]
- Li, Y.; Jia, Z.J.; Sun, Q.Y.; Zhan, J.; Yang, Y.; Wang, D. Ecological restoration alter microbial communities in mine tailings profiles. Sci. Rep. 2016, 6, 25193. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Dominant Species | Plants Associated with Symbiotic Nitrogen-Fixers | Richness of all Plant | Shannon Diversity ofall Plant | Richness of N2-Fixing Plant | Shannon Diversity of N2-Fixing Plant |
|---|---|---|---|---|---|---|
| Plot1 | Croton lachnocarpus, Mahonia fortunei, Ficus tinctoria | Bauhinia hypochrysa, Dalbergia hancei | 42 | 4.69 | 2 | 0.6 |
| Plot2 | Loropetalum chinense, Cyclobalanopsis glauca, Croton lachnocarpus | Bauhinia acuminata, Bauhinia hypochrysa, Pterolobium punctatum, Albizia odoratissima, Derris fordii | 56 | 4.91 | 5 | 0.67 |
| Plot3 | Sinosideroxylon pedunculatum, Derris fordii, Chukrasia tabularis | Bauhinia hypochrysa, Pterolobium punctatum, Albizia odoratissima, Derris fordii | 50 | 4.95 | 4 | 0.5 |
| Plot4 | Litsea coreana, Pyrus calleryana, Syzygium championii | - | 38 | 4.11 | 0 | 0 |
| Plot5 | Viburnum triplinerve, Loropetalum chinense, Pyrus calleryana | Campylotropis delavayi, Sophora tonkinensis | 31 | 4.51 | 2 | 0.11 |
| Plot6 | Pyracantha fortuneana, Rosa laevigata, Mallotus repandus | Bauhinia championii, Indigofera atropurpurea | 53 | 5.00 | 2 | 0.15 |
| Plot7 | Pterolobium punctatum, Alchornea trewioides, Xylosma controversum | Bauhinia hypochrysa, Pterolobium punctatum | 68 | 5.52 | 2 | 0.56 |
| Plot8 | Bauhinia hypochrysa, Millettia pachycarpa, Ficus tinctoria | Millettia pachycarpa, Bauhinia hypochrysa, Pterolobium punctatum | 61 | 5.20 | 3 | 0.73 |
| Plot9 | Chaydaia rubrinervis, Sinoadina racemosa, Xylosma controversum | Bauhinia hypochrysa, Pterolobium punctatum | 61 | 5.45 | 2 | 0.42 |
| Plot10 | Litsea coreana, Pittosporum tonkinense, Syzygium championii | Millettia pachycarpa, Bauhinia championii, Albizia odoratissima | 48 | 4.40 | 3 | 0.12 |
| Plot11 | Loropetalum chinense, Pittosporum tonkinense, Syzygium championii | Indigofera atropurpurea | 34 | 4.43 | 1 | 0.13 |
| Plot12 | Loropetalum chinense, Litsea coreana, Syzygium championii | Campylotropis delavayi, Albizia odoratissima | 40 | 4.78 | 2 | 0.14 |
| Plot13 | Syzygium championii, Pyracantha fortuneana, Viburnum triplinerve | Millettia eurybotrya, Campylotropis delavayi | 49 | 5.21 | 2 | 0.09 |
| Plot14 | Pittosporum tonkinense, Syzygium championii, Litsea coreana | Bauhinia championii, Indigofera atropurpurea, Campylotropis delavayi | 50 | 4.99 | 3 | 0.18 |
| Plot15 | Litsea coreana, Pittosporum tonkinense, Itea chinensis | Bauhinia championii | 35 | 4.47 | 1 | 0.22 |
| Plot16 | Loropetalum chinense, Pyracantha fortuneana, Pyrus calleryana | Albizia odoratissima, Gelsemium elegans | 41 | 4.74 | 2 | 0.21 |
| Plot17 | Loropetalum chinense, Litsea coreana, Pyrus calleryana | Dendrolobium triangulare, | 40 | 4.36 | 0 | 0 |
| Plot18 | Loropetalum chinense, Viburnum fordiae, Viburnum triplinerve | - | 47 | 4.57 | 0 | 0 |
| Plot19 | Alangium chinense, Pyracantha fortuneana, Alchornea trewioides | Dendrolobium triangulare, Gelsemium elegans, Bauhinia championii | 61 | 5.09 | 3 | 0.28 |
| Plot20 | Loropetalum chinense, Pyrus calleryana, Pyracantha fortuneana | Gelsemium elegans, Campylotropis delavayi | 36 | 4.46 | 2 | 0.25 |
| Plot21 | Loropetalum chinense, Pyracantha fortuneana, Pistacia weinmannifolia | Pterolobium punctatum | 34 | 4.34 | 1 | 0.06 |
| Plot | Olsen-P (mg·kg−1) | pH | Total Nitrogen (g·kg−1) | Soil Organic Carbon (g·kg−1) | C/N | NH4+-N (mg·kg−1) | NO3-N (mg·kg−1) | nifH Gene Abundance |
|---|---|---|---|---|---|---|---|---|
| Plot1 | 7.16 | 7.6 | 12.53 | 120.7 | 9.63 | 34.23 | 4.55 | 3.80 × 106 |
| Plot2 | 5.43 | 7.32 | 4.71 | 57.1 | 12.11 | 17.25 | 5.63 | 4.78 × 106 |
| Plot3 | 5.59 | 7.38 | 8.77 | 87.42 | 9.97 | 29.53 | 4.87 | 4.07 × 106 |
| Plot4 | 3.48 | 7.14 | 5.35 | 55.89 | 10.45 | 18.56 | 5.57 | 1.61 × 106 |
| Plot5 | 5.34 | 7.44 | 4.12 | 65.16 | 15.81 | 23.24 | 4.98 | 5.29 × 105 |
| Plot6 | 5.3 | 7.81 | 4.32 | 63.09 | 14.59 | 22.68 | 4.69 | 6.65 × 105 |
| Plot7 | 4.78 | 7.92 | 4.46 | 58.73 | 13.17 | 24.84 | 6.45 | 4.04 × 106 |
| Plot8 | 5.43 | 7.32 | 6.76 | 78.21 | 11.56 | 27.47 | 4.32 | 3.89 × 106 |
| Plot9 | 4.02 | 7.88 | 3.99 | 57.54 | 14.42 | 19.54 | 5.86 | 1.51 × 106 |
| Plot10 | 4.56 | 7.59 | 5.78 | 66.78 | 11.55 | 23.46 | 5.02 | 5.34 × 106 |
| Plot11 | 6.44 | 7.8 | 5.56 | 67.94 | 12.23 | 23.52 | 5.45 | 2.27 × 106 |
| Plot12 | 5.46 | 7.82 | 6.26 | 64.75 | 10.34 | 24.48 | 5.12 | 5.27 × 106 |
| Plot13 | 6.57 | 7.98 | 4.93 | 71.45 | 14.5 | 24.89 | 4.86 | 1.32 × 106 |
| Plot14 | 6.75 | 7.87 | 6.25 | 72.86 | 11.66 | 25.35 | 4.68 | 2.98 × 106 |
| Plot15 | 6.24 | 7.83 | 6.35 | 75.53 | 11.89 | 26.46 | 4.56 | 2.42 × 106 |
| Plot16 | 5.22 | 7.77 | 4.35 | 65.67 | 15.09 | 25.68 | 5.36 | 1.19 × 107 |
| Plot17 | 3.74 | 7.27 | 3.76 | 56.5 | 15.01 | 18.56 | 5368 | 2.87 × 107 |
| Plot18 | 2.79 | 6.52 | 2.54 | 45.37 | 17.84 | 46.36 | 5.87 | 5.01 × 107 |
| Plot19 | 4.73 | 7.78 | 4.1 | 57.35 | 14 | 24.87 | 5.96 | 5.36 × 105 |
| Plot20 | 3.71 | 7.95 | 3.16 | 50.09 | 15.86 | 16.78 | 6.54 | 4.98 × 106 |
| Plot21 | 4.16 | 8.03 | 4.51 | 69.21 | 15.36 | 25.45 | 5.67 | 4.69 × 105 |
| nifH Abundance | Shannon Diversity Index of Plants Associated with Symbiotic N-Fixers | Richness Index of Plants Associated with Symbiotic N-Fixers | Shannon Diversity Index of all Plants | Richness Index of all Plants | C/N | SOC | TN | pH | AP | NO3−-N | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NH4+-N | 0.574 ** | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| NO3−-N | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | - |
| AP | −0.516 * | ns | ns | ns | ns | −0.534 * | 0.727 ** | 0.662 ** | ns | - | |
| pH | −0.719 * | ns | ns | ns | ns | ns | ns | ns | - | ||
| TN | ns | 0.468 * | ns | ns | ns | 0.796 ** | 0.686 ** | - | |||
| SOC | ns | ns | ns | ns | ns | −0.61 ** | - | ||||
| C/N | 0.475 * | ns | ns | ns | ns | - | |||||
| Richness index of all plants | ns | 0.602 * | 0.502 * | 0.857 ** | - | ||||||
| Shannon diversity index of all plants | ns | 0.601 * | 0.509 * | - | |||||||
| Richness index of plants associated with symbiotic N-fixers | −0.443 * | 0.676 * | - | ||||||||
| Shannon diversity index of plants associated with symbiotic N-fixers | ns | - | |||||||||
| nifH abundance | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; He, X.; Chen, X.; Su, Y.; Pan, F.; Hu, L. Low Frequency of Plants Associated with Symbiotic Nitrogen-Fixers Exhibits High Frequency of Free-Living Nitrogen Fixing Bacteria: A Study in Karst Shrub Ecosystems of Southwest China. Forests 2022, 13, 163. https://doi.org/10.3390/f13020163
Liang Y, He X, Chen X, Su Y, Pan F, Hu L. Low Frequency of Plants Associated with Symbiotic Nitrogen-Fixers Exhibits High Frequency of Free-Living Nitrogen Fixing Bacteria: A Study in Karst Shrub Ecosystems of Southwest China. Forests. 2022; 13(2):163. https://doi.org/10.3390/f13020163
Chicago/Turabian StyleLiang, Yueming, Xunyang He, Xiangbi Chen, Yirong Su, Fujing Pan, and Lening Hu. 2022. "Low Frequency of Plants Associated with Symbiotic Nitrogen-Fixers Exhibits High Frequency of Free-Living Nitrogen Fixing Bacteria: A Study in Karst Shrub Ecosystems of Southwest China" Forests 13, no. 2: 163. https://doi.org/10.3390/f13020163
APA StyleLiang, Y., He, X., Chen, X., Su, Y., Pan, F., & Hu, L. (2022). Low Frequency of Plants Associated with Symbiotic Nitrogen-Fixers Exhibits High Frequency of Free-Living Nitrogen Fixing Bacteria: A Study in Karst Shrub Ecosystems of Southwest China. Forests, 13(2), 163. https://doi.org/10.3390/f13020163