
Origin, Persistence, and Vulnerability to Climate Changes of Podocarpus Populations in Central African Mountains

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nuclear Microsatellites Development
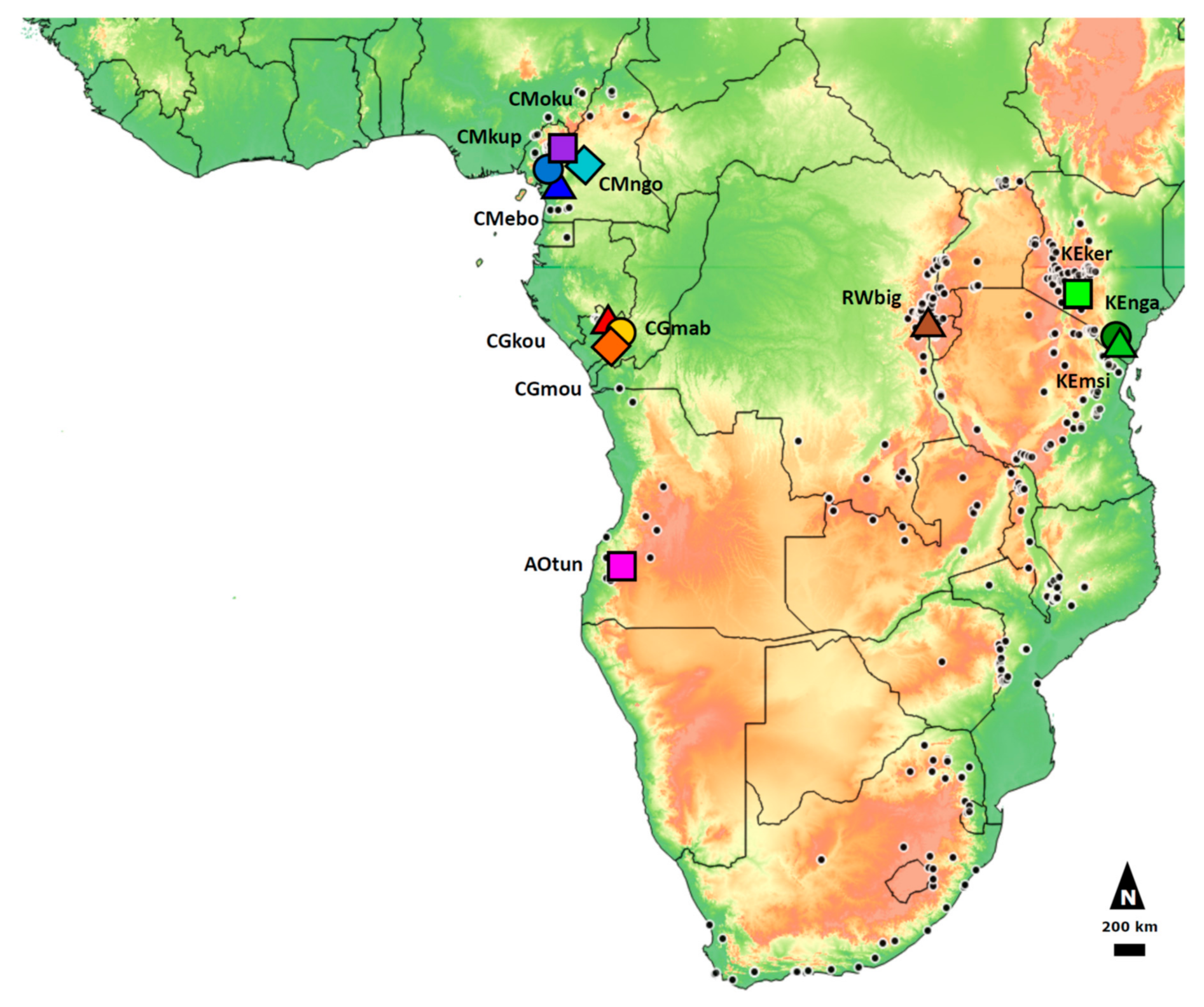
2.2. Population Sampling and Nuclear Microsatellites Genotyping
2.3. Genetic Diversity and Fine-Scale Genetic Structure within Populations
2.4. Genetic Structure
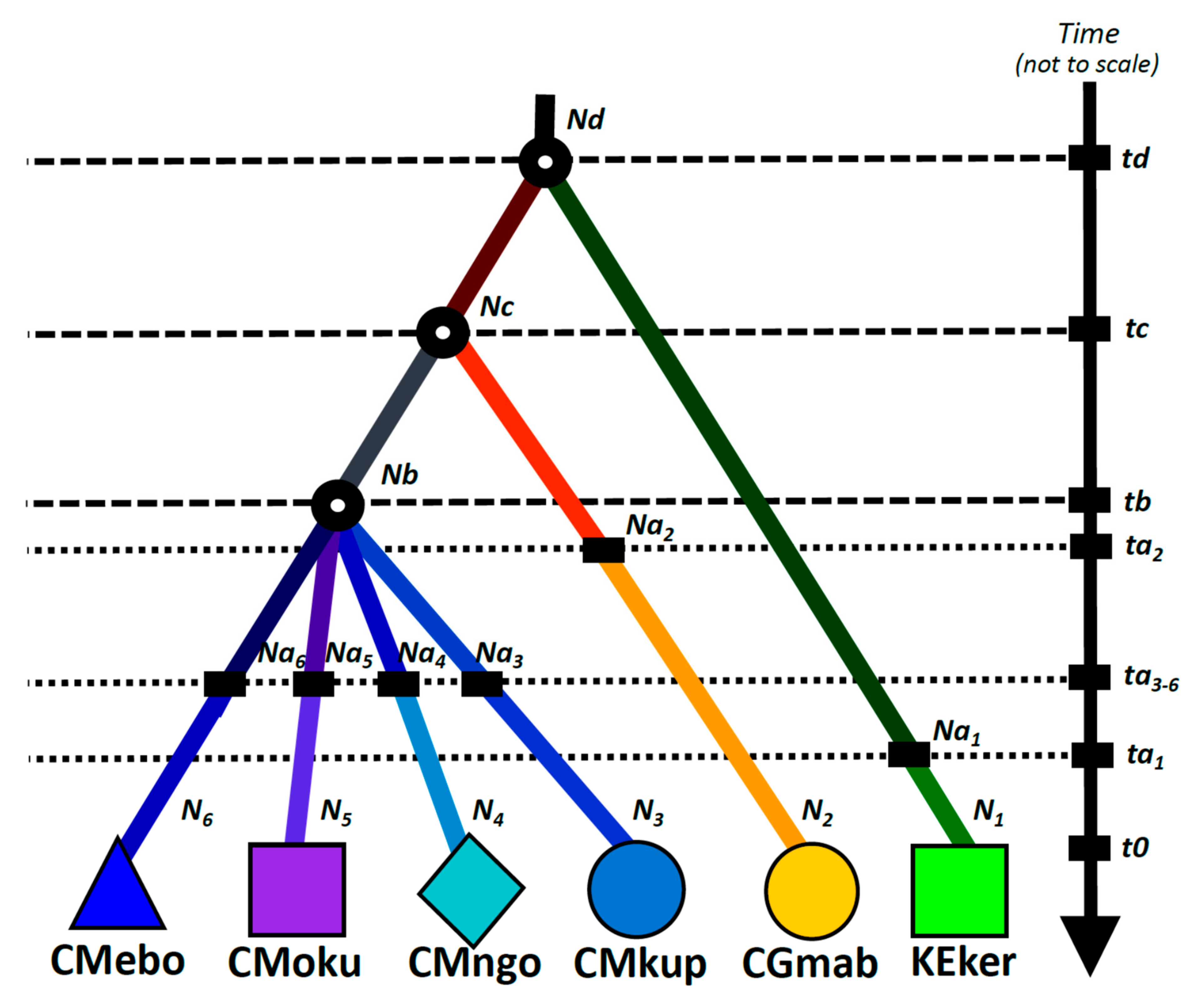
2.5. Demographic Inference
3. Results
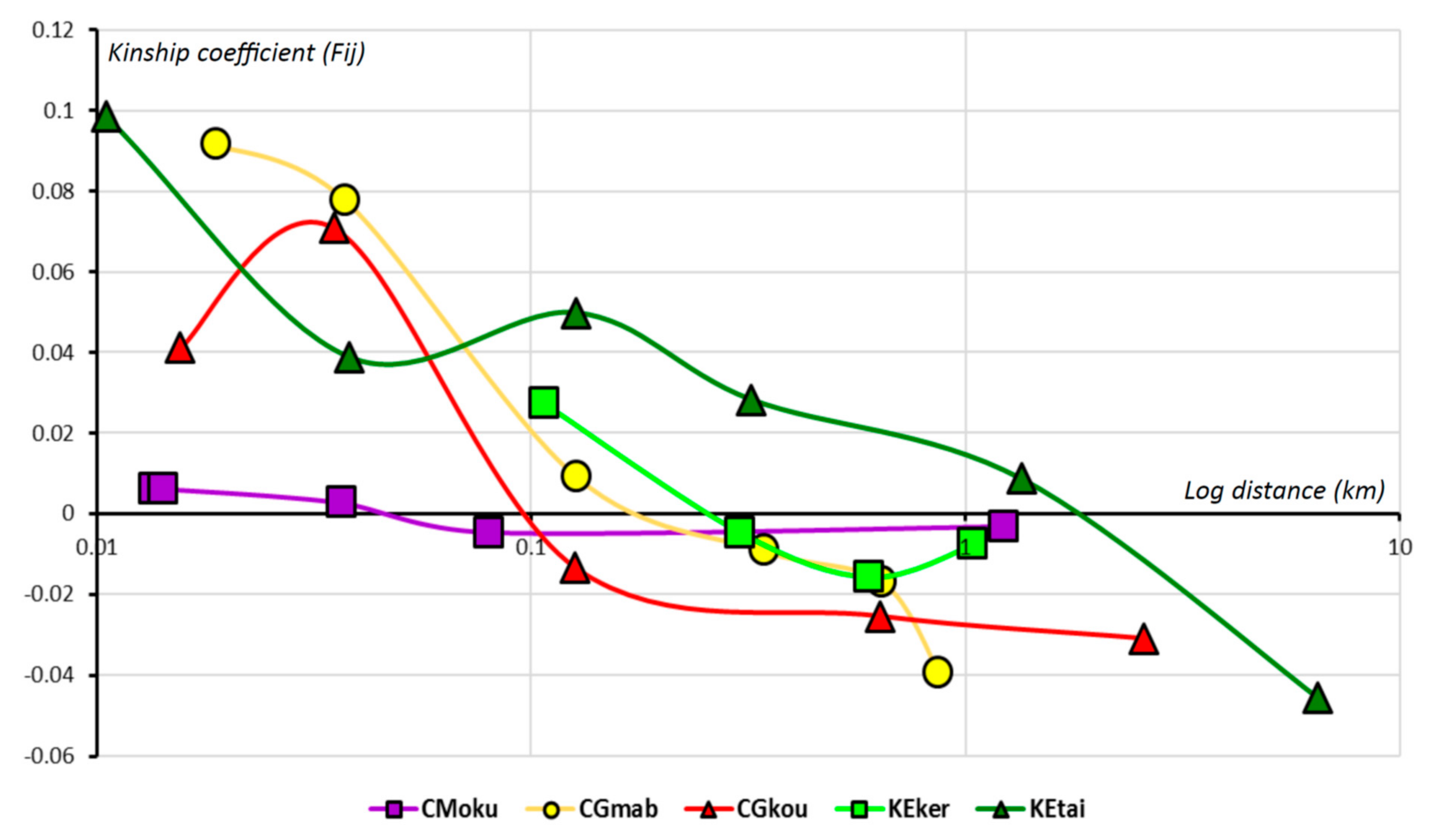
3.1. Genetic Diversity and Fine-Scale Genetic Structure within Populations
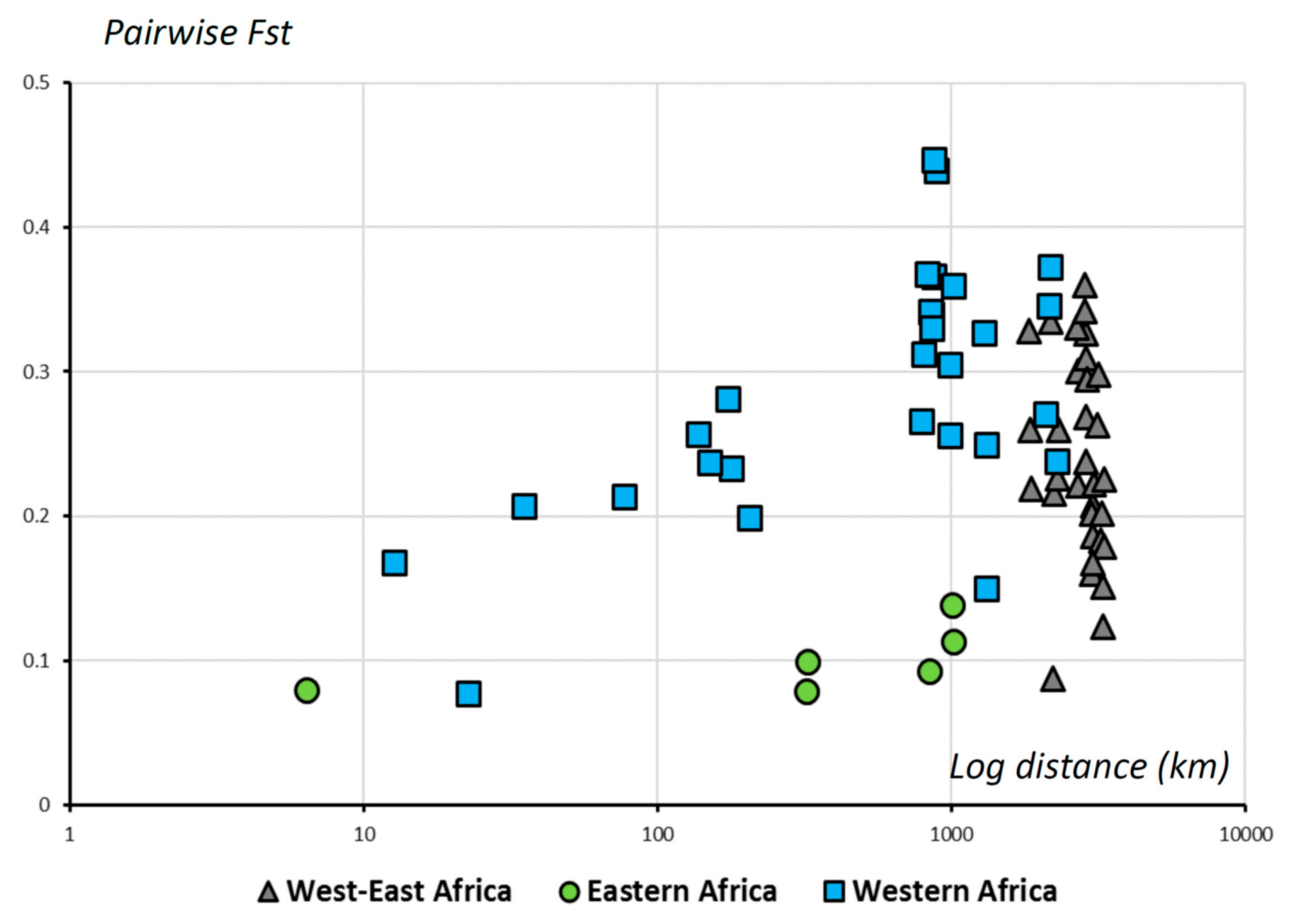
3.2. Genetic Structure
3.3. Demographic Inference
4. Discussion
4.1. Wide Distribution despite Currently Limited Dispersal Capacities
4.2. Recent and Strong Bottleneck in the West despite Past Connections throughout Africa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, F. The Afromontane Region. In Biogeography and Ecology of Southern Africa; Springer: Dordrecht, The Netherlands, 1978; pp. 463–513. ISBN 978-94-009-9953-4. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Küper, W.; Sommer, J.H.; Lovett, J.C.; Mutke, J.; Linder, H.P.; Beentje, H.J.; Rompaey, R.S.A.R.V.; Chatelain, C.; Sosef, M.; Barthlott, W. Africa’s Hotspots of Biodiversity Redefined. Ann. Mo. Bot. Gard. 2004, 91, 525–535. [Google Scholar]
- Dagallier, L.-P.M.J.; Janssens, S.B.; Dauby, G.; Blach-Overgaard, A.; Mackinder, B.A.; Droissart, V.; Svenning, J.-C.; Sosef, M.S.M.; Stévart, T.; Harris, D.J.; et al. Cradles and museums of generic plant diversity across tropical Africa. New Phytol. 2020, 225, 2196–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, F. The history of the Afromontane archipelago and the scientific need for its conservation. Afr. J. Ecol. 1981, 19, 33–54. [Google Scholar] [CrossRef]
- Platts, P.J.; Gereau, R.E.; Burgess, N.D.; Marchant, R. Spatial heterogeneity of climate change in an Afromontane centre of endemism. Ecography 2013, 36, 518–530. [Google Scholar] [CrossRef]
- Adie, H.; Lawes, M. Podocarps in Africa: Temperate zone relicts or rainforest survivors? Smithson. Contrib. Bot. 2011, 95, 79–100. [Google Scholar] [CrossRef] [Green Version]
- Dupont, L.M.; Jahns, S.; Marret, F.; Ning, S. Vegetation change in equatorial West Africa: Time-slices for the last 150 ka. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 155, 95–122. [Google Scholar] [CrossRef]
- Lézine, A.-M.; Assi-Kaudjhis, C.; Roche, E.; Vincens, A.; Achoundong, G. Towards an understanding of West African montane forest response to climate change. J. Biogeogr. 2013, 40, 183–196. [Google Scholar] [CrossRef]
- Lézine, A.-M.; Izumi, K.; Kageyama, M.; Achoundong, G. A 90,000-year record of Afromontane forest responses to climate change. Science 2019, 363, 177–181. [Google Scholar] [CrossRef]
- Osmaston, H.A.; Harrison, S.P. The Late Quaternary glaciation of Africa: A regional synthesis. Quat. Int. 2005, 138–139, 32–54. [Google Scholar] [CrossRef]
- Finch, J.; Leng, M.J.; Marchant, R. Late Quaternary vegetation dynamics in a biodiversity hotspot, the Uluguru Mountains of Tanzania. Quat. Res. 2009, 72, 111–122. [Google Scholar] [CrossRef]
- Izumi, K.; Lézine, A.-M. Pollen-based biome reconstructions over the past 18,000 years and atmospheric CO2 impacts on vegetation in equatorial mountains of Africa. Quat. Sci. Rev. 2016, 152, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Elenga, H.; Schwartz, D.; Vincens, A. Pollen evidence of late Quaternary vegetation and inferred climate changes in Congo. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 109, 345–356. [Google Scholar] [CrossRef]
- Vincens, A.; Schwartz, D.; Elenga, H.; Reynaud-Farrera, I.; Alexandre, A.; Bertaux, J.; Mariotti, A.; Martin, L.; Meunier, J.-D.; Nguetsop, F.; et al. Forest response to climate changes in Atlantic Equatorial Africa during the last 4000 years BP and inheritance on the modern landscapes. J. Biogeogr. 1999, 26, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Migliore, J.; Lézine, A.-M.; Hardy, O.J. The recent colonization history of the most widespread Podocarpus tree species in Afromontane forests. Ann. Bot. 2020, 126, 73–83. [Google Scholar] [CrossRef]
- Adie, H.; Lawes, M.J. Role reversal in the stand dynamics of an angiosperm–conifer forest: Colonising angiosperms precede a shade-tolerant conifer in Afrotemperate forest. For. Ecol. Manag. 2009, 258, 159–168. [Google Scholar] [CrossRef]
- Hart, L.A.; Grieve, G.R.H.; Downs, C.T. Fruiting phenology and implications of fruit availability in the fragmented Ngele forest complex, KwaZulu-Natal, South Africa. S. Afr. J. Bot. 2013, 88, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Uriarte, M.; Anciães, M.; Da Silva, M.T.; Rubim, P.; Johnson, E.; Bruna, E.M. Disentangling the drivers of reduced long-distance seed dispersal by birds in an experimentally fragmented landscape. Ecology 2011, 92, 924–937. [Google Scholar] [CrossRef]
- Maley, J.; Caballé, G.; Sita, P. Etude d’un peuplement résiduel à basse altitude de Podocarpus latifolius sur le flanc congolais du massif du Chaillu. Implications paléoclimatiques et biogéographiques. Etude de la pluie pollinique actuelle. Paysages Quat. L’afrique Cent. Atl. 1990, 336–352. [Google Scholar]
- Verlhac, L.; Izumi, K.; Lézine, A.-M.; Lemonnier, K.; Buchet, G.; Achoundong, G.; Tchiengué, B. Altitudinal distribution of pollen, plants and biomes in the Cameroon highlands. Rev. Palaeobot. Palynol. 2018, 259, 21–28. [Google Scholar] [CrossRef]
- Vekemans, X.; Hardy, O.J. New insights from fine-scale spatial genetic structure analyses in plant populations. Mol. Ecol. 2004, 13, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Demenou, B.B.; Piñeiro, R.; Hardy, O.J. Origin and history of the Dahomey Gap separating West and Central African rain forests: Insights from the phylogeography of the legume tree Distemonanthus benthamianus. J. Biogeogr. 2016, 43, 1020–1031. [Google Scholar] [CrossRef]
- Demenou, B.B.; Doucet, J.-L.; Hardy, O.J. History of the fragmentation of the African rain forest in the Dahomey Gap: Insight from the demographic history of Terminalia superba. Heredity 2018, 120, 547. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, R.; Dauby, G.; Kaymak, E.; Hardy, O.J. Pleistocene population expansions of shade-tolerant trees indicate fragmentation of the African rainforest during the Ice Ages. Proc. R. Soc. B 2017, 284, 20171800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Meglécz, E.; Pech, N.; Gilles, A.; Dubut, V.; Hingamp, P.; Trilles, A.; Grenier, R.; Martin, J.-F. QDD version 3.1: A user friendly computer program for microsatellite selection and primer design revisited: Experimental validation of variables determining genotyping success rate. Mol. Ecol. Resour. 2014, 14, 1302–1313. [Google Scholar] [CrossRef]
- Micheneau, C.; Dauby, G.; Bourland, N.; Doucet, J.-L.; Hardy, O.J. Development and characterization of microsatellite loci in Pericopsis elata (Fabaceae) using a cost-efficient approach. Am. J. Bot. 2011, 98, e268–e270. [Google Scholar] [CrossRef]
- Holleley, C.E.; Geerts, P.G. Multiplex Manager 1.0: A cross-platform computer program that plans and optimizes multiplex PCR. Bio Tech. 2009, 46, 511–517. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Hardy, O.J.; Charbonnel, N.; Fréville, H.; Heuertz, M. Microsatellite Allele Sizes: A Simple Test to Assess Their Significance on Genetic Differentiation. Genetics 2003, 163, 1467–1482. [Google Scholar] [CrossRef]
- Hardy, O.J.; Vekemans, X. SPAGeDi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, J.-M.; Santos, F.; Beaumont, M.A.; Robert, C.P.; Marin, J.-M.; Balding, D.J.; Guillemaud, T.; Estoup, A. Inferring population history with DIY ABC: A user-friendly approach to approximate Bayesian computation. Bioinformatics 2008, 24, 2713–2719. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, J.-M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.-M.; Estoup, A. DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef] [Green Version]
- Vincens, A. Palynologie, Environnements Actuels et Plio-Pléistocènes à l’est du lac Turkana (Kenya). Ph.D. Thesis, Université d’Aix-Marseille, Marseille, France, 1982. [Google Scholar]
- Gajewski, K.; Lézine, A.-M.; Vincens, A.; Delestan, A.; Sawada, M. Modern climate–vegetation–pollen relations in Africa and adjacent areas. Quat. Sci. Rev. 2002, 21, 1611–1631. [Google Scholar] [CrossRef]
- Hadley, A.S.; Betts, M.G. The effects of landscape fragmentation on pollination dynamics: Absence of evidence not evidence of absence. Biol. Rev. 2012, 87, 526–544. [Google Scholar] [CrossRef]
- Bizoux, J.-P.; Daïnou, K.; Bourland, N.; Hardy, O.J.; Heuertz, M.; Mahy, G.; Doucet, J.-L. Spatial genetic structure in Milicia excelsa (Moraceae) indicates extensive gene dispersal in a low-density wind-pollinated tropical tree. Mol. Ecol. 2009, 18, 4398–4408. [Google Scholar] [CrossRef]
- Debout, G.D.G.; Doucet, J.-L.; Hardy, O.J. Population history and gene dispersal inferred from spatial genetic structure of a Central African timber tree, Distemonanthus benthamianus (Caesalpinioideae). Heredity 2011, 106, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duminil, J.; Brown, R.P.; Ewédjè, E.-E.B.; Mardulyn, P.; Doucet, J.-L.; Hardy, O.J. Large-scale pattern of genetic differentiation within African rainforest trees: Insights on the roles of ecological gradients and past climate changes on the evolution of Erythrophleum spp (Fabaceae). BMC Evol. Biol. 2013, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gichira, A.W.; Li, Z.-Z.; Saina, J.K.; Hu, G.-W.; Gituru, R.W.; Wang, Q.-F.; Chen, J.-M. Demographic history and population genetic structure of Hagenia abyssinica (Rosaceae), a tropical tree endemic to the Ethiopian highlands and eastern African mountains. Tree Genet. Genomes 2017, 13, 72. [Google Scholar] [CrossRef]
- Kadu, C.A.C.; Konrad, H.; Schueler, S.; Muluvi, G.M.; Eyog-Matig, O.; Muchugi, A.; Williams, V.L.; Ramamonjisoa, L.; Kapinga, C.; Foahom, B.; et al. Divergent pattern of nuclear genetic diversity across the range of the Afromontane Prunus africana mirrors variable climate of African highlands. Ann. Bot. 2013, 111, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Kormann, U.; Scherber, C.; Tscharntke, T.; Klein, N.; Larbig, M.; Valente, J.J.; Hadley, A.S.; Betts, M.G. Corridors restore animal-mediated pollination in fragmented tropical forest landscapes. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152347. [Google Scholar] [CrossRef] [Green Version]
- Kadu, C.A.C.; Schueler, S.; Konrad, H.; Muluvi, G.M.M.; Eyog-Matig, O.; Muchugi, A.; Williams, V.L.; Ramamonjisoa, L.; Kapinga, C.; Foahom, B.; et al. Phylogeography of the Afromontane Prunus africana reveals a former migration corridor between East and West African highlands. Mol. Ecol. 2011, 20, 165–178. [Google Scholar] [CrossRef]
- Chartier, M.; Dressler, S.; Schönenberger, J.; Mora, A.R.; Sarthou, C.; Wang, W.; Jabbour, F. The evolution of afro-montane Delphinium (Ranunculaceae): Morphospecies, phylogenetics and biogeography. Taxon 2016, 65, 1313–1327. [Google Scholar] [CrossRef]
- Mairal, M.; Namaganda, M.; Gizaw, A.; Chala, D.; Brochmann, C.; Catalán, P. Multiple mountain-hopping colonization of sky-islands on the two sides of Tropical Africa during the Pleistocene: The afroalpine Festuca grasses. J. Biogeogr. 2021, 48, 1858–1874. [Google Scholar] [CrossRef]
- Dupont, L.M.; Donner, B.; Schneider, R.; Wefer, G. Mid-Pleistocene environmental change in tropical Africa began as early as 1.05 Ma. Geology 2001, 29, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Measey, G.J.; Tolley, K.A. Sequential Fragmentation of Pleistocene Forests in an East Africa Biodiversity Hotspot: Chameleons as a Model to Track Forest History. PLoS ONE 2011, 6, e26606. [Google Scholar] [CrossRef] [Green Version]
- Voje, K.L.; Hemp, C.; Flagstad, Ø.; Sætre, G.-P.; Stenseth, N.C. Climatic change as an engine for speciation in flightless Orthoptera species inhabiting African mountains. Mol. Ecol. 2009, 18, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Huhndorf, M.H.; Peterhans, J.C.K.; Loew, S.S. Comparative phylogeography of three endemic rodents from the Albertine Rift, east central Africa. Mol. Ecol. 2007, 16, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Bryja, J.; Mikula, O.; Patzenhauerová, H.; Oguge, N.O.; Šumbera, R.; Verheyen, E. The role of dispersal and vicariance in the Pleistocene history of an East African mountain rodent, Praomys delectorum. J. Biogeogr. 2014, 41, 196–208. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J.; Hackett, S.J.; Bates, J.M.; Crowe, T.M. Coalescent models reveal the relative roles of ancestral polymorphism, vicariance, and dispersal in shaping phylogeographical structure of an African montane forest robin. Mol. Phylogenetics Evol. 2006, 38, 171–188. [Google Scholar] [CrossRef]
- Bouiges, A.; Yassin, A.; Ikogou, M.; Lelarge, C.; Sikoa, A.-R.; Mona, S.; Veuille, M. Detecting recent changes in the demographic parameters of drosophilid populations from western and central Africa. Comptes Rendus Geosci. 2013, 345, 297–305. [Google Scholar] [CrossRef]
- Lézine, A.-M.; Izumi, K.; Achoundong, G. Mbi Crater (Cameroon) illustrates the relations between mountain and lowland forests over the past 15,000 years in western equatorial Africa. Quat. Int. 2020, in press. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G.S. Range Shifts and Adaptative Responses to Quaternary Climate Change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Ayele, T.B.; Gailing, O.; Finkeldey, R. Assessment and integration of genetic, morphological and demographic variation in Hagenia abyssinica (Bruce) J.F. Gmel to guide its conservation. J. Nat. Conserv. 2011, 19, 8–17. [Google Scholar] [CrossRef]
- Kahindo, C.; Bowie, R.C.K.; Bates, J.M. The relevance of data on genetic diversity for the conservation of Afro-montane regions. Biol. Conserv. 2007, 134, 262–270. [Google Scholar] [CrossRef]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrerias-Diego, Y.; Lobo, J. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and methodological approaches. Mol. Ecol. 2008, 17, 5177–5188. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | n | Missing Genotypes | NA | AR | Ho | He | Fi | Pval |
|---|---|---|---|---|---|---|---|---|
| AOtun | 9 | 1.4 (16.0%) | 3.56 | 2.72 | 0.210 | 0.560 | 0.650 | 0 |
| CGkou | 28 | 1.8 (6.3%) | 4.33 | 2.69 | 0.417 | 0.582 | 0.288 | 0 |
| CGmab | 33 | 2.6 (7.7%) | 3.22 | 2.25 | 0.302 | 0.483 | 0.381 | 0 |
| CGmou | 8 | 0.6 (6.9%) | 2.22 | 1.87 | 0.306 | 0.336 | 0.097 | 0.396 |
| CMebo | 23 | 0.7 (2.9%) | 4.11 | 2.71 | 0.560 | 0.613 | 0.089 | 0.047 |
| CMkup | 12 | 0.9 (7.4%) | 3.22 | 2.55 | 0.546 | 0.570 | 0.047 | 0.543 |
| CMngo | 16 | 0.2 (1.4%) | 2.67 | 2.12 | 0.480 | 0.458 | −0.050 | 0.473 |
| CMoku | 37 | 0.6 (1.5%) | 5.89 | 3.02 | 0.516 | 0.606 | 0.150 | 0 |
| KEker | 32 | 0.6 (1.7%) | 8.33 | 3.55 | 0.454 | 0.705 | 0.359 | 0 |
| KEmsi | 16 | 1.7 (10.4%) | 5.22 | 3.21 | 0.445 | 0.690 | 0.365 | 0 |
| KEnga | 22 | 1.3 (6.1%) | 7.00 | 3.49 | 0.415 | 0.707 | 0.420 | 0 |
| RWbig | 11 | 1.1 (10.1%) | 5.44 | 3.33 | 0.439 | 0.651 | 0.339 | 0 |
| All populations | 247 | 13.3 (5.4%) | 12.56 | 3.80 | 0.443 | 0.768 | 0.423 | 0 |
| Populations | AOtun | CGkou | CGmab | CGmou | CMebo | CMkup | CMngo | CMoku | KEker | KEmsi | KEnga | RWbig |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AOtun | 0.18 | 0.15 | 0.17 | 0.33 | 0.26 | 0.39 | 0.29 | 0.15 | 0.16 | 0.21 | 0.07 | |
| CGkou | 0.16 | 0.03 | 0.14 | 0.47 | 0.37 | 0.42 | 0.32 | 0.21 | 0.32 | 0.32 | 0.32 | |
| CGmab | 0.25 | 0.16 | 0.05 | 0.43 | 0.38 | 0.43 | 0.32 | 0.16 | 0.31 | 0.30 | 0.28 | |
| CGmou | 0.33 | 0.19 | 0.08 | 0.41 | 0.40 | 0.48 | 0.27 | 0.14 | 0.31 | 0.29 | 0.32 | |
| CMebo | 0.25 | 0.25 | 0.30 | 0.35 | 0.23 | 0.28 | 0.19 | 0.19 | 0.21 | 0.27 | 0.29 | |
| CMkup | 0.34 | 0.34 | 0.37 | 0.44 | 0.21 | 0.19 | 0.23 | 0.21 | 0.19 | 0.29 | 0.15 | |
| CMngo | 0.37 | 0.33 | 0.33 | 0.45 | 0.25 | 0.28 | 0.22 | 0.26 | 0.26 | 0.27 | 0.25 | |
| CMoku | 0.24 | 0.25 | 0.30 | 0.36 | 0.19 | 0.23 | 0.24 | 0.10 | 0.15 | 0.18 | 0.28 | |
| KEker | 0.16 | 0.23 | 0.30 | 0.33 | 0.20 | 0.22 | 0.29 | 0.17 | 0.05 | 0.10 | 0.14 | |
| KEmsi | 0.21 | 0.27 | 0.31 | 0.34 | 0.18 | 0.18 | 0.26 | 0.15 | 0.10 | 0.00 | 0.11 | |
| KEnga | 0.19 | 0.24 | 0.33 | 0.36 | 0.20 | 0.23 | 0.30 | 0.12 | 0.08 | 0.08 | 0.16 | |
| RWbig | 0.09 | 0.23 | 0.26 | 0.33 | 0.21 | 0.26 | 0.33 | 0.23 | 0.09 | 0.14 | 0.11 | |
| Average: | FST = 0.24 | RST = 0.25 | ||||||||||
| Parameter | Prior Distribution | Mean | Mode | q025 | q975 | |
|---|---|---|---|---|---|---|
| N1 | Effective population size KEker at t0 | Uniform (10–50,000) | 37,700 | 46,600 | 16,200 | 49,600 |
| N2 | Effective population size CGmab at t0 | Uniform (10–20,000) | 2440 | 960 | 325 | 11,900 |
| N3 | Effective population size CMkup at t0 | Uniform (10–2000) | 1010 | 793 | 257 | 1900 |
| N4 | Effective population size CMngo at t0 | Uniform (10–2000) | 737 | 476 | 150 | 1760 |
| N5 | Effective population size CMoku at t0 | Uniform (10–20,000) | 8440 | 4130 | 1810 | 19,000 |
| N6 | Effective population size CMebo at t0 | Uniform (10–2000) | 1210 | 1140 | 372 | 1940 |
| Na1 | Effective population size KEker at ta1 | Uniform (10–100,000) | 51,800 | 34,800 | 14,400 | 96,600 |
| Na2 | Effective population size CGmab at ta3–6 | Uniform (10–100,000) | 53,700 | 96,300 | 7740 | 98,200 |
| Na3 | Effective population size CMkup at ta3–6 | Uniform (10–100,000) | 54,900 | 81,400 | 6560 | 98,000 |
| Na4 | Effective population size CMngo at ta3–6 | Uniform (10–100,000) | 57,700 | 93,300 | 6730 | 98,500 |
| Na5 | Effective population size CMoku at ta3–6 | Uniform (10–100,000) | 31,500 | 8950 | 2690 | 92,300 |
| Na6 | Effective population size CMebo at ta3–6 | Uniform (10–100,000) | 47,700 | 20,200 | 4970 | 96,600 |
| Nb | Effective population size CM at tb | Uniform (10–100,000) | 42,500 | 22,700 | 4860 | 94,900 |
| Nc | Effective population size CM+CG at tc | Uniform (10–100,000) | 53,200 | 47,200 | 7590 | 97,700 |
| Nd | Effective population size CM+CG+KE at td | Uniform (10–100,000) | 47,000 | 29,500 | 9130 | 93,000 |
| ta1 | Last demographic changes for KEker | Uniform (500–250,000) | 91,500 | 14,750 | 3350 | 237,000 |
| ta2 | Last demographic changes for CGmab | Uniform (500–250,000) | 151,500 | 232,000 | 17,350 | 246,000 |
| ta3–6 | Last demographic changes for CM | Uniform (500–250,000) | 32,650 | 31,800 | 8800 | 67,000 |
| tb | Divergence CM pops | Uniform (500–1,000,000) | 311,000 | 250,500 | 75,000 | 680,000 |
| tc | Divergence CG pop | Uniform (500–1,000,000) | 560,000 | 499,000 | 231,000 | 895,000 |
| td | Divergence KE pop | Uniform (500–1,000,000) | 845,000 | 980,000 | 525,000 | 995,000 |
| Mean µ | Mean mutation rate | Uniform (1.0 × 10−5–1.0 × 10−4) | 4.00 × 10−5 | 2.56 × 10−5 | 1.28 × 10−5 | 8.59 × 10−5 |
| Mean P | Mean coefficient P | Uniform (0.1–0.3) | 0.226 | 0.300 | 0.122 | 0.300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliore, J.; Lézine, A.-M.; Veuille, M.; Achoundong, G.; Tchiengué, B.; Boom, A.F.; Monthe, F.K.; Bouka, G.U.D.; Omondi, S.F.; Wagura, L.; et al. Origin, Persistence, and Vulnerability to Climate Changes of Podocarpus Populations in Central African Mountains. Forests 2022, 13, 208. https://doi.org/10.3390/f13020208
Migliore J, Lézine A-M, Veuille M, Achoundong G, Tchiengué B, Boom AF, Monthe FK, Bouka GUD, Omondi SF, Wagura L, et al. Origin, Persistence, and Vulnerability to Climate Changes of Podocarpus Populations in Central African Mountains. Forests. 2022; 13(2):208. https://doi.org/10.3390/f13020208
Chicago/Turabian StyleMigliore, Jérémy, Anne-Marie Lézine, Michel Veuille, Gaston Achoundong, Barthélémy Tchiengué, Arthur F. Boom, Franck K. Monthe, Gaël U. D. Bouka, Stephen F. Omondi, Lawrence Wagura, and et al. 2022. "Origin, Persistence, and Vulnerability to Climate Changes of Podocarpus Populations in Central African Mountains" Forests 13, no. 2: 208. https://doi.org/10.3390/f13020208
APA StyleMigliore, J., Lézine, A. -M., Veuille, M., Achoundong, G., Tchiengué, B., Boom, A. F., Monthe, F. K., Bouka, G. U. D., Omondi, S. F., Wagura, L., Gonçalves, F. M. P., Stévart, T., Farminhão, J. N. M., & Hardy, O. J. (2022). Origin, Persistence, and Vulnerability to Climate Changes of Podocarpus Populations in Central African Mountains. Forests, 13(2), 208. https://doi.org/10.3390/f13020208