Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Preventing Damage Mechanisms for Desert Plants under Drought Stress
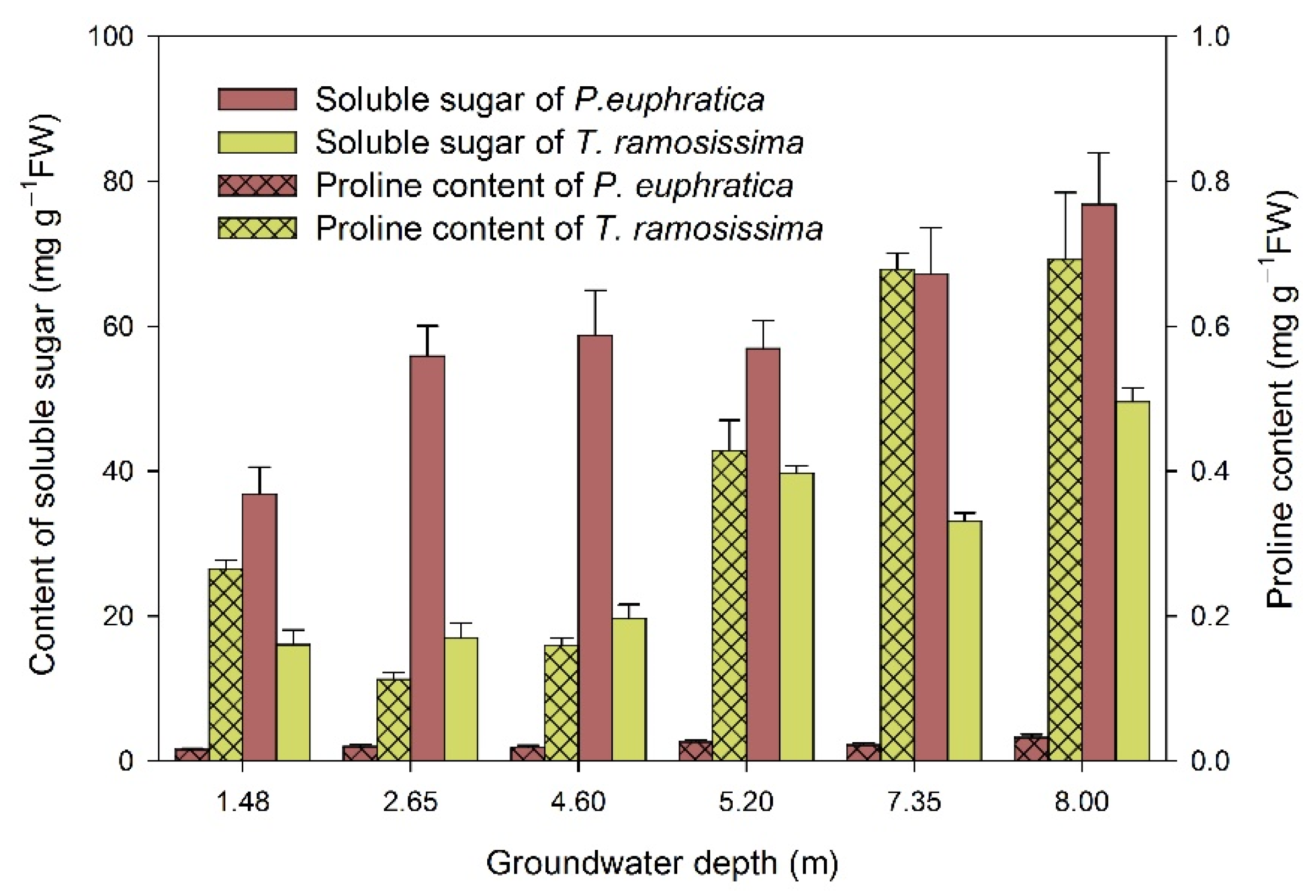
2.1. Plant Physiological Metabolic Processes under Drought Stress
2.2. Plant Damage-Avoidance Mechanisms under Drought Stress
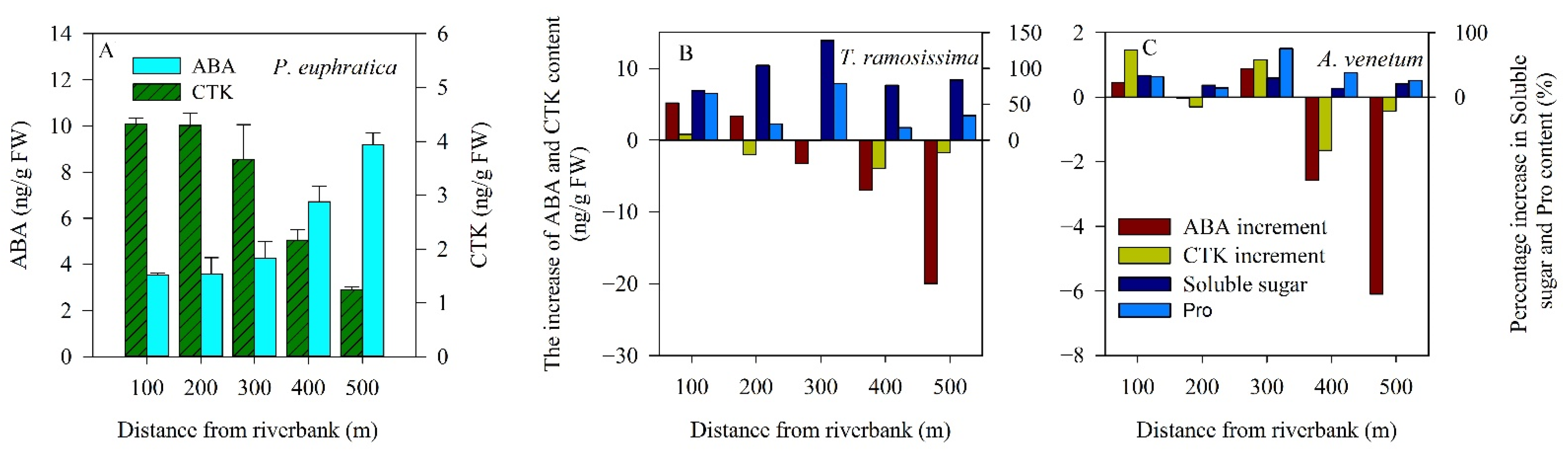
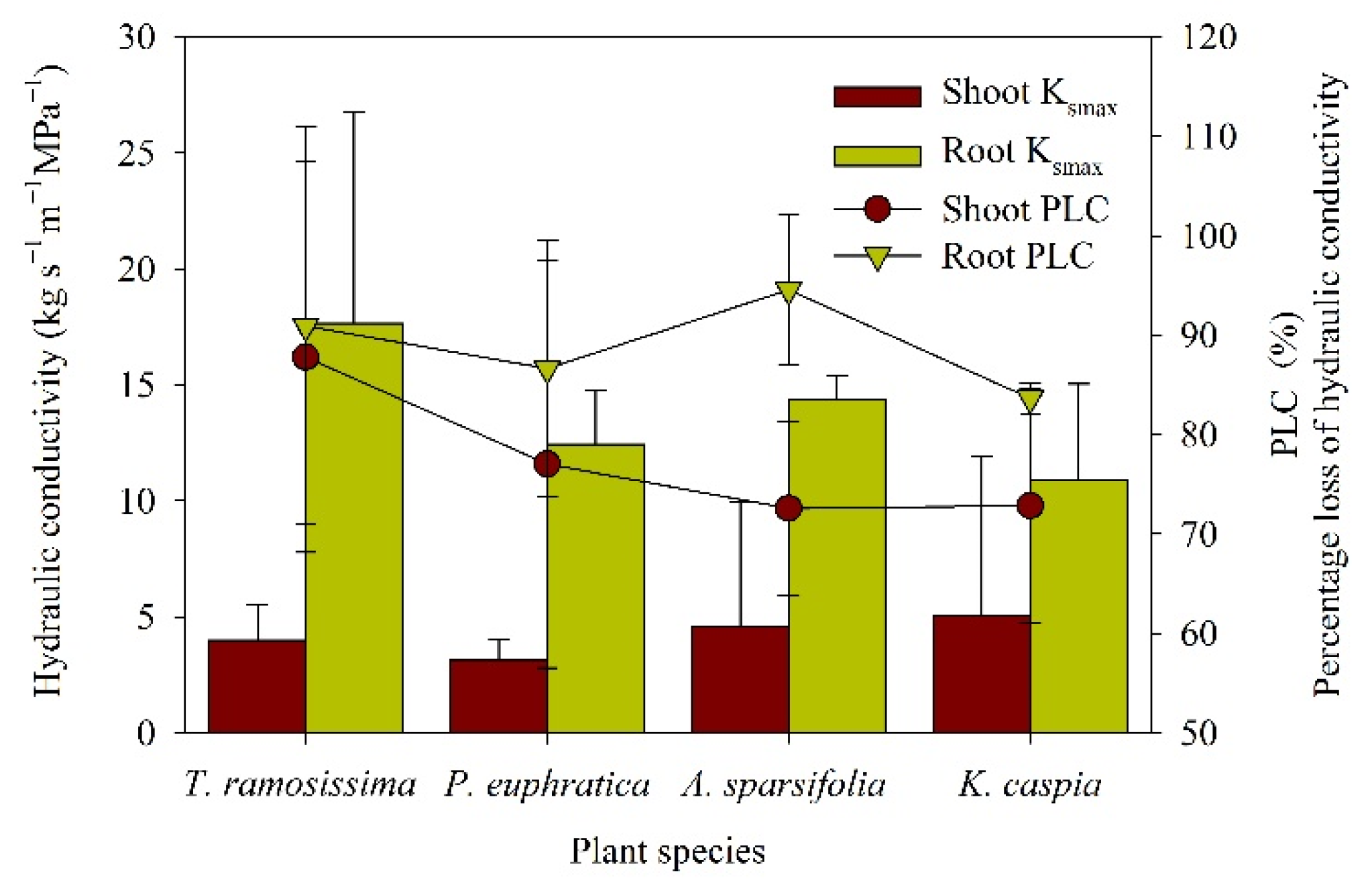
2.3. Drought-Resistance Mechanism of Plants under Drought Stress
3. Water Use and Regulation Strategies of Desert Riparian Plants
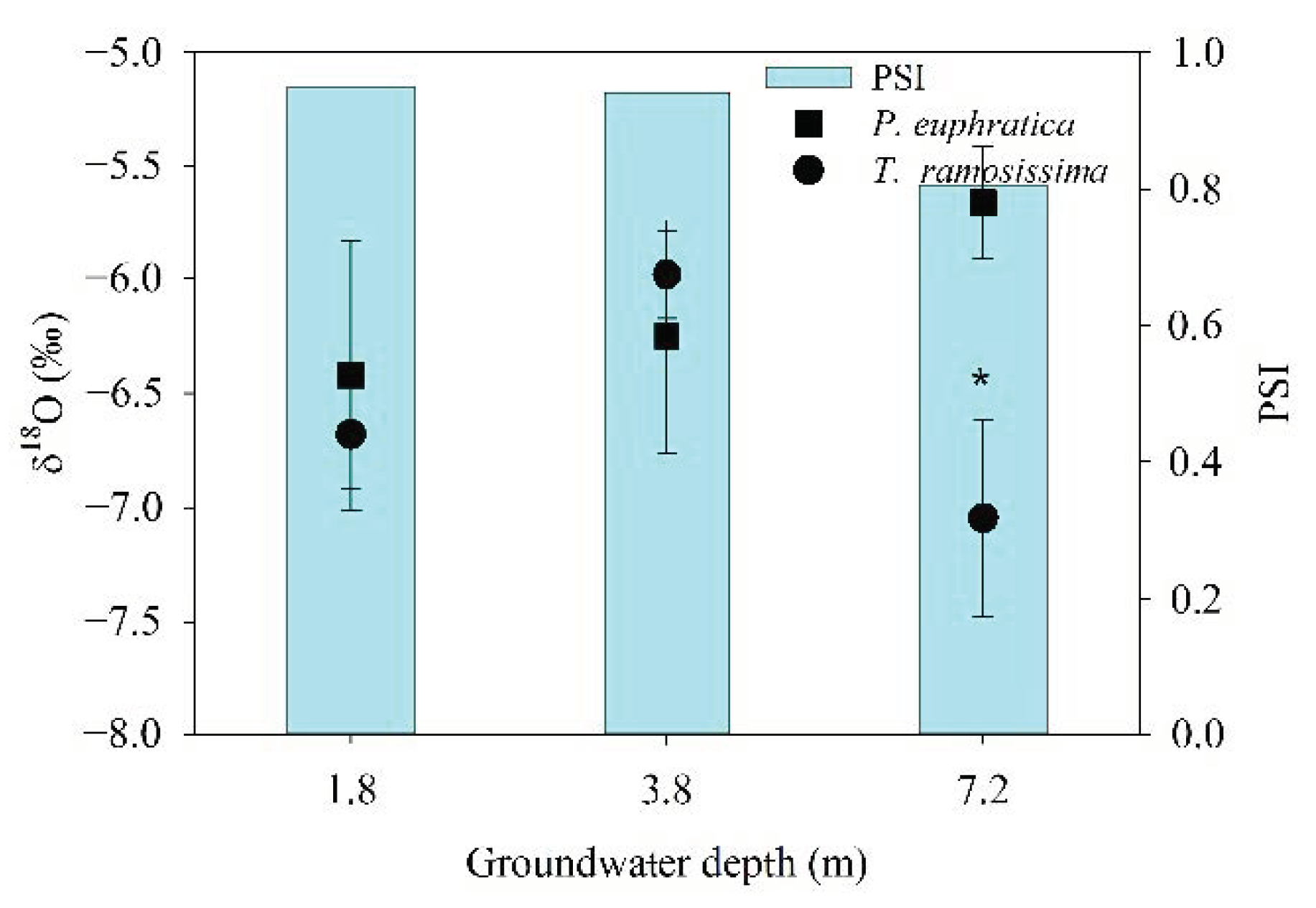
3.1. Individual-Scale Water Use Strategies
3.2. Population-Scale Water Use Strategies
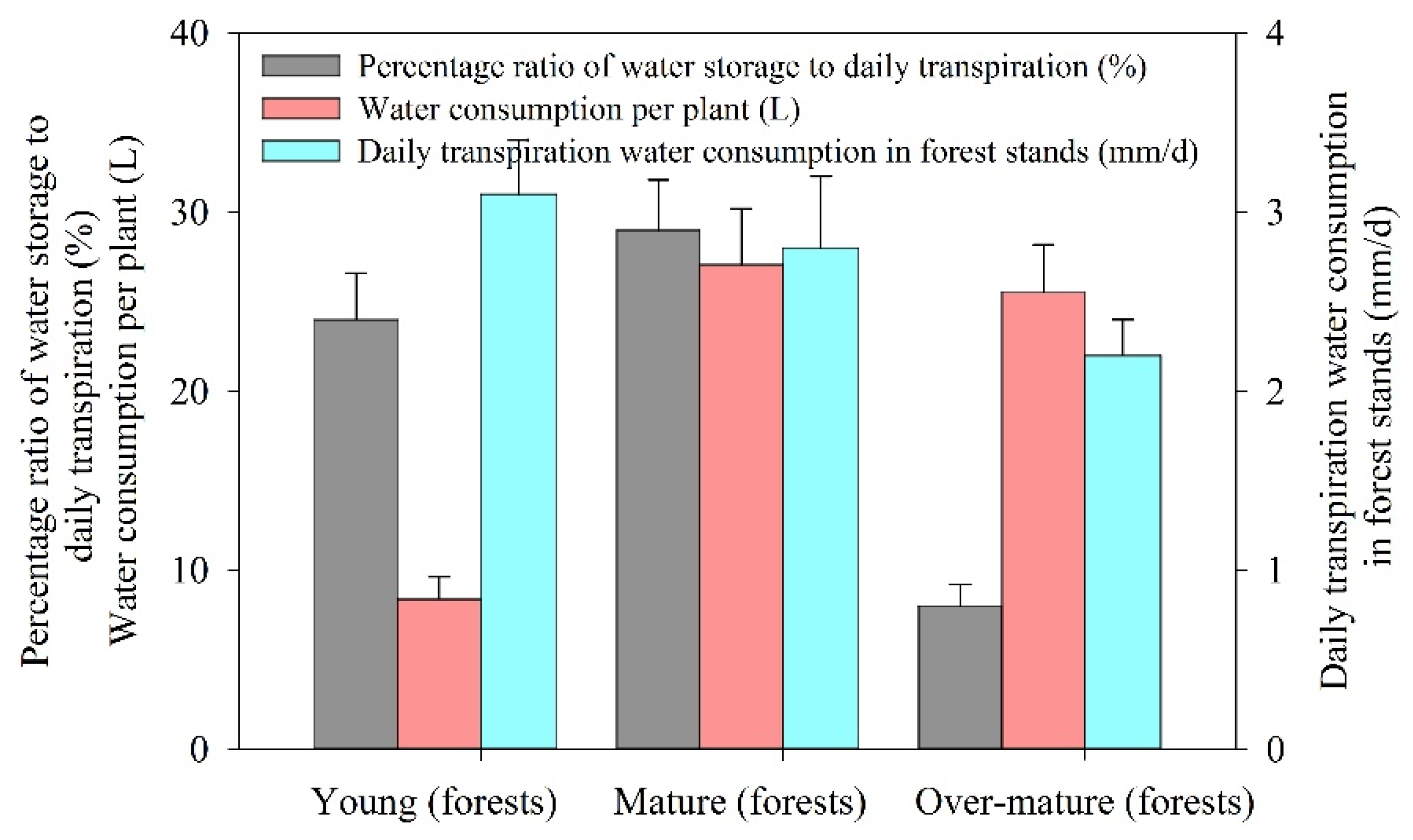
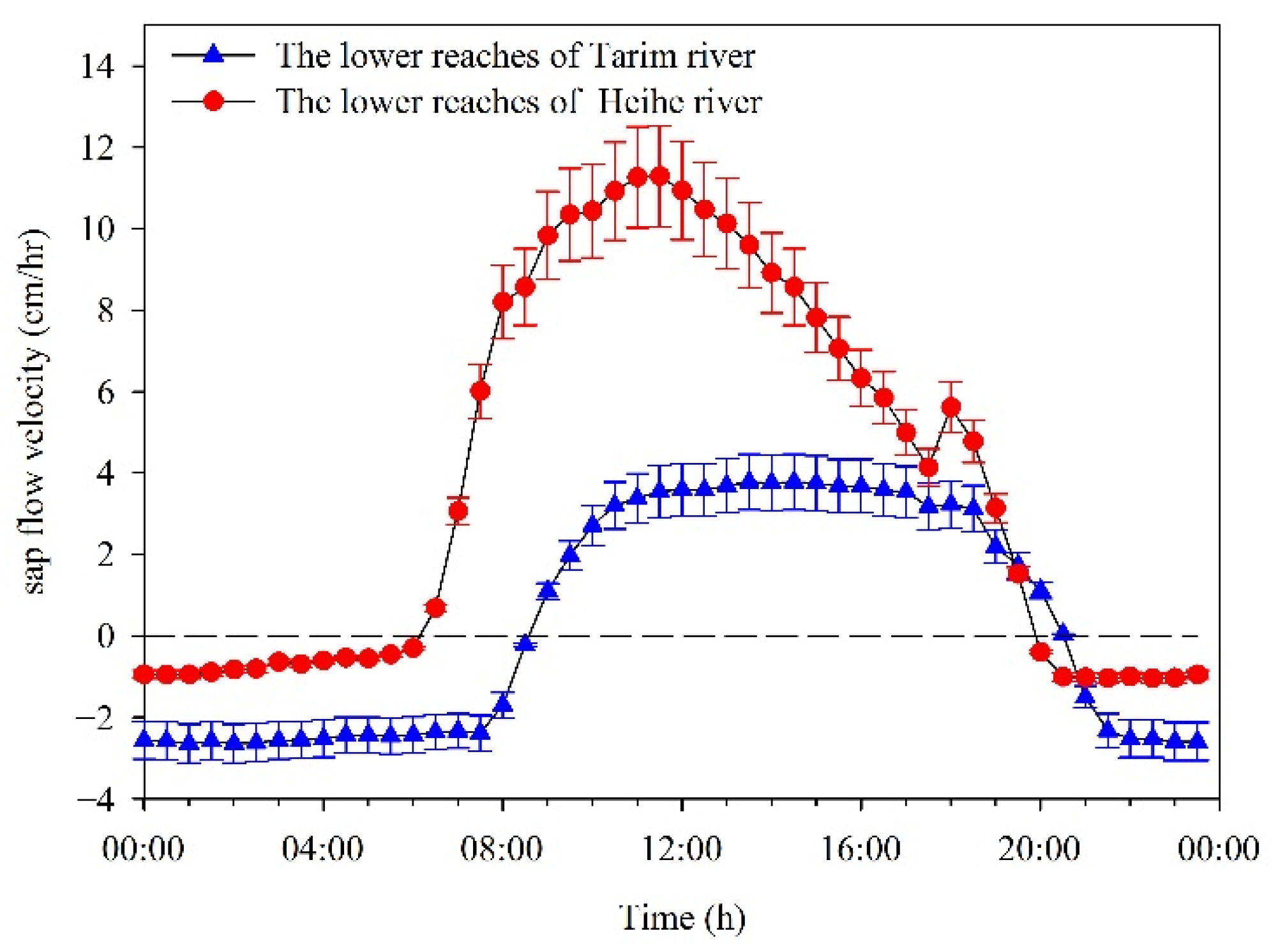
3.3. Transpiration Water Consumption Characteristics of Desert Riparian Forest Plants
4. Interspecific Water Competition and Sharing Mechanisms of Desert Riparian Forest Plants
4.1. Water Competition among Plant Species
4.2. Hydraulic Uplift and Water Redistribution of Plant Root System
4.3. Plant Habitat Range and Plant Life Form Relationship
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ahmed, S.; Kouser, S.; Asgher, M.; Gandhi, S.G. Plant Aquaporins: A frontward to make crop plants drought resistant. Physiol. Plant 2021, 172, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, S.W.; Baskerville, M.; Oelbermann, M.; Gordon, A.M.; Thevathasan, N.V.; Isaac, M.E. Plant diversity and agroecosystem function in riparian agroforests: Providing ecosystem services and land-use transition. Sustainability 2020, 12, 568. [Google Scholar] [CrossRef] [Green Version]
- Zaimes, G.N.; Iakovoglou, V. Assessing riparian areas of Greece—An overview. Sustainability 2021, 13, 309. [Google Scholar] [CrossRef]
- Chen, Y.N. An Overview of the Arid Region Science; Science Press: Beijing, China, 2021. [Google Scholar]
- Ding, Y.J.; Zhao, Q.D.; WU, J.K.; Zhang, S.Q.; Wang, S.X.; Chang, Y.P.; Li, X.Y.; Shangguan, D.H.; Han, H.D.; Qin, J.; et al. The future changes of Chinese cryospheric hydrology and their impacts on water security in arid areas. J. Glaciol. Geocryol. 2020, 42, 23–32. [Google Scholar]
- Chen, Y.; Xu, C.; Chen, P.; Liu, Y.; Li, W. Progress, Challenges and Prospects of Eco-Hydrological Studies in the Tarim River Basin of Xinjiang, China. Environ. Manag. 2013, 51, 138–153. [Google Scholar] [CrossRef]
- Rohde Melissa, M.; Stella John, C.; Roberts Dar, A.; Singer Michael, B. Groundwater dependence of riparian woodlands and the disrupting effect of anthropogenically altered streamflow. Proc. Natl. Acad. Sci. USA 2021, 118, e2026453118. [Google Scholar] [CrossRef]
- Imin, B.; Dai, Y.; Shi, Q.; Guo, Y.; Li, H.; Nijat, M. Responses of two dominant desert plant species to the changes in groundwater depth in hinterland natural oasis, Tarim Basin. Ecol. Evol. 2020, 11, 9460–9471. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Hao, X.; Zhao, Y.; Fang, G.; Yang, Y. Tree rings: A key ecological indicator for reconstruction of groundwater depth in the lower Tarim River, Northwest China. Ecohydrology 2019, 12, e2142. [Google Scholar] [CrossRef]
- Xi, H.Y.; Zhang, J.T.; Feng, Q.; Liu, Z.; Yu, T.F. How changes of groundwater level affect the desert riparian forest ecosystem in the Ejina Oasis, Northwest China. Sci. Cold Arid Reg. 2019, 11, 62–80. [Google Scholar]
- Chen, Y.; Chen, Y.; Xu, C.; Li, W. Groundwater depth affects the daily course of gas exchange parameters of Populus euphratica in arid areas. Environ. Earth Sci. 2012, 66, 433–440. [Google Scholar] [CrossRef]
- Zhu, C.G.; Li, W.H.; Chen, Y.N.; Chen, Y.P. Characteristics of water physiological integration and its ecological significance for Populus euphratica young ramets in an extremely drought environment. J. Geophys. Res. Atmos. 2018, 123, 5657–5666. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Q.; Li, W.; Ruan, X.; Chen, Y.; Zhang, L. Rational groundwater table indicated by the eco-physiological parameters of the vegetation. Chin. Sci. Bull. 2006, 51, 8–15. [Google Scholar] [CrossRef]
- Ruan, X.; Wang, Q.; Pan, C.D.; Chen, Y.N.; Jiang, H. Physiological acclimation strategies of riparian plants to environment change in the delta of the Tarim River, China. Econ. Environ. 2009, 57, 1761–1773. [Google Scholar] [CrossRef]
- Su, Y.H.; Feng, Q.; Zhu, G.F.; Zhang, Q. Evaluating the different methods for estimating groundwater evapotranspiration using diurnal water table fluctuations. J. Hydrol. 2022, 607, 127508. [Google Scholar]
- Xia, J.; Lang, Y.; Zhao, Q.; Liu, P.; Su, L. Photosynthetic characteristics of Tamarix chinensis under different groundwater depths in freshwater habitats. Sci. Total Environ. 2020, 761, 143221. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; Zhu, X.; Li, W.; Zhang, Y.; Xu, H.; Zhang, H.; Chen, Y. Analysis on the ecological benefits of the stream water conveyance to the dried-up river of the lower reaches of Tarim River, China. Sci. China Ser. D Earth Sci. 2004, 47, 1053–1064. [Google Scholar] [CrossRef]
- Chen, Y.N.; Li, W.H.; Xu, H.L.; Liu, J.Z.; Zhang, H.F.; Chen, Y.P. The influence of groundwater on vegetation in the lower reaches of Tarim River, China. Acta Geogr. Sin. 2003, 58, 542–549. [Google Scholar]
- Evenari, M. Adaptations of plants and animals to the desert environment. In Ecosystems of the World: Hot Deserts and Arid Shrublands; Evenari, M., Noy-Meir, I., Goodall, D.W., Eds.; Elsevier Science Publishers: New York, NY, USA, 1985; pp. 79–92. [Google Scholar]
- Luo, Y.Z.; Liu, H.; Yan, G.; Li, G.; Turner, N.C. Roots of Lucerne Seedlings are More Resilient to a Water Deficit than Leaves or Stems. Agronomy 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Saad-Allah, K.M.; Nessem, A.A.; Ebrahim, M.K.H.; Gad, D. Evaluation of Drought Tolerance of Five Maize Genotypes by Virtue of Physiological and Molecular Responses. Agronomy 2021, 12, 59. [Google Scholar] [CrossRef]
- Chen, Y.; Li, W.; Zhou, H.; Chen, Y.; Hao, X.; Fu, A.; Ma, J. Experimental study on water transport observations of desert riparian forests in the lower reaches of the Tarim River in China. Int. J. Biometeorol. 2017, 61, 1055–1062. [Google Scholar] [CrossRef]
- Ayup, M.; Chen, Y.; Nyongesah, M.J.; Zhang, Y.M. Xylem anatomy and hydraulic traits of two co-occurring riparian desert plants. IAWA J. 2015, 36, 69–83. [Google Scholar] [CrossRef]
- Hao, X.M.; Chen, Y.N.; Guo, B.; Ma, J.X. Hydraulic redistribution of soil water in Populus euphratica Oliv. in a central Asian desert riparian forest. Ecohydrology 2013, 6, 974–982. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Li, W.; Ayup, M. Xylem hydraulic conductivity and embolism in riparian plants and their responses to drought stress in desert of Northwest China. Ecohydrology 2013, 6, 984–993. [Google Scholar] [CrossRef]
- Fu, A.; Li, W.; Chen, Y. The threshold of soil moisture and salinity influencing the growth of Populus euphratica and Tamarix ramosissima in the extremely arid region. Environ. Earth Sci. 2012, 66, 2519–2529. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Chen, Y.; Xiang, Y.; Liu, Y.; Kayumba, P.M.; Li, X. Drylands face potential threat of robust drought in the CMIP6 SSPs scenarios. Environ. Res. Lett. 2021, 16, 114004. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Chen, Y.; Liu, Y.; Hu, Y.; Sun, F.; Kayumba, P.M. Projected meteorological drought over Asian drylands under different CMIP6 Scenarios. Remote Sens. 2021, 13, 4409. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Zhu, C.; Li, Z.; Fang, G.; Li, Y.; Fu, A. Climate change may accelerate the decline of desert riparian forest in the lower Tarim River, Northwestern China: Evidence from tree-rings of Populus euphratica. Ecol. Indic. 2020, 111, 105997. [Google Scholar] [CrossRef]
- Ling, H.; Xu, H.; Guo, B.; Deng, X.; Zhang, P.; Wang, X. Regulating water disturbance for mitigating drought stress to conserve and restore a desert riparian forest ecosystem. J. Hydrol. 2019, 572, 659–670. [Google Scholar] [CrossRef]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Ullah, A. Drought Tolerance Strategies in Plants: A Mechanistic Approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Zhu, C.G.; Abula, A.; Li, W.H.; Zhou, H.H. Ecosystem restoration of Populus euphratica forest under the ecological water conveyance in the lower reaches of Tarim River. Arid Land Geogr. 2021, 44, 629–936. [Google Scholar]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide Dismutase and Stress Tolerance. Annu. Rev. Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Zhuang, L.; Chen, Y. Physiological responses of Tamarix ramosissima under water stress along the lower reaches of Tarim River. Chin. Sci. Bull. 2006, 51, 1123–1129. [Google Scholar] [CrossRef]
- Li, D.; Si, J.; Zhang, X.; Gao, Y.; Luo, H.; Qin, J.; Ren, L. Ecological adaptation of Populus euphratica to drought stress. J. Desert Res. 2020, 40, 17–23. [Google Scholar]
- Liu, J.Z.; Chen, Y.N.; Li, W.H.; Chen, Y.J. The process and mechanism of degradation of desert riparian vegetation. Acta Geogr. Sin. 2006, 61, 946–956. [Google Scholar]
- Wang, T.Y.; Wang, P.; Wang, Z.L.; Niu, G.Y.; Yu, J.J.; Ma, N.; Wu, Z.N.; Pozdniakov, S.P.; Yan, D.H. Drought adaptability of phreatophytes: Insight from vertical root distribution in drylands of China. J. Plant Ecol. 2021, 14, 1128–1142. [Google Scholar] [CrossRef]
- Ye, Z.Q.; Wang, J.M.; Wang, W.J.; Zhang, T.H.; Li, J.W. Effects of root phenotypic changes on the deep rooting of Populus euphratica seedlings under drought stresses. PeerJ 2019, 7, e6513. [Google Scholar] [CrossRef] [Green Version]
- Wiehle, M.; Eusemann, P.; Thevs, N.; Schnittler, M. Root suckering patterns in Populus euphratica (Euphrates poplar, Salicaceae). Trees 2009, 23, 991–1001. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, H.; Chen, Y. Adaptation strategies of desert riparian forest vegetation in response and adaption to drought stress. Ecohydrology 2013, 6, 956–973. [Google Scholar] [CrossRef]
- Han, L.; Wang, H.; Niu, J.; Wang, J.; Liu, W. Response of Populus euphratica communities in a desert riparian forest to the groundwater level gradient in the Tarim Basin. Acta Ecol. Sin. 2017, 37, 6836–6846. [Google Scholar]
- Buckley, T.N. How do stomata respond to water status? New Phytol. 2019, 224, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Pirasteh-Anosheh, H.; Saed-Moucheshi, A.; Pakniyat, H.; Pessarakli, M. Stomatal responses to drought stress. In Water Stress and Crop Plants: A Sustainable Approach, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons: New York, NY, USA, 2016; Volume 1, pp. 24–40. [Google Scholar]
- Wankmüller, F.J.P.; Carminati, A. Stomatal regulation prevents plants from critical water potentials during drought: Result of a model linking soil–plant hydraulics to abscisic acid dynamics. Ecohydrology 2021, e2386. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Ruan, X.; Wang, Q.; Chen, Y.N.; Li, W.H.; Xu, Y.N. Physiological response of desert plants to water in hyper arid areas of Tarim River. Acta Ecol. Sin. 2005, 25, 1966–1973. [Google Scholar]
- Chen, Y.; Li, W.; Chen, Y.; Zhou, H.; Hao, X.; Zhu, C. Water Use Process of Constructive Plants in Desert Riparian Forest. Arid. Zone Res. 2018, 35, 130–136. [Google Scholar]
- Fu, A.; Chen, Y.; Li, W. Water use strategies of desert riparian forest plant community in the lower reaches of Heihe River Basin, China. Sci. China Earth Sci. 2014, 57, 1293–1305. [Google Scholar] [CrossRef]
- Ayup, M.; Hao, X.; Chen, Y.; Li, W.; Su, R. Changes of xylem hydraulic efficiency and native embolism of Tamarix ramosissima Ledeb. seedlings under different drought stress conditions and after rewatering. S. Afr. J. Bot. 2012, 78, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Si, J.H.; Zhang, X.Y.; Gao, Y.Y.; Luo, H.; Qin, J.; Gao, G.L. The Mechanism of Changes in Hydraulic Properties of Populus euphratica in Response to Drought Stress. Forests 2019, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Si, J.H.; Zhang, X.Y.; Gao, Y.Y.; Luo, H.; Qin, J.; Gao, G.L. Comparison of Branch Water Relations in Two Riparian Species: Populus euphratica and Tamarix ramosissima. Sustainability 2019, 11, 5461. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.P.; Chen, Y.P.; Chen, Y.N.; Wang, R.Z.; Ren, Z.G. Impact of groundwater depth on leaf hydraulic properties and drought vulnerability of Populus euphratica in the Northwest of China. Trees 2016, 30, 2029–2039. [Google Scholar] [CrossRef]
- Bai, X.; Zhao, C.; Kang, M.P. Effects of groundwater depth on biomass allocation and growth of Populus euphratica seedlings in desert riparian forest in the middle reaches of Shule River. Chin. J. Ecol. 2020, 39, 3605–3612. [Google Scholar]
- Su, P.; Zhang, M.; Qu, D.; Wang, J.; Zhang, Y.; Yao, X.; Xiao, H. Contrasting Water Use Strategies of Tamarix ramosissima in Different Habitats in the Northwest of Loess Plateau, China. Water 2020, 12, 2791. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Chen, Y.; Friedman, J.M.; Hati, J.H.A.; Fang, G. Use of 2H and 18O stable isotopes to investigate water sources for different ages of Populus euphratica along the lower Heihe River. Environ. Res. 2015, 30, 581–587. [Google Scholar] [CrossRef]
- Yu, T.; Feng, Q.; Si, J.; Zhang, X.; Alec, D.; Zhao, C. Evidences and magnitude of nighttime transpiration derived from Populus euphratica in the extreme arid region of China. J. Plant Biol. 2016, 59, 648–657. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Si, J.H.; Feng, Q.; Yu, T.F.; Li, P.D. Comparative study of daytime and nighttime sap flow of Populus euphratica. Plant Growth Regul. 2017, 82, 353–362. [Google Scholar] [CrossRef]
- Chen, Y.N.; Li, W.H.; Zhou, H.H. Analysis of water use strategies of the desert riparian forest plant community in inland rivers of two arid regions in northwestern China. Biogeosci. Discuss. 2014, 11, 14819–14856. [Google Scholar] [CrossRef]
- Ma, J.X.; Huang, X.; Li, W.H.; Zhu, C.G. Sap flow and trunk maximum daily shrinkage (MDS) measurements for diagnosing water status of Populus euphratica in an inland river basin of Northwest China. Ecohydrology 2013, 6, 994–1000. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Xu, C.; Li, W. The effects of groundwater depth on water uptake of Populus euphratica and Tamarix ramosissima in the hyper arid region of northwestern China. Environ. Sci. Pollut. Res. 2016, 23, 17404–17412. [Google Scholar] [CrossRef] [PubMed]
- Tron, S.; Perona, P.; Gorla, L.; Schwarz, M.; Laio, F.; Ridolfi, L. The signature of randomness in riparian plant root distributions. Geophys. Res. Lett. 2015, 42, 7098–7106. [Google Scholar] [CrossRef]
- Yu, T.F.; Feng, Q.; Si, J.H.; Xi, H.Y.; Li, Z.X.; Chen, A.F. Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China’s extremely arid region. Plant Soil 2013, 372, 297–308. [Google Scholar] [CrossRef]
- Wu, G.; Jiang, S.; Liu, W.; Zhao, C.; Li, J. Competition between Populus euphratica and Tamarix ramosissima seedlings under simulated high groundwater availability. J. Arid Land 2016, 8, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Jiang, S.; Liu, H.; Zhu, S.; Zhou, D.; Zhang, Y.; Luo, Q.; Li, J. Early direct competition does not determine the community structure in a desert riparian forest. Sci. Rep. 2018, 8, 4531. [Google Scholar] [CrossRef]
- Chen, X.L.; Chen, Y.N.; Chen, Y.P. Relationship among water use of different plants in Heihe River riparian forests. Chin. J. Eco Agric. 2014, 22, 972–979. [Google Scholar]
- Hao, X.M.; Li, W.H.; Guo, B.; Ma, J.X. Simulation of the effect of root distribution on hydraulic redistribution in a desert riparian forest. Environ. Res. 2013, 28, 653–662. [Google Scholar] [CrossRef]
- Hao, X.M.; Li, Y.; Deng, H.J. Assessment of hydraulic redistribution on desert riparian forests in an extremely arid area. Environ. Monit. Assess. 2013, 185, 10027–10038. [Google Scholar] [CrossRef]
- Yu, T.F.; Feng, Q.; Si, J.H.; Xi, H.Y.; Li, W. Patterns, magnitude, and controlling factors of hydraulic redistribution of soil water by Tamarix ramosissima roots. J. Arid Land 2013, 5, 396–407. [Google Scholar] [CrossRef] [Green Version]
- Kikvidze, Z.; Pugnaire, F.I.; Brooker, R.W.; Choler, P.; Lortie, C.J.; Michalet, R.; Callaway, R.M. Linking patterns and processes in alpine plant communities: A global study. Ecology 2005, 86, 1395–1400. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Dubois, D.; Ellenberg, H. Aims and Methods of Vegetation; John Wiley & Sons: New York, NY, USA, 1974; pp. 139–147. [Google Scholar]
- Bejarano, M.D.; Sarneel, J.; Su, X.; Sordo-Ward, A. Shifts in Riparian Plant Life Forms Following Flow Regulation. Forests 2020, 11, 518. [Google Scholar] [CrossRef]
- Gao, X.M.; Chen, L.Z. The revision of plant life-form system and an analysis of the life-form spectrum of forest plants in warn temperate zone of China. Acta Bot. Sin. 1998, 40, 553–559. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Chlorophyll | Soluble Sugar | PRO | MDA | SOD | POD |
|---|---|---|---|---|---|---|
| Populus euphratica | −0.67 ** | 0.33 | 0.49 * | 0.84 ** | 0.73 ** | −0.80 ** |
| Tamarix ramosissima | −0.77 ** | 0.44 | 0.50 * | 0.66 ** | 0.74** | −0.81 ** |
| Phragmites australis | −0.71 | 0.32 | 0.84 * | 0.82 * | 0.87 * | −0.62 |
| Adaptive Strategies | Populus euphratica | Tamarix ramosissima |
|---|---|---|
| Mutual coordination of various physiological metabolisms | Increasing soluble sugars, PRO, MDA, and SOD while decreasing activity of POD | Increasing soluble sugars, PRO, MDA and SOD while decreasing activity of POD |
| Damage-avoidance mechanisms | Mainly regulated by soluble sugars | Mainly regulated by PRO |
| Drought-resistance mechanism | Increasing ABA while decreasing CTK | Increasing both ABA and CTK in mild drought; decreasing both ABA and CTK in stronger drought |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Chen, Y.; Zhou, H.; Hao, X.; Zhu, C.; Fu, A.; Yang, Y.; Li, W. Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress. Forests 2022, 13, 619. https://doi.org/10.3390/f13040619
Chen Y, Chen Y, Zhou H, Hao X, Zhu C, Fu A, Yang Y, Li W. Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress. Forests. 2022; 13(4):619. https://doi.org/10.3390/f13040619
Chicago/Turabian StyleChen, Yaning, Yapeng Chen, Honghua Zhou, Xingming Hao, Chenggang Zhu, Aihong Fu, Yuhai Yang, and Weihong Li. 2022. "Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress" Forests 13, no. 4: 619. https://doi.org/10.3390/f13040619
APA StyleChen, Y., Chen, Y., Zhou, H., Hao, X., Zhu, C., Fu, A., Yang, Y., & Li, W. (2022). Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress. Forests, 13(4), 619. https://doi.org/10.3390/f13040619