
Effects of Clonal Integration on Foraging Behavior of Three Clonal Plants in Heterogeneous Soil Environments



Abstract
:1. Introduction
2. Materials and Methods
2.1. The Species
2.2. Sampling and Cultivation
2.3. Experimental Design
2.4. Measurements
2.5. Data Analysis
3. Results
3.1. Effects of Clonal Integration and Soil Heterogeneity on Biomass of the Mother in the Pot
3.2. Effects of Clonal Integration and Soil Heterogeneity on Offspring Growth in the Box
3.3. Effects of Clonal Integration on Foraging Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, J.J.; Price, J.S. Fitness and evolution in clonal plants: The impact of clonal growth. Evol. Ecol. 2002, 15, 583–600. [Google Scholar] [CrossRef]
- Eckert, C.G. The loss of sex in clonal plants. Evol. Ecol. 2002, 15, 501–520. [Google Scholar] [CrossRef]
- Klimes, L.; Klimesova, J.; Hendriks, R.; Groenendael, J.V. Clonal Plant Architecture: A Comparative Analysis of Form and Function. In The Ecology and Evolution of Clonal Plants; De Kroon, H., Van Groenendael, J., Eds.; Backhuys: Leiden, The Netherlands, 1997; pp. 1–29. [Google Scholar]
- Jackson, J.B.; Buss, L.W.; Cook, R.E.; Ashmun, J.W. Population Biology and Evolution of Clonal Organisms; Yale University Press: New Haven, CT, USA, 1985. [Google Scholar]
- Xu, L.; Yu, F.-H.; Van Drunen, E.; Schieving, F.; Dong, M.; Anten, N.P. Trampling, defoliation and physiological integration affect growth, morphological and mechanical properties of a root-suckering clonal tree. Ann. Bot. 2012, 109, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adomako, M.O.; Alpert, P.; Du, D.-L.; Yu, F.-H. Effects of clonal integration, nutrients and cadmium on growth of the aquatic macrophyte Pistia stratiotes. J. Plant Ecol. 2020, 13, 765–772. [Google Scholar] [CrossRef]
- Wang, P.; Alpert, P.; Yu, F.-H. Physiological integration can increase competitive ability in clonal plants if competition is patchy. Oecologia 2021, 195, 199–212. [Google Scholar] [CrossRef]
- Gao, F.L.; Alpert, P.; Yu, F.H. Parasitism induces negative effects of physiological integration in a clonal plant. New Phytol. 2021, 229, 585–592. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. The advantages of clonal integration under different ecological conditions: A community-wide test. Ecology 2000, 81, 709–716. [Google Scholar] [CrossRef]
- Stuefer, J.F.; During, H.J.; De Kroon, H. High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments. J. Ecol. 1994, 82, 511–518. [Google Scholar] [CrossRef]
- Si, C.; Alpert, P.; Zhang, J.-F.; Lin, J.; Wang, Y.-Y.; Hong, M.-M.; Roiloa, S.R.; Yu, F.-H. Capacity for clonal integration in introduced versus native clones of the invasive plant Hydrocotyle vulgaris. Sci. Total Environ. 2020, 745, 141056. [Google Scholar] [CrossRef]
- Gómez, S.; Onoda, Y.; Ossipov, V.; Stuefer, J.F. Systemic induced resistance: A risk-spreading strategy in clonal plant networks? New Phytol. 2008, 179, 1142–1153. [Google Scholar] [CrossRef]
- Lu, H.Z.; Brooker, R.; Song, L.; Liu, W.Y.; Sack, L.; Zhang, J.L.; Yu, F.H. When facilitation meets clonal integration in forest canopies. New Phytol. 2020, 225, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-P.; Wei, G.-W.; Luo, F.-L.; Li, C.-Y.; Dong, B.-C.; Ji, J.-S.; Yu, F.-H. Effects of salinity and clonal integration on the amphibious plant Paspalum paspaloides: Growth, photosynthesis and tissue ion regulation. J. Plant Ecol. 2019, 12, 45–55. [Google Scholar] [CrossRef]
- Amsberry, L.; Baker, M.A.; Ewanchuk, P.J.; Bertness, M.D. Clonal integration and the expansion of Phragmites australis. Ecol. Appl. 2000, 10, 1110–1118. [Google Scholar] [CrossRef]
- Oborny, B.; Kun, Á.; Czárán, T.; Bokros, S. The effect of clonal integration on plant competition for mosaic habitat space. Ecology 2000, 81, 3291–3304. [Google Scholar] [CrossRef]
- Hutchings, M.J.; Wijesinghe, D.K. Performance of a clonal species in patchy environments: Effects of environmental context on yield at local and whole-plant scales. Evol. Ecol. 2008, 22, 313–324. [Google Scholar] [CrossRef]
- Song, Y.-B.; Yu, F.-H.; Keser, L.H.; Dawson, W.; Fischer, M.; Dong, M.; Van Kleunen, M. United we stand, divided we fall: A meta-analysis of experiments on clonal integration and its relationship to invasiveness. Oecologia 2013, 171, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, T.; Wang, Y.; Li, G.; Abdullah, I.; Zhong, Z.; Liu, J.; Zhu, W.; Wang, L.; Wang, D.; et al. A meta-analysis of effects of physiological integration in clonal plants under homogeneous vs. heterogeneous environments. Funct. Ecol. 2021, 35, 578–589. [Google Scholar] [CrossRef]
- Stuefer, J.F.; Gómez, S.; Mölken, T.V. Clonal integration beyond resource sharing: Implications for defence signalling and disease transmission in clonal plant networks. Evol. Ecol. 2004, 18, 647–667. [Google Scholar] [CrossRef]
- Jackson, R.; Caldwell, M. Geostatistical patterns of soil heterogeneity around individual perennial plants. J. Ecol. 1993, 81, 683–692. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, X.; Yu, F.; Dong, M.; Zhang, S.; Wang, R. Spatial heterogeneity of Ulmus pumila open forest ecosystem in Otindag sandy land. Environ. Sci. 2003, 24, 29–34. [Google Scholar]
- Alpert, P.; Mooney, H.A. Resource heterogeneity generated by shrubs and topography on coastal sand dunes. Vegetatio 1996, 122, 83–93. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Environmental heterogeneity and plant species diversity: A hypothesis. Am. Nat. 1977, 111, 376–381. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Birch, C.; Hutchings, M. Exploitation of patchily distributed soil resources by the clonal herb Glechoma hederacea. J. Ecol. 1994, 82, 653–664. [Google Scholar] [CrossRef]
- Adomako, M.O.; Xue, W.; Roiloa, S.; Zhang, Q.; Du, D.-L.; Yu, F.-H. Earthworms modulate impacts of soil heterogeneity on plant growth at different spatial scales. Front. Plant Sci. 2021, 12, 735495. [Google Scholar] [CrossRef]
- Adomako, M.O.; Xue, W.; Du, D.-L.; Yu, F.-H. Soil biota and soil substrates influence responses of the rhizomatous clonal grass Leymus chinensis to nutrient heterogeneity. Plant Soil 2021, 465, 19–29. [Google Scholar] [CrossRef]
- Hutchings, M.; De Kroon, H. Foraging in plants: The role of morphological plasticity in resource acquisition. Adv. Ecol. Res. 1994, 25, 159–238. [Google Scholar]
- De Kroons, H.; Hutchings, M.J. Morphological plasticity in clonal plants: The foraging concept reconsidered. J. Ecol. 1995, 83, 143–152. [Google Scholar] [CrossRef]
- Si, C.; Xue, W.; Guo, Z.-W.; Zhang, J.-F.; Hong, M.-M.; Wang, Y.-Y.; Lin, J.; Yu, F.-H. Soil heterogeneity and earthworms independently promote growth of two bamboo species. Ecol. Indic. 2021, 130, 108068. [Google Scholar] [CrossRef]
- Wijesinghe, D.K.; Hutchings, M.J. The effects of environmental heterogeneity on the performance of Glechoma hederacea: The interactions between patch contrast and patch scale. J. Ecol. 1999, 87, 860–872. [Google Scholar] [CrossRef]
- Liu, L.; Alpert, P.; Dong, B.-C.; Yu, F.-H. Modification by earthworms of effects of soil heterogeneity and root foraging in eight species of grass. Sci. Total Environ. 2020, 708, 134941. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Alpert, P.; Dong, B.-C.; Li, J.-M.; Yu, F.-H. Combined effects of soil heterogeneity, herbivory and detritivory on growth of the clonal plant Hydrocotyle vulgaris. Plant Soil 2017, 421, 429–437. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Fischer, M. Adaptive evolution of plastic foraging responses in a clonal plant. Ecology 2001, 82, 3309–3319. [Google Scholar] [CrossRef]
- Liang, J.-F.; Yuan, W.-Y.; Gao, J.-Q.; Roiloa, S.R.; Song, M.-H.; Zhang, X.-Y.; Yu, F.-H. Soil resource heterogeneity competitively favors an invasive clonal plant over a native one. Oecologia 2020, 193, 155–165. [Google Scholar] [CrossRef]
- Fransen, B.; De Kroon, H.; Berendse, F. Soil nutrient heterogeneity alters competition between two perennial grass species. Ecology 2001, 82, 2534–2546. [Google Scholar] [CrossRef]
- Xue, W.; Huang, L.; Yu, F.-H.; Bezemer, T.M. Intraspecific aggregation and soil heterogeneity: Competitive interactions of two clonal plants with contrasting spatial architecture. Plant Soil 2018, 425, 231–240. [Google Scholar] [CrossRef]
- Liu, L.; Dong, B.-C.; Alpert, P.; Yu, F.-H. Effects of soil substrate heterogeneity and moisture on interspecific competition between Alternanthera philoxeroides and four native species. J. Plant Ecol. 2017, 10, 528–537. [Google Scholar]
- Louâpre, P.; Bittebière, A.-K.; Clément, B.; Pierre, J.-S.; Mony, C. How past and present influence the foraging of clonal plants? PLoS ONE 2012, 7, e38288. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Yu, J.; Zhang, L.; Lei, N.; Xue, W.; Chen, J.; Zhang, Q.; Yu, F. Effects of soil nutrient heterogeneity and earthworms on aboveground biomass of experimental plant communities. Phyton 2021, 90, 1259. [Google Scholar] [CrossRef]
- Xue, W.; Huang, L.; Yu, F.-H. Increasing soil configurational heterogeneity promotes plant community evenness through equalizing differences in competitive ability. Sci. Total Environ. 2021, 750, 142308. [Google Scholar] [CrossRef]
- Hutchings, M.J.; John, E.A.; Wijesinghe, D.K. Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 2003, 84, 2322–2334. [Google Scholar] [CrossRef]
- Adomako, M.O.; Gao, F.-L.; Li, J.-M.; Du, D.-L.; Xue, W.; Yu, F.-H. Effects of soil nutrient heterogeneity and parasitic plant infection on an experimental grassland community. Flora 2020, 271, 151666. [Google Scholar] [CrossRef]
- Wijesinghe, D.K.; John, E.A.; Hutchings, M.J. Does pattern of soil resource heterogeneity determine plant community structure? An experimental investigation. J. Ecol. 2005, 93, 99–112. [Google Scholar] [CrossRef]
- Geng, Y.; Van Klinken, R.D.; Sosa, A.; Li, B.; Chen, J.; Xu, C.-Y. The relative importance of genetic diversity and phenotypic plasticity in determining invasion success of a clonal weed in the USA and China. Front. Plant Sci. 2016, 7, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.-Y.; Alpert, P.; An, J.; Gao, J.-Q.; Han, G.-X.; Yu, F.-H. Clonal integration in Phagmites australis mitigates effects of oil pollution on greenhouse gas emissions in a coastal wetland. Sci. Total Environ. 2020, 739, 140007. [Google Scholar] [CrossRef]
- Xue, W.; Wang, W.-L.; Yuan, Q.-Y.; Yu, F.-H. Clonal integration in Phragmites australis alters soil microbial communities in an oil-contaminated wetland. Environ. Pollut. 2020, 265, 114828. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Lu, H.-Z.; Alpert, P.; Song, L.; Liu, W.-Y.; Yu, F.-H. Higher benefits of clonal integration in rhizome-derived than in frond-derived ramets of the tropical fern Bolbitis heteroclita. Flora 2019, 257, 151415. [Google Scholar] [CrossRef]
- Dong, B.-C.; Zhang, L.-M.; Li, K.-Y.; Hu, X.-T.; Wang, P.; Wang, Y.-J.; Luo, F.-L.; Li, H.-L.; Yu, F.-H. Effects of clonal integration and nitrogen supply on responses of a clonal plant to short-term herbivory. J. Plant Ecol. 2019, 12, 624–635. [Google Scholar] [CrossRef]
- Portela, R.; Dong, B.-C.; Yu, F.-H.; Barreiro, R.; Roiloa, S.R. Effects of physiological integration on defense strategies against herbivory by the clonal plant Alternanthera philoxeroides. J. Plant Ecol. 2019, 12, 662–672. [Google Scholar] [CrossRef]
- Lin, H.-F.; Alpert, P.; Zhang, Q.; Yu, F.-H. Facilitation of amphibious habit by physiological integration in the clonal, perennial, climbing herb Ipomoea aquatica. Sci. Total Environ. 2018, 618, 262–268. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; Van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Ali, A.; Yong, X.-H.; Lin, C.-G.; Niu, X.-H.; Cai, A.-M.; Dong, B.-C.; Zhou, Z.-X.; Wang, Y.-J.; Yu, F.-H. A multi-species comparison of selective placement patterns of ramets in invasive alien and native clonal plants to light, soil nutrient and water heterogeneity. Sci. Total Environ. 2019, 657, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.C.; Wang, J.Z.; Liu, R.H.; Zhang, M.X.; Luo, F.L.; Yu, F.H. Soil heterogeneity affects ramet placement of Hydrocotyle vulgaris. J. Plant Ecol. 2014, 8, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Cain, M.L. Consequences of foraging in clonal plant species. Ecology 1994, 75, 933–944. [Google Scholar] [CrossRef]
- Liu, F.; Liu, J.; Dong, M. Ecological consequences of clonal integration in plants. Front. Plant Sci. 2016, 7, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xing, F.; Jin, Y.; Nie, D.; Wang, Y. Foraging responses of clonal plants to multi-patch environmental heterogeneity: Spatial preference and temporal reversibility. Plant Soil 2012, 359, 137–147. [Google Scholar] [CrossRef]
- Keser, L.H.; Dawson, W.; Song, Y.-B.; Yu, F.-H.; Fischer, M.; Dong, M.; Van Kleunen, M. Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 2014, 174, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.-H.; Ji, M.-C.; Wang, Y.-Y.; Qiao, D.-D.; Chen, Y.-C. Study on invasion risk of Hydrocotyle vulgaris as an alien species in wetlands. J. Zhejiang Univ. Agric. Life Sci. 2011, 37, 425–431. [Google Scholar]
- Murphy, K.J.; Rørslett, B.; Springuel, I. Strategy analysis of submerged lake macrophyte communities: An international example. Aquat. Bot. 1990, 36, 303–323. [Google Scholar] [CrossRef]
- Wang, M.-Z.; Li, H.-L.; Liu, C.-X.; Dong, B.-C.; Yu, F.-H. Adaptive plasticity in response to light and nutrient availability in the clonal plant Duchesnea indica. J. Plant Ecol. 2022. [Google Scholar] [CrossRef]
- Liao, M.; Yu, F.; Song, M.; Zhang, S.; Zhang, J.; Dong, M. Plasticity in R/S ratio, morphology and fitness-related traits in response to reciprocal patchiness of light and nutrients in the stoloniferous herb, Glechoma longituba L. Acta Oecol. 2003, 24, 231–239. [Google Scholar] [CrossRef]
- Gao, J.-Q.; Duan, M.-Y.; Zhang, X.-Y.; Li, Q.-W.; Yu, F.-H. Effects of frequency and intensity of drying-rewetting cycles on Hydrocotyle vulgaris growth and greenhouse gas emissions from wetland microcosms. Catena 2018, 164, 44–49. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Retuerto, R. Small-scale heterogeneity in soil quality influences photosynthetic efficiency and habitat selection in a clonal plant. Ann. Bot. 2006, 98, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Y.; Schooler, S.S.; Van Klinken, R.D. Effects of clonal integration and light availability on the growth and physiology of two invasive herbs. J. Ecol. 2010, 98, 833–844. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Chen, Y.; Dong, M. Clonal integration enhances survival and performance of Potentilla anserina, suffering from partial sand burial on Ordos plateau, China. Evol. Ecol. 2002, 15, 303–318. [Google Scholar] [CrossRef]
- Estrada, J.A.; Wilson, C.H.; Flory, S.L. Clonal integration enhances performance of an invasive grass. Oikos 2020, 129, 1623–1631. [Google Scholar] [CrossRef]
- Pauliukonis, N.; Gough, L. Effects of the loss of clonal integration on four sedges that differ in ramet aggregation. Plant Ecol. 2004, 173, 1–15. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; During, H.J.; Li, L. Do clonal plants show greater division of labour morphologically and physiologically at higher patch contrasts? PLoS ONE 2011, 6, e25401. [Google Scholar] [CrossRef]
- Martínková, J.; Klimeš, A.; Klimešová, J. No evidence for nutrient foraging in root-sprouting clonal plants. Basic Appl. Ecol. 2018, 28, 27–36. [Google Scholar] [CrossRef]
- Waters, E.M.; Soini, H.A.; Novotny, M.V.; Watson, M.A. Volatile organic compounds (VOCs) drive nutrient foraging in the clonal woodland strawberry, Fragaria vesca. Plant Soil 2016, 407, 261–274. [Google Scholar] [CrossRef]
- Slade, A.; Hutchings, M. The effects of light intensity on foraging in the clonal herb Glechoma hederacea. J. Ecol. 1987, 75, 639–650. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Alpert, P.; Yu, F.-H. Nutrient foraging ability promotes intraspecific competitiveness in the clonal plant Hydrocotyle vulgaris. Ecol. Indic. 2022, 138, 108862. [Google Scholar] [CrossRef]
- Wang, P.; Lei, J.-P.; Li, M.-H.; Yu, F.-H. Spatial heterogeneity in light supply affects intraspecific competition of a stoloniferous clonal plant. PLoS ONE 2012, 7, e39105. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, D.K.; Hutchings, M.J. The effects of spatial scale of environmental heterogeneity on the growth of a clonal plant: An experimental study with Glechoma hederacea. J. Ecol. 1997, 85, 17–28. [Google Scholar] [CrossRef]





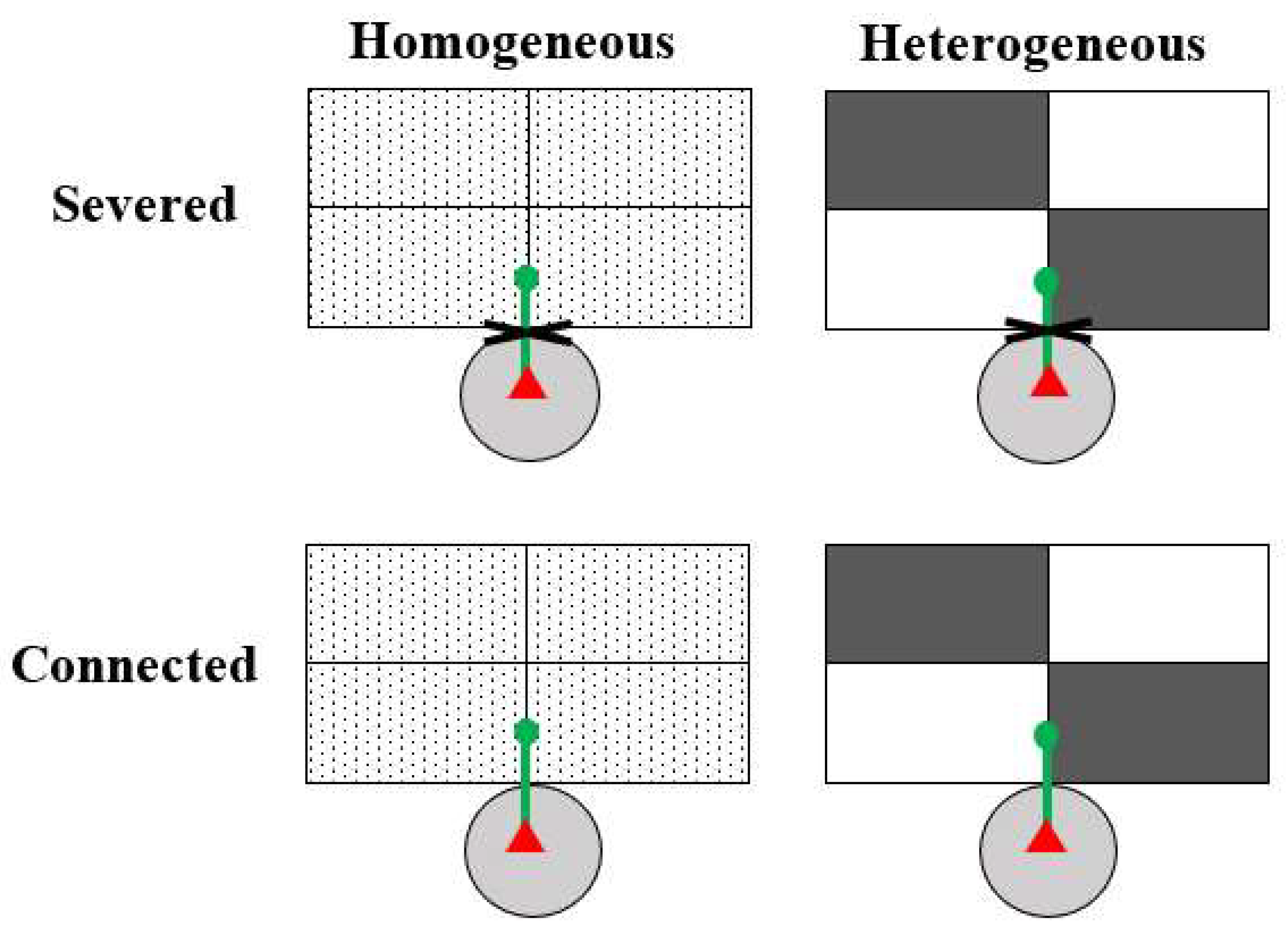{kind=link}
{kind=link}
{kind=link}
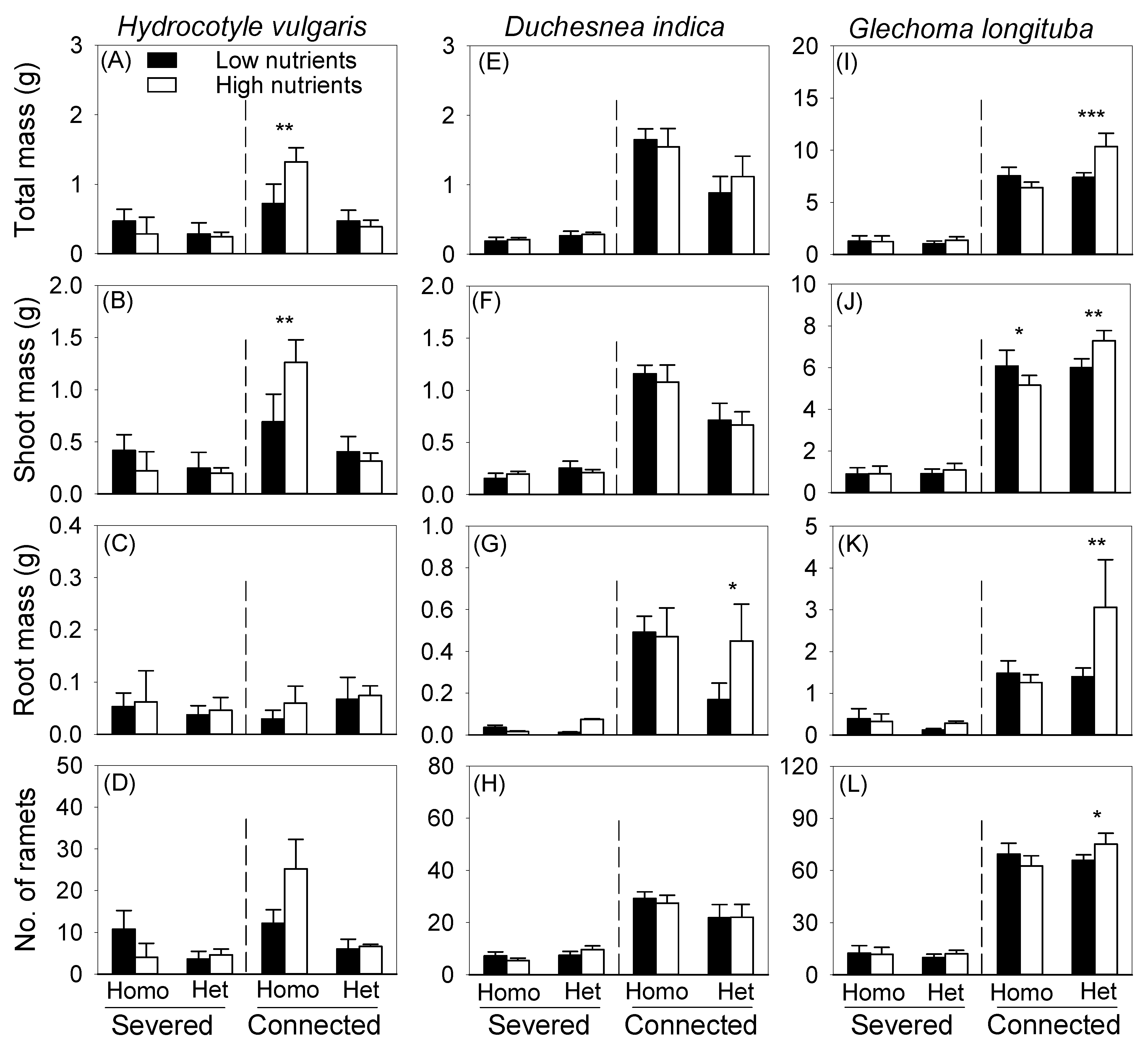{kind=link}
| Effect | Total Mass | Shoot Mass | Root Mass | |||||
|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |||
| (A) Hydrocotyle vulgaris | ||||||||
| I | 0.05 | 0.834 | 0.03 | 0.874 | 0.95 | 0.344 | ||
| H | 2.05 | 0.171 | 0.99 | 0.334 | 4.47 | 0.051 | ||
| I × H | 0.84 | 0.372 | 0.39 | 0.543 | 0.57 | 0.463 | ||
| (B) Duchesnea indica | ||||||||
| I | 4.39 | 0.052 | 4.87 | 0.042 | 0.86 | 0.367 | ||
| H | 0.94 | 0.347 | 1.46 | 0.244 | 0.15 | 0.701 | ||
| I × H | 0.62 | 0.442 | 1.32 | 0.268 | 0.17 | 0.687 | ||
| (C) Glechoma longituba | ||||||||
| I | 0.01 | 0.923 | 1.14 | 0.303 | 2.170 | 0.160 | ||
| H | 1.80 | 0.199 | 1.87 | 0.190 | 0.58 | 0.459 | ||
| I × H | 2.24 | 0.154 | 1.93 | 0.183 | 1.02 | 0.327 | ||
| Effect | Total Mass | Shoot Mass | Root Mass | No. of Ramets | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | ||||
| (A) Hydrocotyle vulgaris | |||||||||||
| I | 9.8 | 0.007 | 11.2 | 0.004 | <0.1 | 0.963 | 5.3 | 0.036 | |||
| H | 6.1 | 0.026 | 7.3 | 0.016 | 0.1 | 0.759 | 5.8 | 0.030 | |||
| I × H | 3.5 | 0.081 | 4.4 | 0.054 | 0.2 | 0.661 | 1.3 | 0.280 | |||
| (B) Duchesnea indica | |||||||||||
| I | 48.9 | <0.001 | 45.2 | <0.01 | 25.9 | <0.001 | 69.3 | <0.001 | |||
| H | 1.0 | 0.758 | 2.4 | 0.145 | 0.1 | 0.750 | <0.1 | 0.944 | |||
| I × H | 4.4 | 0.051 | 4.9 | 0.050 | 4.0 | 0.063 | 5.3 | 0.034 | |||
| (C) Glechoma longituba | |||||||||||
| I | 122.4 | <0.001 | 156.4 | <0.01 | 49.5 | <0.001 | 192.8 | <0.001 | |||
| H | 2.1 | 0.167 | 1.8 | 0.203 | < 0.1 | 0.953 | 0.2 | 0.683 | |||
| I × H | 2.4 | 0.137 | 1.1 | 0.311 | 2.4 | 0.139 | 0.5 | 0.503 | |||
| Effect | Total Mass | Shoot Mass | Root Mass | No. of Ramets | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | ||||
| (A) Hydrocotyle vulgaris | |||||||||||
| Between subject | |||||||||||
| I | 6.0 | 0.027 | 6.7 | 0.021 | 0.1 | 0.765 | 4.8 | 0.044 | |||
| H | 4.6 | 0.050 | 5.4 | 0.035 | <0.1 | 0.847 | 6.5 | 0.022 | |||
| I × H | 2.1 | 0.167 | 2.9 | 0.111 | 0.6 | 0.432 | 2.2 | 0.159 | |||
| Within subject | |||||||||||
| Q | 0.9 | 0.362 | 0.7 | 0.425 | 0.6 | 0.445 | 1.2 | 0.300 | |||
| Q × I | 5.7 | 0.031 | 6.2 | 0.025 | 0.1 | 0.778 | 7.0 | 0.018 | |||
| Q × H | 3.0 | 0.104 | 3.2 | 0.096 | 0.1 | 0.742 | 0.4 | 0.534 | |||
| Q × I × H | 7.1 | 0.018 | 7.7 | 0.014 | 0.1 | 0.744 | 7.6 | 0.015 | |||
| (B) Duchesnea indica | |||||||||||
| Between subject | |||||||||||
| I | 45.1 | 0.001 | 55.8 | 0.001 | 24.9 | 0.001 | 41.0 | 0.001 | |||
| H | 2.8 | 0.117 | 3.9 | 0.065 | 1.1 | 0.306 | 0.6 | 0.458 | |||
| I × H | 4.5 | 0.049 | 6.7 | 0.020 | 1.7 | 0.212 | 2.4 | 0.139 | |||
| Within subject | |||||||||||
| Q | 0.3 | 0.571 | 0.5 | 0.476 | 2.3 | 0.148 | 0.1 | 0.811 | |||
| Q × I | 0.1 | 0.764 | 0.5 | 0.479 | 1.2 | 0.289 | 0.2 | 0.691 | |||
| Q × H | 1.2 | 0.288 | 0.1 | 0.781 | 3.7 | 0.071 | 1.5 | 0.242 | |||
| Q × I × H | 1.2 | 0.281 | 0.5 | 0.494 | 1.2 | 0.283 | 0.2 | 0.691 | |||
| (C) Glechoma longituba | |||||||||||
| Between subject | |||||||||||
| I | 125.9 | 0.001 | 167.6 | 0.001 | 17.5 | 0.001 | 192.8 | 0.001 | |||
| H | 2.3 | 0.147 | 2.0 | 0.180 | 0.9 | 0.350 | 0.2 | 0.683 | |||
| I × H | 2.6 | 0.124 | 1.3 | 0.264 | 1.9 | 0.183 | 0.5 | 0.503 | |||
| Within subject | |||||||||||
| Q | 3.2 | 0.094 | 0.5 | 0.487 | 2.3 | 0.150 | 0.2 | 0.627 | |||
| Q × I | 1.7 | 0.215 | 0.1 | 0.818 | 1.7 | 0.206 | <0.1 | 0.884 | |||
| Q × H | 14.5 | 0.002 | 9.0 | 0.008 | 4.3 | 0.053 | 5.4 | 0.033 | |||
| Q × I × H | 9.9 | 0.006 | 6.7 | 0.020 | 2.7 | 0.121 | 2.8 | 0.111 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.-X.; Xue, W.; Lei, N.-F.; Yu, F.-H. Effects of Clonal Integration on Foraging Behavior of Three Clonal Plants in Heterogeneous Soil Environments. Forests 2022, 13, 696. https://doi.org/10.3390/f13050696
Cao X-X, Xue W, Lei N-F, Yu F-H. Effects of Clonal Integration on Foraging Behavior of Three Clonal Plants in Heterogeneous Soil Environments. Forests. 2022; 13(5):696. https://doi.org/10.3390/f13050696
Chicago/Turabian StyleCao, Xiao-Xiao, Wei Xue, Ning-Fei Lei, and Fei-Hai Yu. 2022. "Effects of Clonal Integration on Foraging Behavior of Three Clonal Plants in Heterogeneous Soil Environments" Forests 13, no. 5: 696. https://doi.org/10.3390/f13050696
APA StyleCao, X. -X., Xue, W., Lei, N. -F., & Yu, F. -H. (2022). Effects of Clonal Integration on Foraging Behavior of Three Clonal Plants in Heterogeneous Soil Environments. Forests, 13(5), 696. https://doi.org/10.3390/f13050696