
Endogenous Hormones and Biochemical Changes during Flower Development and Florescence in the Buds and Leaves of Lycium ruthenicum Murr

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Hormone Analysis
2.3. Determination of Soluble Total Sugar and Sucrose
2.4. Starch Content
2.5. Soluble Proteins
2.6. Statistical Analysis
3. Results
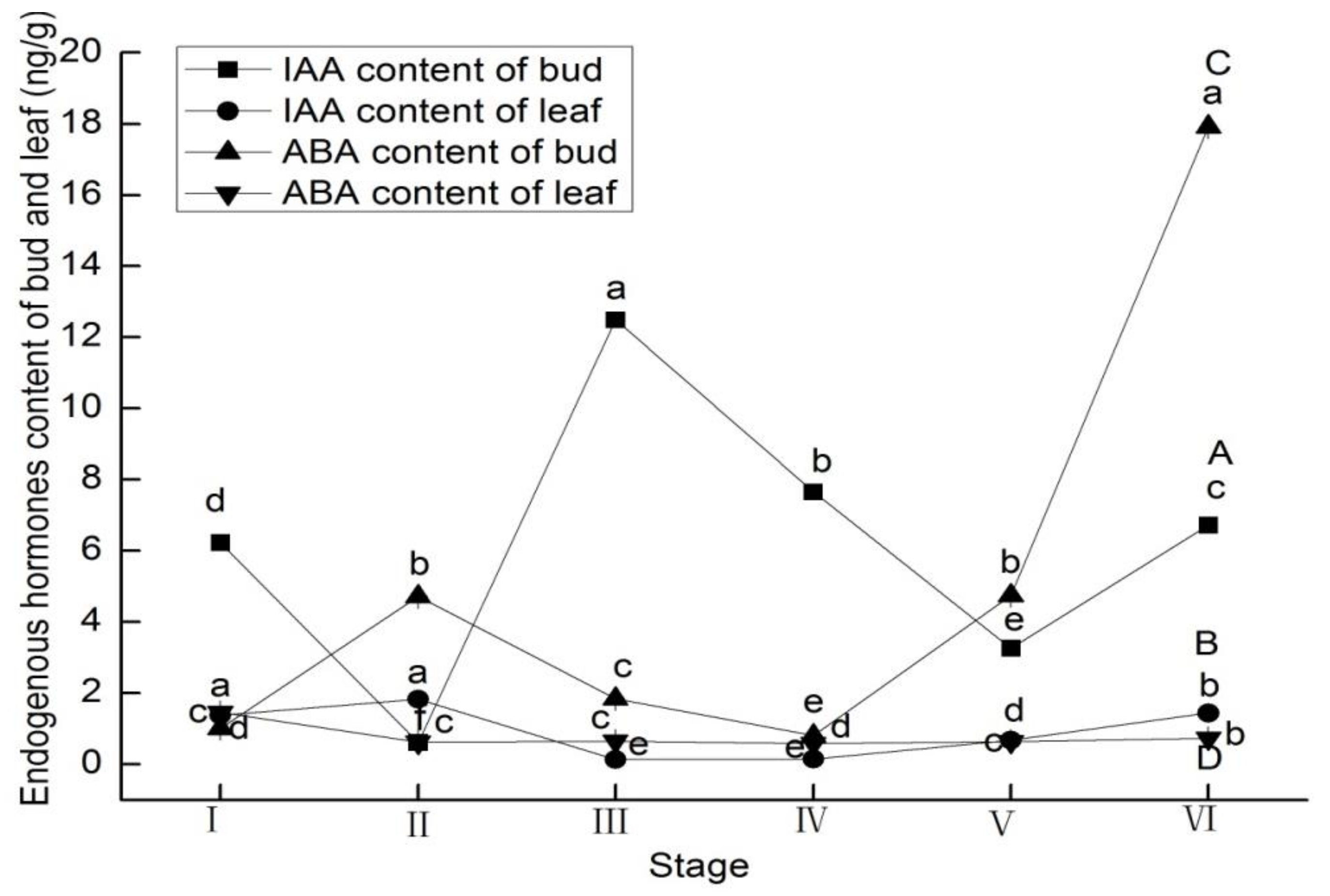
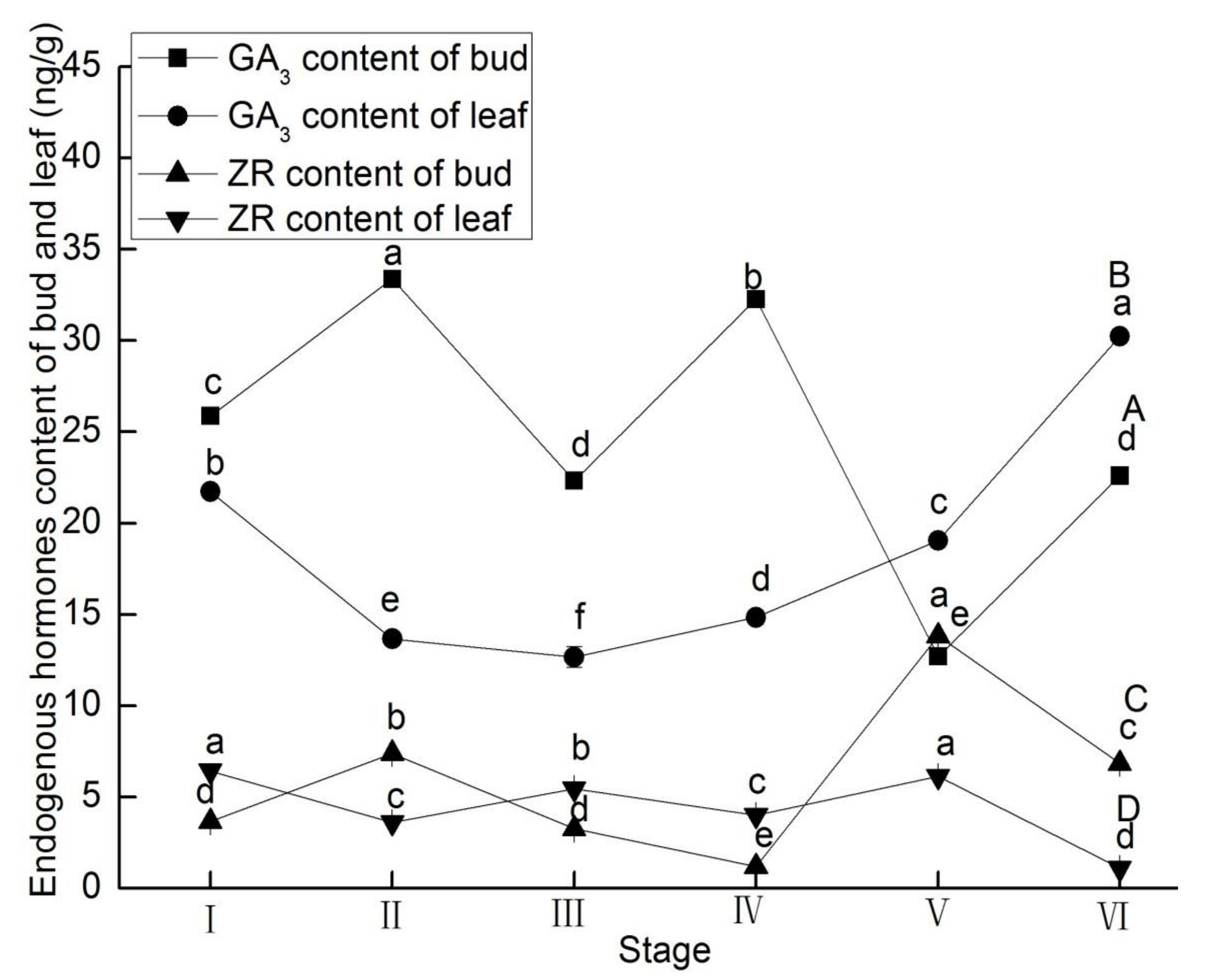
3.1. Endogenous Hormones Content
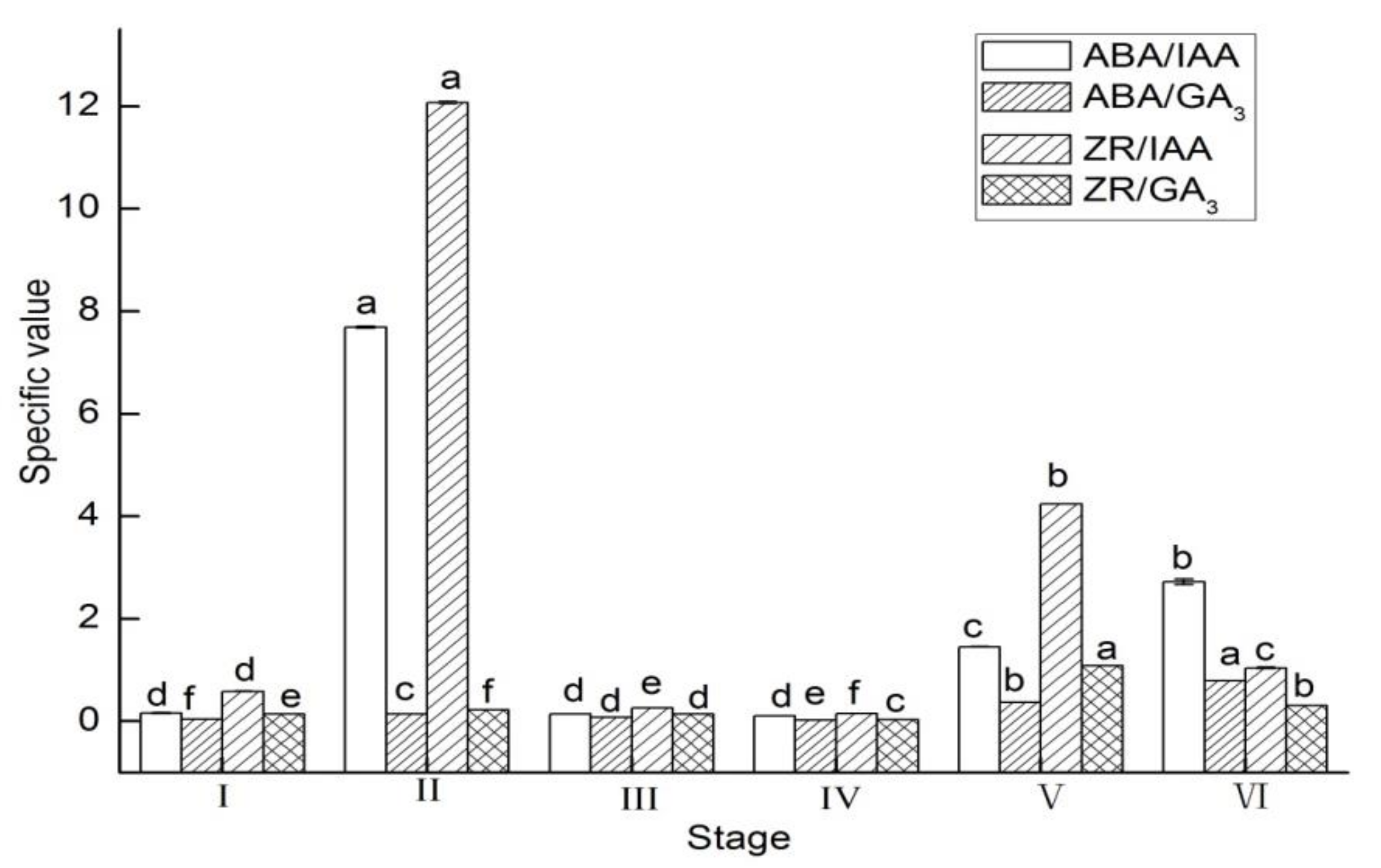
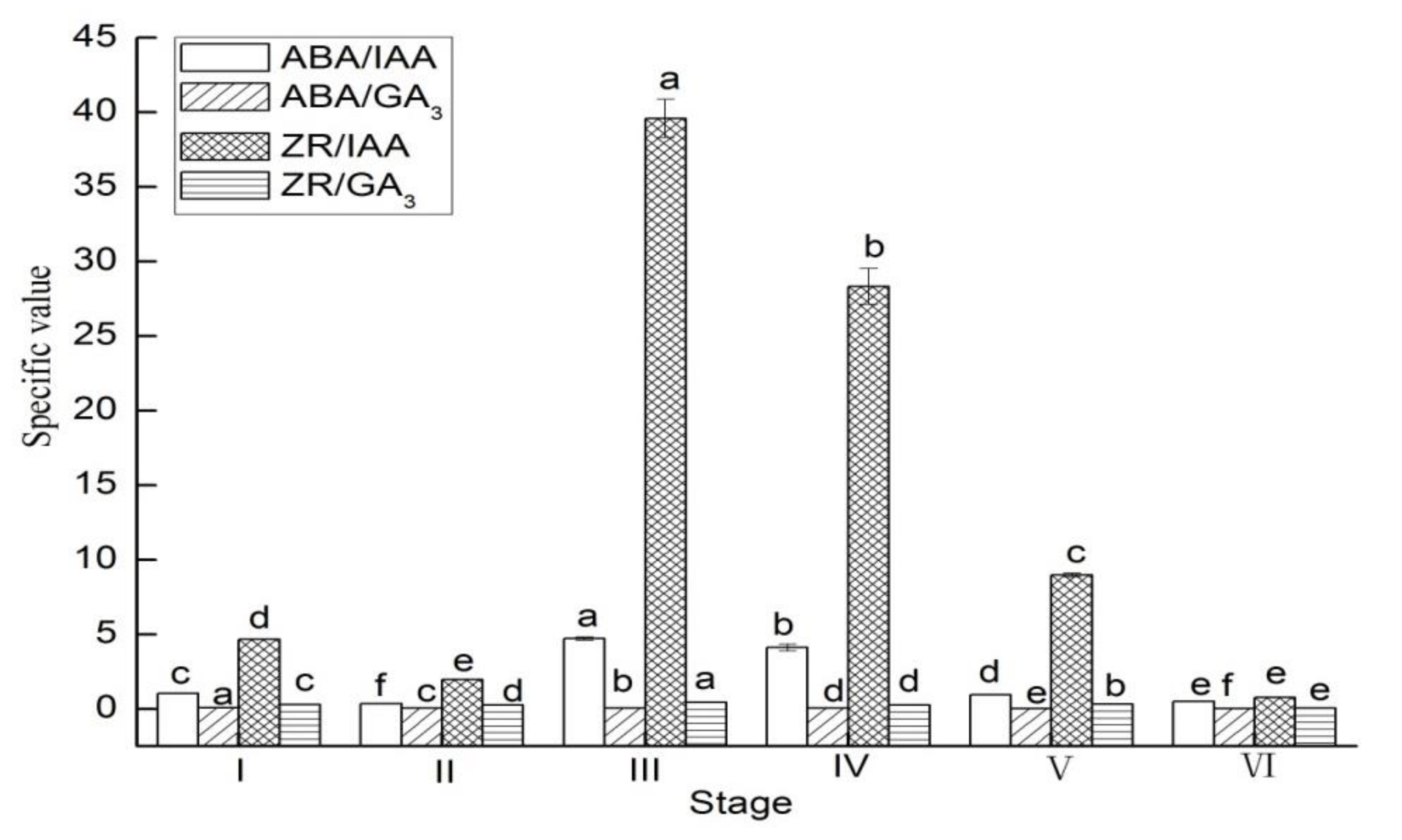
3.2. Endogenous Hormones Specific Value
3.3. Carbohydrates
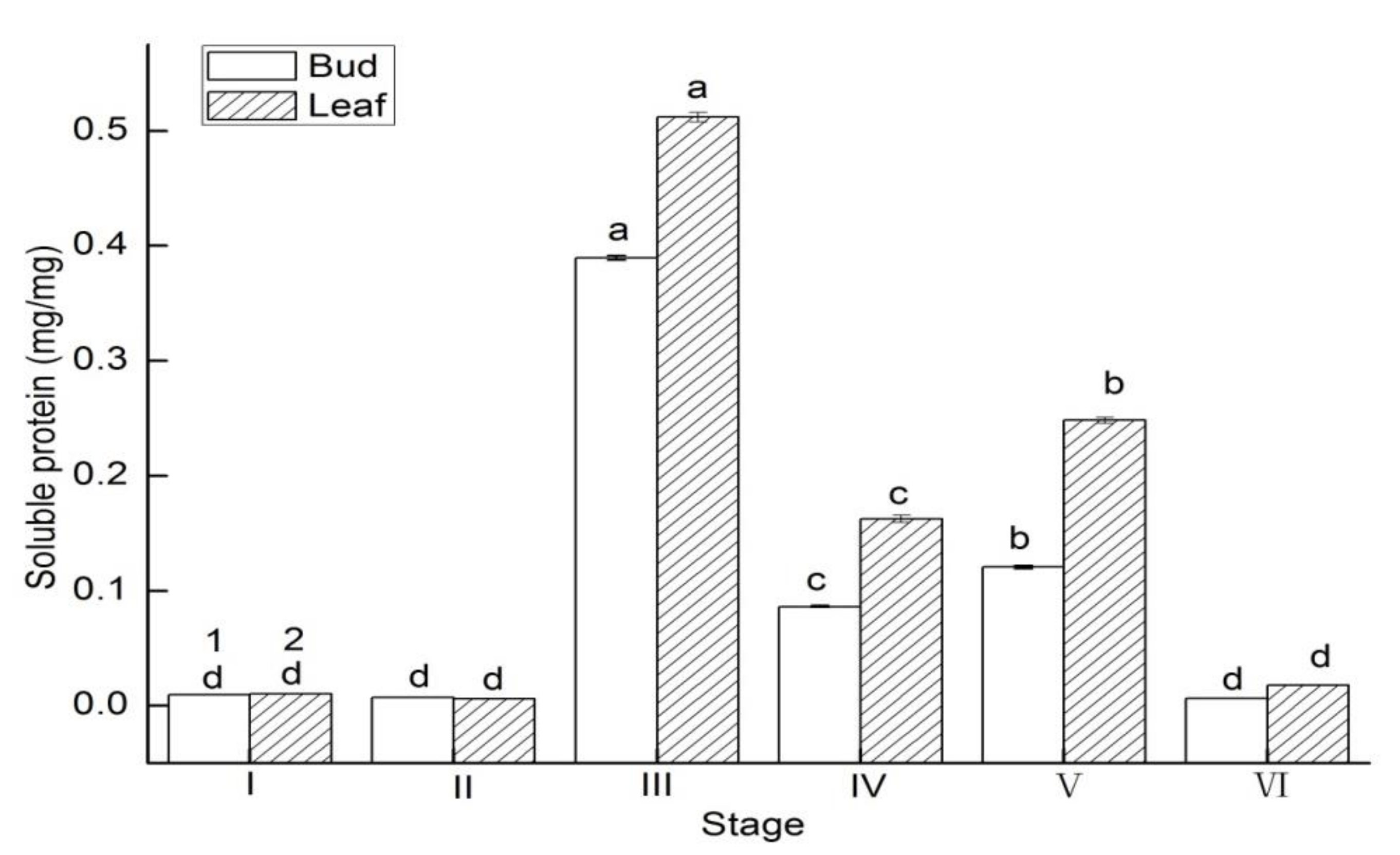
3.4. Soluble Protein
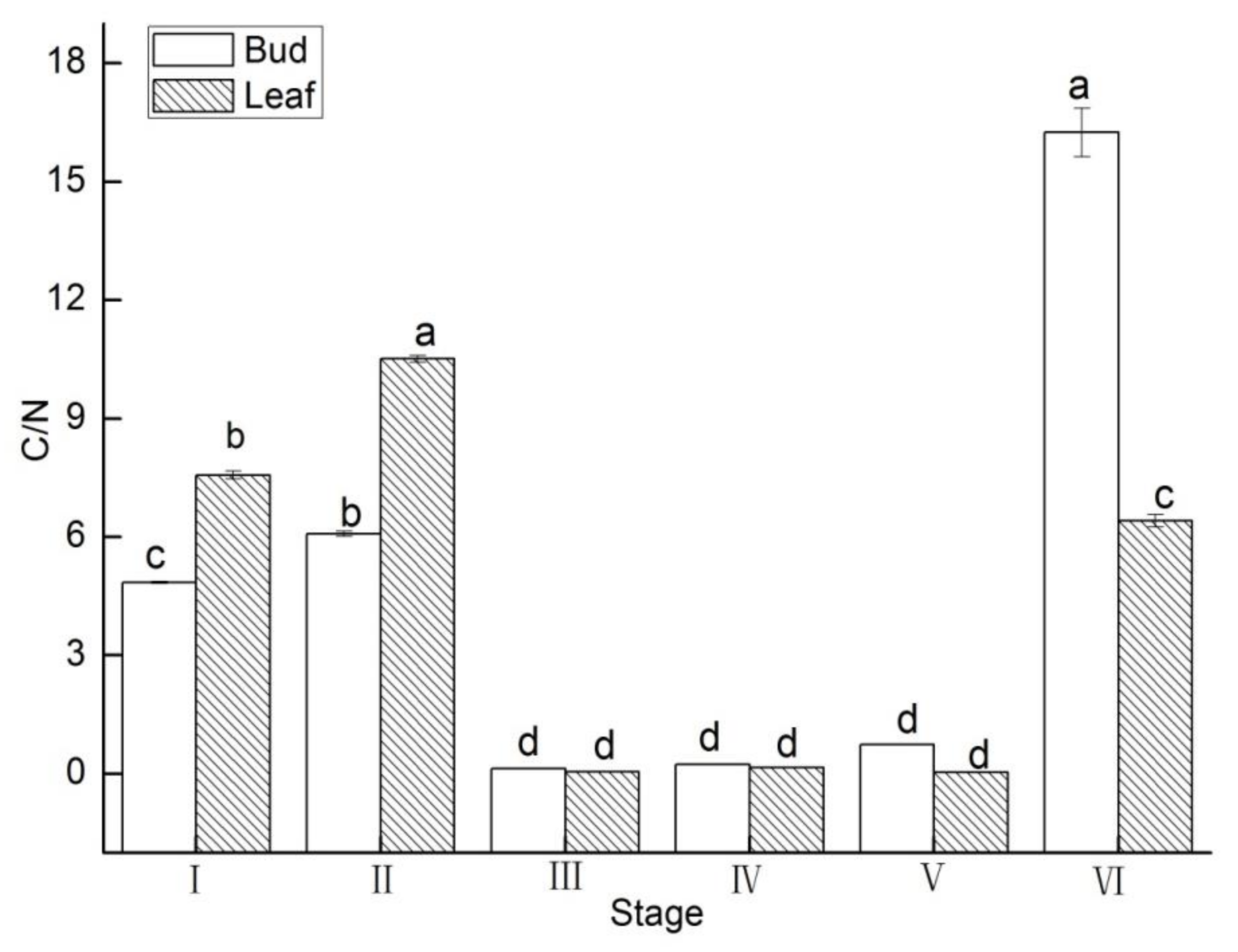
3.5. C/N Value
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zheng, J.; Ding, C.-X.; Wang, L.-S.; Li, G.-L.; Shi, J.-Y.; Li, H.; Wang, H.-L.; Suo, Y.-R. Anthocyanins composition and antioxidant activity of wild Lycium ruthenicum Murr. from Qinghai-Tibet Plateau. Food Chem. 2011, 126, 859–865. [Google Scholar] [CrossRef]
- Wang, J.-H.; Chen, W. Responses of seed germination and seedling growth of Lycium ruthenicum to salt stress. Chin. J. Ecol. 2012, 31, 804–810. [Google Scholar]
- Mert, C.; Barut, E.; İpek, A. Variation in flower bud differentiation and progression of floral organs with respect to crop load in olive. Not. Bot. Horti Agrobot. Clujnapoca 2013, 41, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Riboni, M.; Robustelli, T.-A.; Galblati, M.; Tonelli, C.; Cont, L. ABA-dependent control of gigantea signalling enables drought escape via up-regulation of flowering locust in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6309–6322. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.-Y.; Zhang, Q.-M.; Wu, S. Advances in research on the mechanism of flower-bud differentiation of fruit trees. J. Fruit Sci. 2003, 20, 345–350. [Google Scholar]
- Wyatt, R. Pollinator plant interactions and the evolution of breeding system. In Pollination Biology; Real, L., Ed.; Academic Press: New York, NY, USA, 1983. [Google Scholar]
- Ulger, S.; Sonmez, S.; Karkacier, M.; Ertoy, N.; Akdesir, O.; Aksu, M. Determination of endogenous hormones, sugars and mineral nutrition levels during the induction, initiation and differentiation stage and their effects on flower formation in olive. Plant Growth Regul. 2004, 42, 89–95. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Li, W.-Q.; Liu, X.-H.; Lu, Y.-M. Changes in the morphology of the apical meristem and the levels of endogenous hormones during flower bud development of different Lily (Lilium) cultivars. Philipp. Agric. Sci. 2013, 96, 296–300. [Google Scholar]
- Shaziya, N.; Inayatullah, T.; Syed, S.-A.; Riyaz, A.D. Physiological and biochemical aspects of flower development and senescence in Nicotiana plumbaginifolia Viv. Folia Hort. 2017, 29, 25–31. [Google Scholar]
- Abreu, M.-E.; Munne, B.-S. Salicylic acid deficiency in NahG transgenic lines and sid2 mutants increases seed yield in the annual plant Arabidopsis thaliana. J. Exp. Bot. 2009, 60, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- Nazari, M.; Zarinkamar, F.; Soltani, B.-m. Physiological, biochemical and molecular responses of Mentha aquatica L. to manganese. Plant Physiol. Biochem. 2017, 120, 202–212. [Google Scholar] [CrossRef]
- Robyt, J.-F.; White, B.-J. Protein Purification: Principles and Practices, Biochemical Techniques, Theory and Practice; Cole Publ.: Belmont, CA, USA, 1987; pp. 40–72. [Google Scholar]
- Yan, F. Study on Morphological Differentiation of Flower Bud of Hippeastrum; Northwest Agriculture and Forestry University: Xianyang, China, 2009. [Google Scholar]
- Bradford, M.-M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Ann. Biochem. 1976, 72, 248–253. [Google Scholar] [CrossRef]
- Anton, J.-M.; Will, G.; Gerard, W.-M.; George, J.-W. In vitro flower bud formation in tobacco: Interaction of hormones. Plant Physiol. 1991, 97, 402–408. [Google Scholar]
- Davis, S.-J. Integrating hormones into the floral-transition pathway of Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1201–1210. [Google Scholar] [CrossRef]
- Shalit, A.; Rozman, A.; Alvarez, J.-P.; Bowman, J.-L.; Eshed, Y.; Lifschitz, E. The flowering hormone florigen functions as a general systemic regulator of growth and termination. Proc. Natl. Acad. Sci. USA 2009, 106, 8392–8397. [Google Scholar] [CrossRef] [Green Version]
- Banasik, L.; Saniewski, M. The effect of different auxins on tulip stem elongation. Acta Hortic. 1985, 167, 193–204. [Google Scholar] [CrossRef]
- Wan, C.-Y.; Mi, L.; Chen, B.-Y.; Li, J.-F.; Huo, H.-Z.; Xu, J.-T.; Chen, X.-P. Effects of nitrogen during nursery stage on flower bud differentiation and early harvest after transplanting in strawberry. Braz. J. Bot. 2018, 41, 1–10. [Google Scholar] [CrossRef]
- Feng, F.; Yang, J.-S. Relationship between floral bud differentiation and endogenous hormones in autumn-cutting Chrysanthemum morifolium Jinba. Sci. Agric. Sin. 2011, 44, 552–561. [Google Scholar]
- Hao, Y. The Study of the Endogenesis Hormone and Dynamic Growth of Apricot Varieties during Florescence and Fruit Period; North West Agriculture and Forestry University: Xianyang, China, 2009. [Google Scholar]
- Luckwill, L.-C. Proceedings of the XLX International Horticultural Congress; International Society for Horticultural Science: Vienna, Austria, 1974; Volume 11, pp. 169–177. [Google Scholar]
- Wu, Z.-X.; Zhou, Z.D.; Tao, Z.-L.; Wang, L.-X. Change of endogenous hormones in Feizixiao and Edan litchi during flower bud differentiation. Chin. J. Trop. Crops 2005, 26, 42–45. [Google Scholar]
- Laia, A.; Sergi, M.-B. Hormonal changes during flower development in floral tissues of Lilium. Planta 2012, 236, 343–356. [Google Scholar]
- Niu, H.-L.; Zhang, H.-W.; Bian, Y.; Li, X.-G. Flower Formation and Endogenous Hormones Dynamic in Chinese Jujube. Acta Hortic. Sin. 2015, 42, 655–664. [Google Scholar]
- Wang, Y.H.; Fan, C.-H.; Shen, X.; Qu, G.-M.; Shi, J.D. Changes in endogenous hormones during the flower bud differentiation of sweet cherry. Acta Agric. Boreali-Occident. Sin. 2002, 1, 64–67. [Google Scholar]
- Gao, X.-J.; Wu, X.-E.; Wang, S.-Y.; Chen, C.-L.; Cheng, J.-H.; Dong, G.-P.; Peng, L. Change in endogenous hormone contents of mango during floral differentiation after heading-back. Fujian J. Agri. Sci. 2009, 24, 227–230. [Google Scholar]
- Ai, X.-M.; He, R.-Y.; Hu, Y.-F. Flower bud differentiation and their relationships with cntent changes of endogenous hormones in potatoes. Acta Bot. Boreali-Occident. Sin. 2018, 38, 87–94. [Google Scholar]
- Wang, L.; Tang, G.-G.; Liu, T. Variation of endogenous hormone and nucleic acid content during flower bud differentiation in Lycoris radiata. J. Nanjing For. Univ. 2008, 32, 67–70. [Google Scholar]
- Priestly, G.-A. The annual turnover resources in young olive trees. J. Hortic. Sci. 1977, 52, 105–112. [Google Scholar] [CrossRef]
- Sarmiento, R.; Valpuestra, V.; Catalina, L.; Gonzoles, G.-F. Variation of the contents of starch and soluble carbohydrate of leaves and buds of plants of Olea europaea var Manzanillo in relation to their vegetative or reproductive process. Aneles Edafol. Agrobiol. 1976, 35, 683–695. [Google Scholar]
- Macnish, A.-J.; Jiang, C.-Z.; Negre, Z.-F.; Reid, M.-S. Physiological and molecular changes during opening and senescence of Nicotiana mutabilis flowers. Plant Sci. 2010, 179, 267–272. [Google Scholar] [CrossRef]
- Doorn, W.-G.; Woltering, E.-J. Physiology and molecular biology of petal senescence. J. Exp. Bot. 2008, 59, 453–480. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Soluble Sugar (mg/mg) | Sucrose (mg/mg) | Starch (mg/mg) | |||
|---|---|---|---|---|---|---|
| Bud | Leaf | Bud | Leaf | Bud | Leaf | |
| I | 0.047 ± 0.00 d | 0.078 ± 0.00 b | 0.021 ± 0.00 e | 0.078 ± 0.00 e | 0.012 ± 0.00 e | 0.017 ± 0.00 d |
| II | 0.044 ± 0.00 d | 0.064 ± 0.00 c | 0.024 ± 0.00 d | 0.064 ± 0.00 f | 0.009 ± 0.00 f | 0.011 ± 0.00 e |
| III | 0.051 ± 0.00 c | 0.026 ± 0.00 d | 0.070 ± 0.00 a | 0.134 ± 0.00 a | 0.207 ± 0.00 a | 0.075 ± 0.00 b |
| IV | 0.020 ± 0.00 e | 0.026 ± 0.00 d | 0.047 ± 0.00 b | 0.121 ± 0.00 b | 0.095 ± 0.00 c | 0.045 ± 0.00 c |
| V | 0.090 ± 0.00 b | 0.011 ± 0.00 e | 0.047 ± 0.00 b | 0.083 ± 0.00 d | 0.106 ± 0.00 b | 0.101 ± 0.00 a |
| VI | 0.103 ± 0.00 a | 0.113 ± 0.00 a | 0.031 ± 0.00 c | 0.113 ± 0.00 c | 0.024 ± 0.00 d | 0.015 ± 0.00 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; An, L.; Yu, H.; Yang, M. Endogenous Hormones and Biochemical Changes during Flower Development and Florescence in the Buds and Leaves of Lycium ruthenicum Murr. Forests 2022, 13, 763. https://doi.org/10.3390/f13050763
Guo Y, An L, Yu H, Yang M. Endogenous Hormones and Biochemical Changes during Flower Development and Florescence in the Buds and Leaves of Lycium ruthenicum Murr. Forests. 2022; 13(5):763. https://doi.org/10.3390/f13050763
Chicago/Turabian StyleGuo, Youyan, Lizhe An, Hongyuan Yu, and Miaomiao Yang. 2022. "Endogenous Hormones and Biochemical Changes during Flower Development and Florescence in the Buds and Leaves of Lycium ruthenicum Murr" Forests 13, no. 5: 763. https://doi.org/10.3390/f13050763
APA StyleGuo, Y., An, L., Yu, H., & Yang, M. (2022). Endogenous Hormones and Biochemical Changes during Flower Development and Florescence in the Buds and Leaves of Lycium ruthenicum Murr. Forests, 13(5), 763. https://doi.org/10.3390/f13050763