
Discovery and Biology of Spathius verustus Chao (Hymenoptera: Braconidae), a Potential Biological Agent on Two Monochamus Vectors of the Pinewood Nematode

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sentinel Log Preparation
2.2. Survey Site
2.3. Field Survey
2.4. Rearing and Identification of the Parasitoid
2.5. Indoor Oviposition Observation for the Parasitoid
2.6. Data Analysis
2.7. Depository
3. Results
3.1. Taxonomic Account of Spathius verustus
3.2. Biology of Spathius verustus
3.3. Indoor Oviposition Observation of S. verustus
3.4. Parasitism Rates of S. verustus from the Field Surveys in 2016 and 2017
4. Discussion
4.1. Indoor Oviposition Observation of S. verustus
4.2. The Ecological Relationship between the Host and S. verustus
4.3. In Spatial Effect of Parasitism Rate of S. verustus According to Environmental Variables
4.4. Possibility of S. verustus as a Biological Control Agent on the Insect Vectors of PWN
4.5. Compensate the Defect and Future Study
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, W.I.; Park, Y.S. Dispersal patterns of exotic forest pests in South Korea. Insect Sci. 2012, 19, 535–548. [Google Scholar] [CrossRef]
- Dwinell, L.D. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Enda, N. The damage of pine wilt disease in Taiwan. For. Pests 1988, 37, 161–166. [Google Scholar]
- Malek, R.B.; Appleby, J.E. Epidemiology of pine wilt in Illinois. Disease distribution. Plant Dis. 1984, 68, 180–186. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar] [PubMed]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Zamora, P.; Rodríguez, V.; Renedo, F.; Sanz, A.V.; Domínguez, J.C.; Pérez-Escolar, G.; Miranda, J.; Álvarez, B.; González-Casas, A.; Mayor, E.; et al. First report of Bursaphelenchus xylophilus causing pine wilt disease on Pinus radiata in Spain. Plant Dis. 2015, 99, 1449. [Google Scholar] [CrossRef]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 18–25. [Google Scholar]
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.I.; Chang, K.H. First finding of the pinewood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) nickle and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. [Google Scholar]
- Tóth, Á. Bursaphelenchus xylophilus, the pinewood nematode: Its significance and a historical review. Acta Biol. Szegediensis 2011, 55, 213–217. [Google Scholar]
- Shin, S.C. Pine wilt disease in Korea. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 32–36. [Google Scholar]
- Mamiya, Y.; Enda, N. Transmission of Bursaphelenchus lignicolus (nematoda: Aphelenchoididae) by Monochamus alternatus (coleoptera: Cerambycidae). Nematologica 1972, 18, 159–162. [Google Scholar] [CrossRef]
- Linit, M.J. Nemtaode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227–235. [Google Scholar] [PubMed]
- Sousa, E.; Bravo, M.A.; Pires, J.; Naves, P.; Penas, A.C.; Bonifácio, L.; Mota, M.M. Bursaphelenchus xylophilus (nematoda; aphelenchoididae) associated with Monochamus galloprovincialis (coleoptera; Cerambycidae) in Portugal. Nematology 2001, 3, 89–91. [Google Scholar]
- National Institute of Biological Resources. National Species List of Korea. III. Insects (Hexapoda); Designzip: Seoul, Korea, 2019; Volume 988. [Google Scholar]
- Kwon, T.S.; Lim, J.H.; Sim, S.J.; Kwon, Y.D.; Son, S.; Lee, K.Y.; Kim, Y.T.; Shin, C.H.; Ryu, S.B.; Lee, C.K.; et al. Distribution patterns of Monochamus alternatus and M. saltuarius (Coleoptera: Cerambycidae) in Korea. J. Korean For. Soc. 2006, 95, 543–550. [Google Scholar]
- Kwon, S.H.; Go, M.S.; Park, J.; Han, T.W.; Kim, K.B.; Shin, C.H.; Kim, D.S. The bimodal adult activity of Monochamus alternatus (coleoptera: Cerambycidae) caught in pheromone traps in Jeju can be explained by the competitive attractiveness of dying pine trees. Entomol. Res. 2019, 49, 172–178. [Google Scholar] [CrossRef]
- Choi, W.I.; Jeon, M.J.; Park, Y.S. Structural dynamics in the host-parasitoid system of the pine needle gall midge (Thecodiplosis japonensis) during invasion. PeerJ 2017, 5, e3610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T.S.; Shin, J.H.; Lim, J.H.; Kim, Y.K.; Lee, E.J. Management of Pine Wilt Disease in Korea Through Preventative Silvicultural Control. Forest Ecol. Manag. 2011, 261, 562–569. [Google Scholar] [CrossRef]
- Suh, D.Y.; Jung, J.K.; Lee, S.K.; Seo, S.T. Effect of aerial spraying of thiacloprid on pine sawyer beetles (Monochamus alternatus) and honey bees (Apis mellifera) in pine forests. Entomol. Res. 2021, 51, 83–89. [Google Scholar] [CrossRef]
- Kim, J.B.; Kim, D.Y.; Park, N.C. Development of an aerial precision forecasting techniques for the pine wilt disease damaged area based on GIS and GPS. J. Korean Assoc. Geogr. Inf. 2010, 13, 28–34. [Google Scholar] [CrossRef]
- Park, Y.S.; Park, Y.K.; Yang, H.M. Effects of clear-cutting on forest arthropod communities at two different vertical levels (crown and ground surface). Korean J. Ecol. Environ. 2016, 49, 271–278. [Google Scholar] [CrossRef]
- Jeon, K.S.; Kim, C.S.; Park, N.C.; Hur, T.C.; Hong, S.C. Effects on control of pine wilt disease (Bursaphelenchus xylophilus) by thinning methods in red pine (Pinus densiflora) forest. J. Korean For. Soc. 2011, 100, 165–171. [Google Scholar]
- Wang, Y.; Chen, F.; Wang, L.; Li, M. Investigation of beetle species that carry the pinewood nematode, Bursaphelenchus xylophilus (Steiner and Buhrer) nickle, in China. J. For. Res. 2020, 32, 1745–1751. [Google Scholar] [CrossRef]
- Li, L.; Wei, W.; Liu, Z.; Sun, J. Host Adaptation of a gregarious parasitoid Sclerodermus harmandi in artificial rearing. BioControl. 2010, 55, 465–472. [Google Scholar] [CrossRef]
- Tian, S.P.; Xu, Z.Q. Effects of different temperatures on the development of Scleroderma guani reared with Tenebrio molitor. Entomol. Knowl. 2003, 40, 356–359. [Google Scholar]
- Morimoto, K.; Iwasaki, A. Role of Monochamus alternatus (Coleoptera: Cerambycidae) as a vector of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae). J. Jpn. For. Soc. 1972, 54, 177–183. [Google Scholar] [CrossRef]
- Takizawa, Y.; Shoji, T. Distribution of Monochamus saltuarius gebler, and its possible transmission of pinewood nematodes in Iwate prefecture. For. Pests 1982, 31, 4–6. [Google Scholar]
- Kobayashi, F.; Yamane, A.; Ikeda, T. The Japanese pine sawyer beetle as the vector of pine wilt disease. Annu. Rev. Entomol. 1984, 29, 115–135. [Google Scholar] [CrossRef]
- Togashi, K. A Field experiment on dispersal of newly emerged adults of Monochamus alternatus (coleoptera: Cerambycidae). Res. Popul. Ecol. 1990, 32, 1–13. [Google Scholar] [CrossRef]
- Hong, J.I.; Koh, S.H.; Chung, Y.J.; Shin, S.C.; Kim, G.H.; Choi, K.S. Biological characteristics of Sclerodermus harmandi (Hymenoptera: Bethylidae) parasitized on cerambycid. Korean J. Appl. Entomol. 2008, 47, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Sin, S.C. Biological control of tree insect pest. Landscaping Tree 2012, 129, 28–34. [Google Scholar]
- Choi, W.I.; Nam, Y.; Lee, C.Y.; Choi, B.K.; Shin, Y.J.; Lim, J.H.; Koh, S.H.; Park, Y.S. Changes in major insect pests of pine forests in Korea over the last 50 years. Forests 2019, 10, 692. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.O.; Lyu, D.P.; Choi, G.S.; Jeong, Y.J.; Shin, S.C.; Lee, S.H. A taxonomic note on Sclerodermus harmandi, ectoparasite of stem and wood boring insect larvae (Hymenoptera: Chrysidoidea: Bethylidae) in South Korea. J. Asia Pac. Entomol. 2006, 9, 115–119. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, B.; Xie, Y.; Yang, S. Potential non-target effects of Sclerodermus harmandi (Hymenoptera: Bethylidae) on Triaspis sp. (Hymenoptera: Braconidae). Biocontrol Sci. Technol. 2017, 27, 909–917. [Google Scholar] [CrossRef]
- Kim, J.K.; Won, D.S.; Park, Y.C.; Koh, S.H. Natural Enemies of Wood Borers and Seasonal Occurrence of Major Natural Enemies of Monochamus saltuarius on Pine Trees. J. Korean Soc. For. Sci. 2010, 99, 439–445. [Google Scholar]
- Petersen-Silva, R.; Pujade-Villar, J.; Naves, P.; Sousa, E.; Belokobylskij, S. Parasitoids of Monochamus galloprovincialis (Coleoptera, Cerambycidae), vector of the pinewood nematode, with identification key for the palaearctic region. ZooKeys 2012, 251, 29. [Google Scholar] [CrossRef]
- Brabbs, T.; Collins, D.; Hérard, F.; Maspero, M.; Eyre, D. Prospects for the use of biological control agents against Anoplophora in Europe. Pest Manag. Sci. 2015, 71, 7–14. [Google Scholar] [CrossRef]
- Duan, J.J.; Aparicio, E.; Tatman, D.; Smith, M.T.; Luster, D.G. Potential new associations of north american parasitoids with the invasive asian longhorned beetle (Coleoptera: Cerambycidae) for biological control. J. Econ. Entomol. 2016, 109, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Hérard, F.; Bon, M.C.; Maspero, M.; Cocquempot, C.; Lopez, J. Survey and evaluation of potential natural enemies of Anoplophora glabripennis and A. chinensis. In Proceedings of the 16th US Department of Agriculture Interagency Research Forum on Gypsy Moth and Other Invasive Species, Newton Square, PA, USA, 18–21 January 2005; pp. 18–21. [Google Scholar]
- Wang, X.; Aparicio, E.M.; Murphy, T.C.; Duan, J.J.; Elkinton, J.S.; Gould, J.R. Assessing the host range of the north american parasitoid Ontsira mellipes: Potential for biological control of asian longhorned beetle. Biol. Control 2019, 137, 104028. [Google Scholar] [CrossRef]
- Delvare, G.; Bon, M.C.; Hérard, F.; Cocquempot, C.; Maspero, M.; Colombo, M. Description of Aprostocetus anoplophorae n. sp. (Hymenoptera: Eulophidae), a new egg parasitoid of the invasive pest Anoplophora chinensis (Förster) (Coleoptera: Cerambycidae). Ann. Société Entomol. Fr. 2004, 40, 227–233. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, H.L.; Ku, D.S.; Hérard, F.; Gould, J.R.; Williams, D.W.; Kim, I.K.; Hong, K.J. Discovery of Spathius ibarakius Belokobylskij et Maeto (Hymenoptera: Braconidae) as a larval ectoparasitoid of citrus longhorned beetle in Korea. Korean J. Appl. Entomol. 2016, 55, 285–291. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Strazanac, J.S.; Marsh, P.M.; Van Achterberg, C.; Choi, W.Y. First Recorded Parasitoid from China of Agrilus planipennis: A New Species of Spathius (Hymenoptera: Braconidae: Doryctinae). Ann. Entomol. Soc. Am. 2005, 98, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.J.; Yang, Z.Q. Studies on biological control of Anoplophora glabripennis (Coleoptera: Cerambycidae) with a parasitoid, Sclerodermus guani (Hymenoptera: Bethylidae). J. Environ. Entomol. 2008, 30, 127–134. [Google Scholar]
- Abell, K.J.; Duan, J.J.; Shrewsbury, P.M. Determining Optimal parasitoid release timing for the biological control of emerald ash borer (Coleoptera: Buprestidae). Fla. Entomol. 2020, 102, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Gould, J.S.; Bauer, L.S.; Lelito, J.; Duan, J. Emerald Ash Borer Biological Control Release and Recovery Guidelines; US Department of Agriculture. Animal Plant Health Inspection Service; Forest Service Northern Research Station and Agricultural Research Service: Riverdale, MD, USA, 2012; pp. 1–76. [Google Scholar]
- Gould, J.R.; Bauer, L.S.; Duan, J.J.; Williams, D.; Liu, H. History of emerald ash borer biological control. In Biology and Control of Emerald Ash Borer. FHTET-2014-09; Van Driesche, R.G., Reardon, R.C., Eds.; United States Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2015; Volume 5, pp. 83–95. [Google Scholar]
- Wang, X.; Wang, X.Y.; Kenis, M.; Cao, L.M.; Duan, J.J.; Gould, J.R.; Hoelmer, K.A. Exploring the potential for novel associations of generalist parasitoids for biological control of invasive woodboring beetles. BioControl 2021, 66, 97–112. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Hansen, J.A.; Abell, K.J.; Van Driesche, R. An improved method for monitoring parasitism and establishment of Oobius Agrili (Hymenoptera: Encyrtidae), an egg parasitoid introduced for biological control of the emerald ash borer (Coleoptera: Buprestidae) in North America. Biol. Control 2012, 60, 255–261. [Google Scholar] [CrossRef]
- Abell, K.J.; Bauer, L.S.; Miller, D.L.; Duan, J.J.; Van Driesche, R.G.V. Monitoring the establishment and flight phenology of parasitoids of emerald ash borer (Coleoptera: Buprestidae) in Michigan by using sentinel eggs and larvae. Fla. Entomol. 2016, 99, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Futai, K.; Shirakikawa, S.; Nakai, I. The suitability of korean pine (Pinus koraiensis Sieb. et Zucc.) and japanese red pine (P. densiflora Sieb. et Zucc.) as a host of the japanese pine sawyer, Monochamus alternatus hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1994, 29, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.C.; Kim, J.H.; Park, J.B.; Shin, S.C.; Chung, Y.J.; Cho, S.; Park, Y.C. Growth, development, and reproduction of Monochamus saltuarius (Coleoptera: Cerambycidae) on conifers fed to larvae. Korean J. Appl. Entomol. 2008, 47, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Ochi, K. Ecological studies on cerambycid injurious to pine trees (II) biology of two monochamus (Coleoptera, Cerambycidae). J. Jpn. For. Soc. 1969, 51, 188–192. [Google Scholar] [CrossRef]
- Han, J.H.; Yoon, C.; Shin, S.C.; Kim, G.H. Seasonal occurrence and morphological measurements of pine sawyer, Monochamus saltuarius adults (Coleoptera: Cerambycidae). J. Asia Pac. Entomol. 2007, 10, 63–67. [Google Scholar] [CrossRef]
- Tang, P.; Belokobylskij, S.; Chen, X.X. Spathius Nees, 1818 (Hymenoptera: Braconidae, Doryctinae) from China with a key to species. Zootaxa 2015, 3960, 1–132. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, C.E. A survey of identified kairomones and synomones used by insect parasitoids to locate and accept their hosts. Chemoecology 1996, 7, 121–131. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yang, Z.Q.; Gould, J.R.; Wu, H.; Ma, J.H. Host-seeking behavior and parasitism by Spathius agrili yang (Hymenoptera: Braconidae), A parasitoid of the emerald ash borer. Biol. Control 2010, 52, 24–29. [Google Scholar] [CrossRef]
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.0. Available online: http://CRAN.R-project.org/package=rstatix (accessed on 14 May 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Veinna, Asutria, 2021; Available online: https://www.R-project.org/ (accessed on 14 May 2022).
- Douglas, B.; Martin, M.; Ben, B.; Steve, W. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Murdoch, W.W.; Reeve, J.D.; Huffaker, C.B.; Kennett, C.E. Biological control of olive scale and its relevance to ecological theory. Am. Nat. 1984, 123, 371–392. [Google Scholar] [CrossRef]
- Ku, D.S.; Belokobylskij, S.A.; Cha, J.Y. Family Braconidae. Economic insects of Korea 16. Insecta Koreana 2001, 23, 1–281. [Google Scholar]
- Lee, H.R.; An, T.H.; Ku, D.S.; Byun, B.K. A newly recorded species of the genus Spathius nees (Hymenoptera: Braconidae: Doryctinae) with a checklist in Korea. Korean J. Appl. Entomol. 2017, 56, 295–299. [Google Scholar] [CrossRef]
- Belokobylskij, S.A.; Maeto, K. Doryctinae (Hymenoptera: Braconidae) of Japan; Natura Optima Dux Foundation: Warszawa, Poland, 2009; pp. 1–806. [Google Scholar]
- Kishi, Y. Pine Wood Nematode and the Japanese Pine Sawyer; Thomas Company Limited: Tokyo, Japan, 1995; pp. 1–302. [Google Scholar]
- Go, M.S.; Kwon, S.H.; Kim, S.B.; Kim, D.S. The developmental characteristics for the head capsule width of Monochamus alternatus (Coleoptera: Cerambycidae) larvae and determination of the number of instars. J. Insect Sci. 2019, 19, 26. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Dai, Y.; Wang, Y.; Wang, L.; Sun, S.; Chen, F. New insights in to the life history of Monochamus saltuarius (Cerambycidae: Coleoptera) can enhance surveillance strategies for pine wilt disease. J. For. Res. 2021, 32, 2699–2707. [Google Scholar] [CrossRef]
- Gonçalves, E.; Figueiredo, A.C.; Barroso, J.G.; Millar, J.G.; Henriques, J.; Sousa, E.; Bonifácio, L. Characterization of cuticular compounds of the cerambycid beetles Monochamus galloprovincialis, Arhopalus syriacus, and Pogonocherus perroudi, potential vectors of pinewood nematode. Entomol. Exp. Appl. 2021, 169, 183–194. [Google Scholar] [CrossRef]
- Häckermann, J.; Rott, A.S.; Dorn, S. How two different host species influence the performance of a gregarious parasitoid: Host size is not equal to host quality. J. Anim. Ecol. 2007, 76, 376–383. [Google Scholar] [CrossRef]
- Nicol, C.M.Y.; Mackauer, M. The scaling of body size and mass in a host-parasitoid association: Influence of host species and stage. Entomol. Exp. Appl. 1999, 90, 83–92. [Google Scholar] [CrossRef]
- Jamil, A.; Ul Abdin, Z.; Arshad, M.; Falabella, P.; Abbas, S.K.; Tahir, M.; Jamil, A.; Manzoor, A.; Shaina, H. Multiple effects of host density on egg density and the sex ratio of progeny of Bracon hebetor (Say.) (Hymenoptera: Braconidae). Pak. J. Zool. 2015, 47, 455–460. [Google Scholar]
- Yu, S.-H.; Ryoo, M.I.; Na, J.H.; Choi, W.I. Effect of host density on egg dispersion and the sex ratio of progeny of Bracon hebetor (Hymenoptera: Braconidae). J. Stored Prod. Res. 2003, 39, 385–393. [Google Scholar] [CrossRef]
- Furlong, M.J.; Zalucki, M.P. Climate change and biological control: The consequences of increasing temperatures on host–parasitoid interactions. Curr. Opin. Insect Sci. 2017, 20, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.J.; Choi, K.S.; Lee, S.M.; Kim, D.S.; Lee, S.G.; Park, J.G. Measurement and within-tree distribution of larval entrance and adult emergence holes of japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Korean J. Appl. Entomol. 2003, 42, 315–321. [Google Scholar]
- Ma, T.; Shi, X.; Shen, J.; Wang, C.; Zhang, S.; Lu, X.; Sun, Z.; Chen, X.; Wang, C.; Xie, W.; et al. Field evaluation of commercial attractants and trap placement for monitoring pine sawyer beetle, Monochamus alternatus (Coleoptera: Cerambycidae) in Guangdong, China. J. Econ. Entomol. 2018, 111, 239–246. [Google Scholar] [CrossRef]
- Han, J.H.; You, J.H.; Koo, C.D.; Yoon, C.M.; Choi, K.S.; Shin, S.C.; Kim, G.H. Emergence timing of the pine sawyer beetle, Monochamus saltuarius (Coleoptera: Cerambycidae) by tree species. Korean J. Appl. Entomol. 2009, 48, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.Q.; Wang, X.Y.; Gould, J.R.; Reardon, R.C.; Zhang, Y.N.; Liu, G.J.; Liu, E.S. Biology and behavior of Spathius Agrili, a parasitoid of the emerald ash borer, Agrilus planipennis, in China. J. Insect Sci. 2010, 10, 30. [Google Scholar] [CrossRef]
- Gonçalves, E.; Figueiredo, A.C.; Barroso, J.G.; Henriques, J.; Sousa, E.; Bonifácio, L. Effect of Monochamus galloprovincialis feeding on Pinus pinaster and Pinus pinea, oleoresin and insect volatiles. Phytochemistry 2020, 169, 112–159. [Google Scholar] [CrossRef]
- Pershing, J.C.; Linit, M.J. Biology of Monochamus carolinensis (Coleoptera: Cerambycidae) on scotch pine in Missouri. J. Kans. Entomol. Soc. 1986, 59, 706–711. [Google Scholar]
- Gould, J.R.; Ayer, T.; Fraser, I. Effects of rearing conditions on reproduction of Spathius Agrili (Hymenoptera: Braconidae), a parasitoid of the emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2011, 104, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Le Masurier, A.D. A comparative study of the relationship between host size and brood size in Apanteles spp. (Hymenoptera: Braconidae). Ecol. Entomol. 1987, 12, 383–393. [Google Scholar] [CrossRef]
- Seo, B.S.; Jeon, H.S.; Kim, J.B.; Cho, G.S. Development of pine wilt disease (Bursaphelenchus Xylophilus) prevention system. In Proceedings of the Korean Society of Surveying Geodesy Photogrammetry Cartography Conference, Jeju, Korea; Korean Society of Surveying, Geodesy, Photogrammetry and Cartography: Seoul, Korea, 2007; pp. 283–287. [Google Scholar]
- Rebek, E.J.; Sadof, C.S. Effects of pesticide applications on the euonymus scale (Homoptera: Diaspididae) and its parasitoid, Encarsia citrina (Hymenoptera: Aphelinidae). J. Econ. Entomol. 2003, 96, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Hassell, M.P.; Waage, J.K. Host-parasitoid population interactions. Annu. Rev. Entomol. 1984, 29, 89–114. [Google Scholar] [CrossRef]
- Frank, S.D. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biol. Control 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides? A brief review. Environ. Int. 2016, 89–90, 7–11. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Larval Stages of Parasitized Hosts (n) | Ave. No of Parasitoids on a Host Larva | Ave. Pupation Period (Day) | Sex Ratio (♂: Total) |
|---|---|---|---|---|
| M. alternatus | 2nd (7) | 3.15 ± 1.64 | 6.6 ± 1.39 | 0.16 |
| 3rd (1) | ||||
| 4th (3) | ||||
| 5th (1) | ||||
| M. saltuarius | 1st (33) | 1.48 ± 0.62 | 7.76 ± 3.06 | 0.37 |
| 2nd (3) |
| Host | Variable | n | r | df | t | p |
|---|---|---|---|---|---|---|
| M. alternatus | Size of parasitized host larvae × no. of S. verustus | 12 | 0.35 | 10 | 1.16 | 0.27 |
| Size of parasitized host larvae × size of female S. verustus | 25 | 0.26 | 23 | 1.3 | 0.21 | |
| Host density × sex ratio | 10 | −0.55 | 8 | −1.87 | 0.1 | |
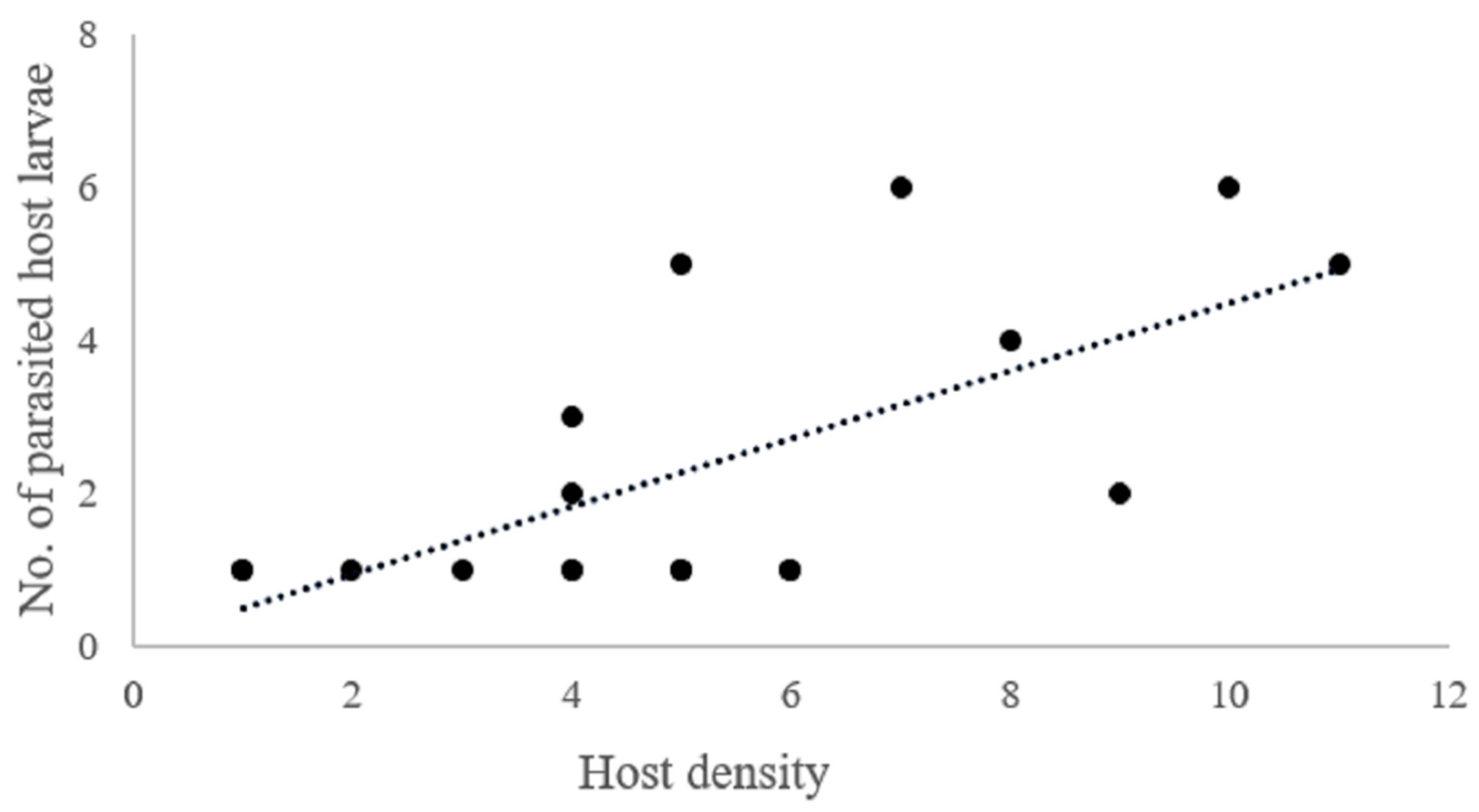
| Host density × no. of parasitized host larvae | 18 | 0.66 | 16 | 3.56 | 0.002 ** | |
| M. saltuarius | Size of parasitized host larvae × no. of S. verustus | 36 | −0.27 | 34 | −1.64 | 0.1 |
| Size of parasitized host larvae × size of female S. verustus | 57 | 0.09 | 55 | 0.6 | 0.55 | |
| Host density × sex ratio | 29 | −0.27 | 27 | −1.44 | 0.16 | |
| Host density × no. of parasitized host larvae | 45 | 0.17 | 43 | 1.11 | 0.27 |
| Year | Variable | Parameter | Mean (%) ± SD | n | df | p |
|---|---|---|---|---|---|---|
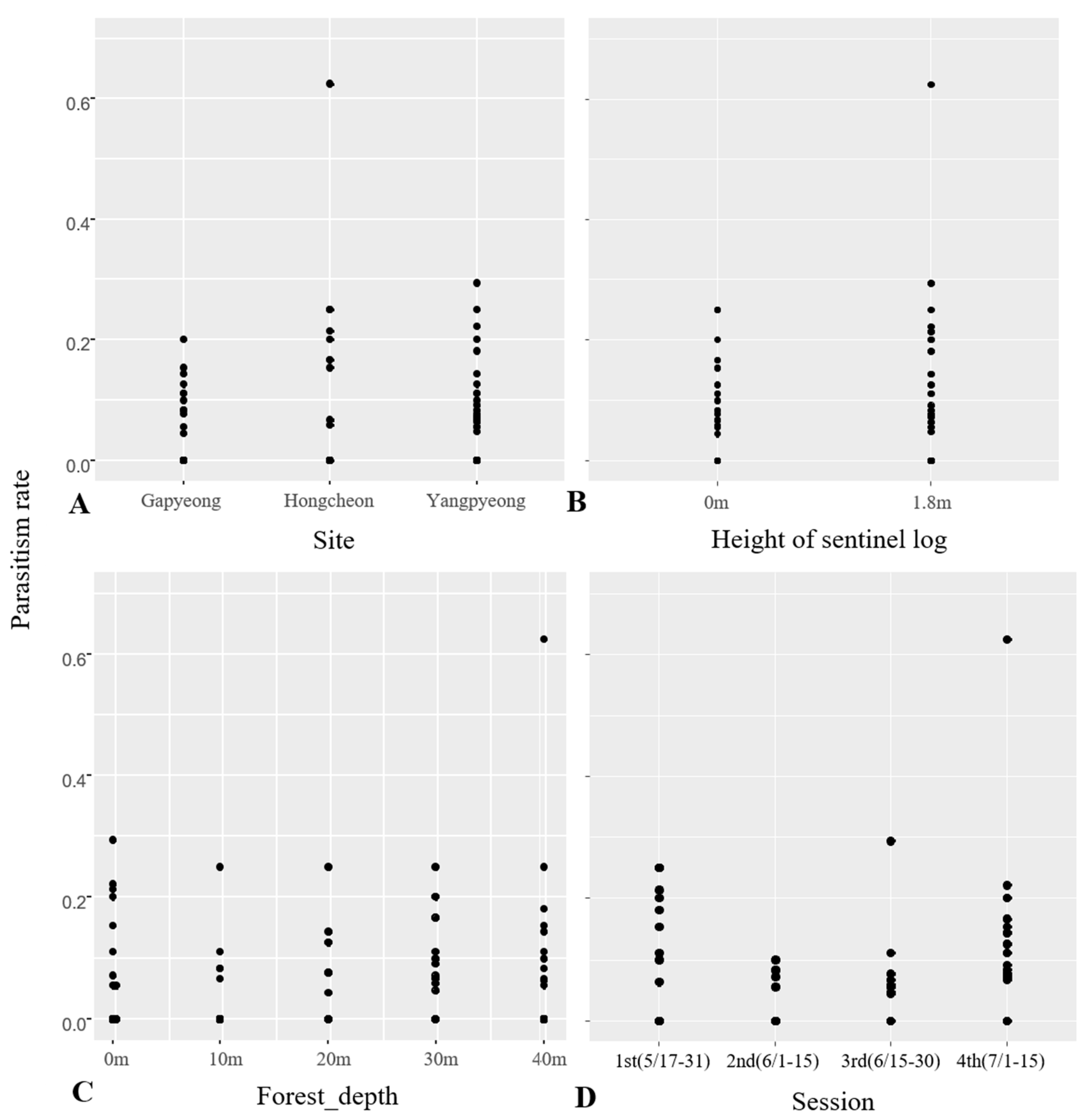
| 2016 | Site | Geoje | 1.66% ± 6.61 | 96 † | 2 | 4.279 × 10−12 *** |
| Suncheon | 5.59% ± 18.2 | |||||
| Jinju | 17.62% ± 31.26 | |||||
| Height of sentinel log | 0 m | 7.75% ± 21.18 | 1 | 0.94 | ||
| 1.8 m | 8.83% ± 23.33 | |||||
| 2017 | Site | Gapyeong | 0.68% ± 0.03 | 480 † | 2 | 0.001501 ** |
| Yangpyeong | 1.25% ± 0.04 | |||||
| Hongcheon | 3.01% ± 0.09 | |||||
| Height of sentinel log | 0 m | 1.2% ± 0.04 | 1 | 0.38 | ||
| 1.8 m | 1.4% ± 0.05 | |||||
| Forest depth | 0 m | 1.43 ± 0.05 | 4 | 0.52 | ||
| 10 m | 0.7% ± 0.03 | |||||
| 20 m | 0.89% ± 0.03 | |||||
| 30 m | 1.6% ± 0.04 | |||||
| 40 m | 1.91% ± 0.04 | |||||
| Session | 1st (5/17–31) | 3.35% ± 0.07 | 3 | 2.849 × 10−7 *** | ||
| 2nd (6/1–15) | 0.28% ± 0.01 | |||||
| 3rd (6/15–30) | 0.64% ± 0.02 | |||||
| 4th (7/1–15) | 2.12% ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-S.; Jung, J.-K.; Hong, K.-J.; Kim, C.-J.; Lee, B.-W.; Kim, I.-K. Discovery and Biology of Spathius verustus Chao (Hymenoptera: Braconidae), a Potential Biological Agent on Two Monochamus Vectors of the Pinewood Nematode. Forests 2022, 13, 955. https://doi.org/10.3390/f13060955
Kim M-S, Jung J-K, Hong K-J, Kim C-J, Lee B-W, Kim I-K. Discovery and Biology of Spathius verustus Chao (Hymenoptera: Braconidae), a Potential Biological Agent on Two Monochamus Vectors of the Pinewood Nematode. Forests. 2022; 13(6):955. https://doi.org/10.3390/f13060955
Chicago/Turabian StyleKim, Moo-Sung, Jong-Kook Jung, Ki-Jeong Hong, Chang-Jun Kim, Bong-Woo Lee, and Il-Kwon Kim. 2022. "Discovery and Biology of Spathius verustus Chao (Hymenoptera: Braconidae), a Potential Biological Agent on Two Monochamus Vectors of the Pinewood Nematode" Forests 13, no. 6: 955. https://doi.org/10.3390/f13060955
APA StyleKim, M. -S., Jung, J. -K., Hong, K. -J., Kim, C. -J., Lee, B. -W., & Kim, I. -K. (2022). Discovery and Biology of Spathius verustus Chao (Hymenoptera: Braconidae), a Potential Biological Agent on Two Monochamus Vectors of the Pinewood Nematode. Forests, 13(6), 955. https://doi.org/10.3390/f13060955