
Wood Anatomical Traits Respond to Climate but More Individualistically as Compared to Radial Growth: Analyze Trees, Not Means

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Tree Species
2.2. Climate Data
2.3. Tree-Ring Width and Wood Anatomical Data
2.4. Statistical Analyses
3. Results
3.1. Statistics and Variability of Radial Growth and Wood Anatomy
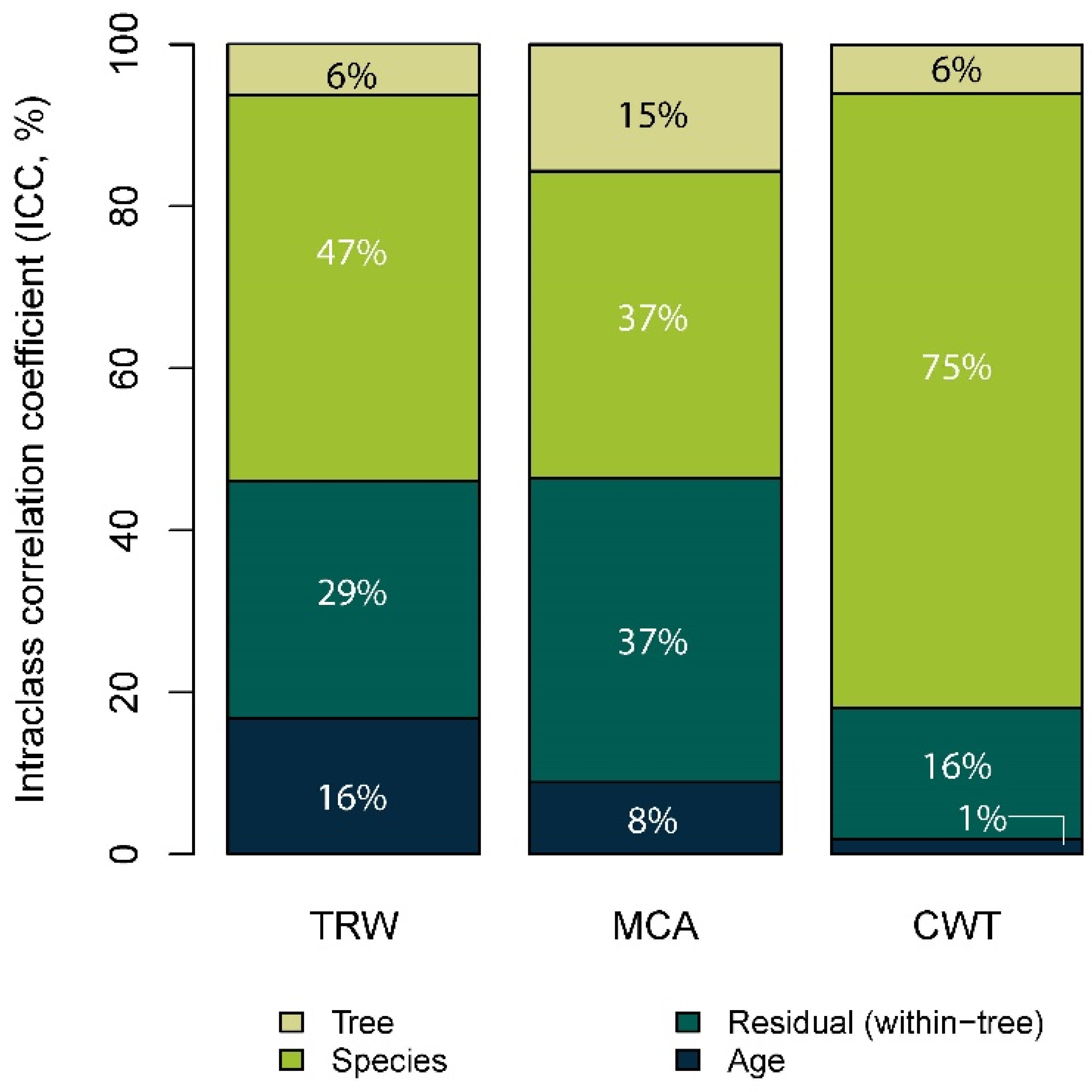
3.2. Trait Variance Decomposition between Species and Individuals
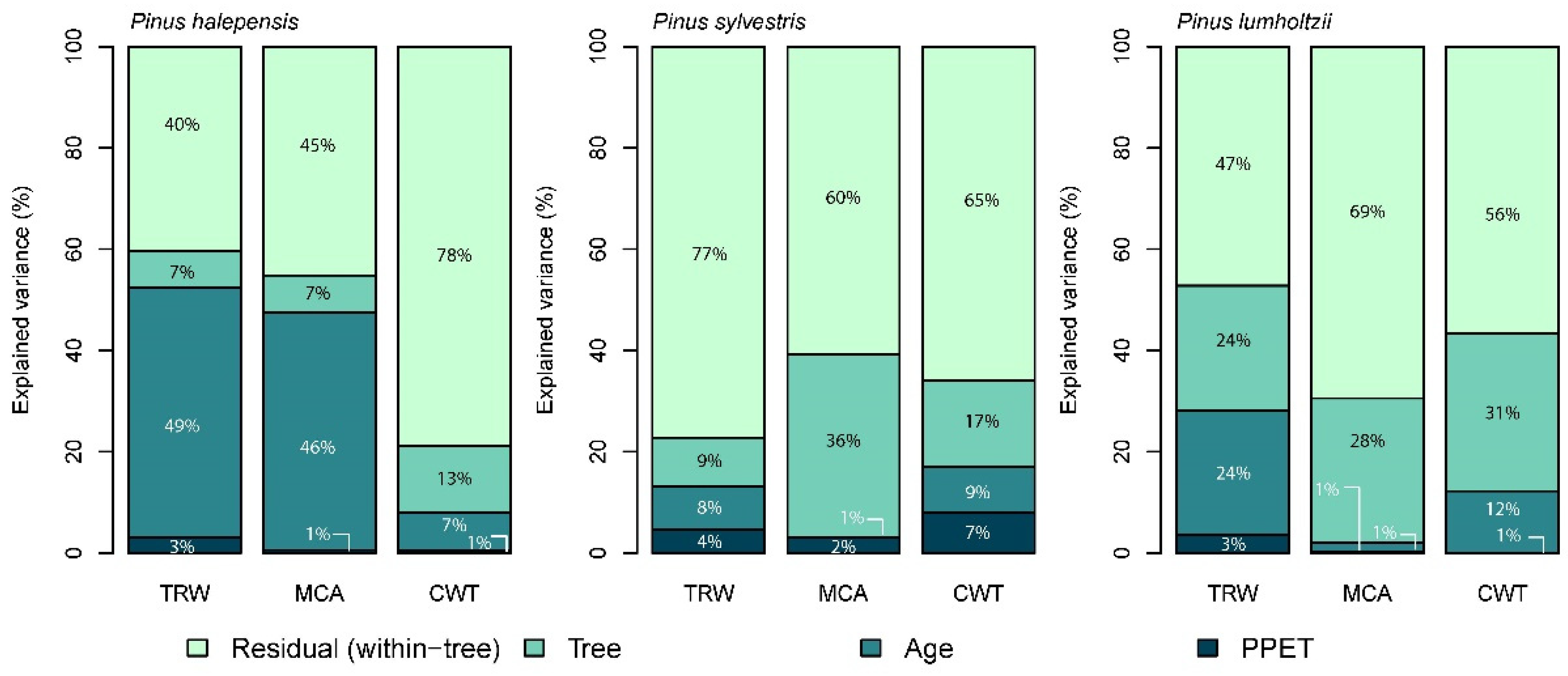
3.3. Effects of Seasonal Climate on Measured Traits and Variance Decomposition
4. Discussion
4.1. The Percentage of Variance Explained for Each Trait Differs between Tree Species
4.2. The Percentage of Variance Explained Differs between Traits and Individuals
4.3. There Is Great Variability between Individuals of the Same Population for Wood Anatomical Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Fritts, H.C. Tree Rings and Climate; Elsevier: Amsterdam, The Netherlands, 1976; ISBN 978-0-12268-450-0. [Google Scholar]
- Amoroso, M.M.; Daniels, L.D.; Baker, P.J.; Camarero, J.J. Dendroecology: Tree-Ring Analyses Applied to Ecological Studies; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-31961-668-1. [Google Scholar]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Voltas, J.; Saurer, M.; Berninger, F.; Esper, J.; Andreu-Hayles, L.; Daux, V.; Helle, G.; Leuenberger, M.; Loader, N.J.; et al. Spatio-temporal patterns of tree growth as related to carbon isotope fractionation in European forests under changing climate. Glob. Ecol. Biogeogr. 2019, 28, 1295–1309. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Gazol, A.; Piovesan, G.; Borghetti, M.; Baliva, M.; Gentilesca, T.; Rita, A.; Schettino, A.; Ripullone, F. Mediterranean old-growth forests exhibit resistance to climate warming. Sci. Total Environ. 2021, 801, 149684. [Google Scholar] [CrossRef]
- Camarero, J.J.; González de Andrés, E.; Sangüesa-Barreda, G.; Rita, A.; Colangelo, M. Long- and short-term impacts of a defoliating moth plus mistletoe on tree growth, wood anatomy and water-use efficiency. Dendrochronologia 2019, 56, 125598. [Google Scholar] [CrossRef]
- Rodríguez-González, P.M.; Colangelo, M.; Sánchez-Miranda, Á.; Sánchez-Salguero, R.; Campelo, F.; Rita, A.; Gomes Marques, I.; Albuquerque, A.; Ripullone, F.; Camarero, J.J. Climate, drought and hydrology drive narrow-leaved ash growth dynamics in southern European riparian forests. For. Ecol. Manag. 2021, 490, 119128. [Google Scholar] [CrossRef]
- Castellaneta, M.; Rita, A.; Camarero, J.J.; Colangelo, M.; Ripullone, F. Declines in canopy greenness and tree growth are caused by combined climate extremes during drought-induced dieback. Sci. Total Environ. 2022, 813, 152666. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Hughes, M.K.; Shashkin, A.V. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 3-54-026086-2. [Google Scholar]
- Pellizzari, E.; Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Carrer, M. Wood anatomy and carbon-isotope discrimination support long-term hydraulic deterioration as a major cause of drought-induced dieback. Glob. Chang. Biol. 2016, 22, 2125–2137. [Google Scholar] [CrossRef]
- Puchi, P.F.; Camarero, J.J.; Battipaglia, G.; Carrer, M. Retrospective analysis of wood anatomical traits and tree-ring isotopes suggests site-specific mechanisms triggering Araucaria araucana drought-induced dieback. Glob. Chang. Biol. 2021, 27, 6394–6408. [Google Scholar] [CrossRef]
- Fonti, P.; Bryukhanova, M.V.; Myglan, V.S.; Kirdyanov, A.V.; Naumova, O.V.; Vaganov, E.A. Temperature-induced responses of xylem structure of Larix sibirica (Pinaceae) from the Russian Altay. Am. J. Bot. 2013, 100, 1332–1343. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; Del Castillo, E.M.; Campelo, F.; Vavrčík, H.; et al. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 15160. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.; Camarero, J.J.; Carrer, M. Linking wood anatomy and xylogenesis allows pinpointing of climate and drought influences on growth of coexisting conifers in continental Mediterranean climate. Tree Physiol. 2016, 36, 502–512. [Google Scholar] [CrossRef] [Green Version]
- González-Cásares, M.; Camarero, J.J.; Colangelo, M.; Rita, A.; Pompa-García, M. High responsiveness of wood anatomy to water availability and drought near the equatorial rear edge of Douglas-fir. Can. J. For. Res. 2019, 49, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Colangelo, M.; Rodríguez-González, P.M.; Sánchez-Miranda, Á.; Sánchez-Salguero, R.; Campelo, F.; Rita, A.; Ripullone, F. Wood anatomy and tree growth covary in riparian ash forests along climatic and ecological gradients. Dendrochronologia 2021, 70, 125891. [Google Scholar] [CrossRef]
- Gao, J.; Yang, B.; Peng, X.; Rossi, S. Tracheid development under a drought event producing intra-annual density fluctuations in the semi-arid China. Agric. For. Meteorol. 2021, 308–309, 108572. [Google Scholar] [CrossRef]
- Von Arx, G.; Carrer, M. ROXAS—A new tool to build centuries-long tracheid-lumen chronologies in conifers. Dendrochronologia 2014, 32, 290–293. [Google Scholar] [CrossRef]
- Fonti, P.; García-González, I. Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol. 2004, 163, 77–86. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Fajardo, A.; Caetano-Sánchez, C.; Camarero, J.J.; Olano, J.M. Xylem anatomy needs to change, so that conductivity can stay the same: Xylem adjustments across elevation and latitude in Nothofagus pumilio. Ann. Bot. 2020, 125, 1101–1112. [Google Scholar] [CrossRef]
- Alla, A.Q.; Camarero, J.J. Contrasting responses of radial growth and wood anatomy to climate in a Mediterranean ring-porous oak: Implications for its future persistence or why the variance matters more than the mean. Eur. J. For. Res. 2012, 131, 1537–1550. [Google Scholar] [CrossRef]
- Hacke, U.G.; Spicer, R.; Schreiber, S.G.; Plavcová, L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ. 2017, 40, 831–845. [Google Scholar] [CrossRef] [PubMed]
- Yasue, K.; Funada, R.; Kobayashi, O.; Ohtani, J. The effects of tracheid dimensions on variations in maximum density of Picea glehnii and relationships to climatic factors. Trees 2000, 14, 223–229. [Google Scholar] [CrossRef]
- González-Cásares, M.; Pompa-García, M.; Camarero, J.J. Differences in climate–growth relationship indicate diverse drought tolerances among five pine species coexisting in Northwestern Mexico. Trees 2017, 31, 531–544. [Google Scholar] [CrossRef]
- Pacheco, A.; Camarero, J.J.; Pompa-García, M.; Battipaglia, G.; Voltas, J.; Carrer, M. Growth, wood anatomy and stable isotopes show species-specific couplings in three Mexican conifers inhabiting drought-prone areas. Sci. Total Environ. 2020, 698, 134055. [Google Scholar] [CrossRef]
- Pompa-García, M.; Hevia, A.; Camarero, J.J. Minimum and maximum wood density as proxies of water availability in two Mexican pine species coexisting in a seasonally dry area. Trees 2021, 35, 597–607. [Google Scholar] [CrossRef]
- Pacheco, A.; Camarero, J.J.; Ribas, M.; Gazol, A.; Gutierrez, E.; Carrer, M. Disentangling the climate-driven bimodal growth pattern in coastal and continental Mediterranean pine stands. Sci. Total Environ. 2018, 615, 1518–1526. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- González Elizondo, M.; González Elizondo, M.D.S.; Villanueva Díaz, J.; Cerano Paredes, J. Assessment of the dendroclimatic potential of Pinus lumholtzii BL Rob. & Fernald. Rev. Mex. Cienc. For. 2017, 8, 28–54. [Google Scholar]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958-2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef] [Green Version]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement; Tree-Ring Society: Loveland, CO, USA, 1983; ISSN 0041-2198. [Google Scholar]
- Gärtner, H.; Nievergelt, D. The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 2010, 28, 85–92. [Google Scholar] [CrossRef]
- Dyachuk, P.; Arzac, A.; Peresunko, P.; Videnin, S.; Ilyin, V.; Assaulianov, R.; Babushkina, E.A.; Zhirnova, D.; Belokopytova, L.; Vaganov, E.A.; et al. AutoCellRow (ACR)—A new tool for the automatic quantification of cell radial files in conifer images. Dendrochronologia 2020, 60, 125687. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology; Springer: Dordrecht, The Netherlands, 1990; ISBN 978-9-04814-060-2. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Clim. Appl. Meteor. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program. 2021. Available online: https://CRAN.R-project.org/package=dplR (accessed on 27 August 2021).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS.; Springer: New York, NY, USA, 2000; ISBN 0-38-798957-9. [Google Scholar]
- Firn, J.; Nguyen, H.; Schütz, M.; Risch, A.C. Leaf trait variability between and within subalpine grassland species differs depending on site conditions and herbivory. Proc. R. Soc. B 2019, 286, 20190429. [Google Scholar] [CrossRef] [PubMed]
- Dobbert, S.; Pape, R.; Löffler, J. How does spatial heterogeneity affect inter-and intraspecific growth patterns in tundra shrubs? J. Ecol. 2021, 109, 4115–4131. [Google Scholar] [CrossRef]
- Nakagawa, S.; Johnson, P.C.D.; Schielzeth, H. The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interface 2017, 14. [Google Scholar] [CrossRef] [Green Version]
- Barton, K. MuMIn: Multi-Model Inference. 2022. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 27 August 2021).
- Shaw, M.; Rights, J.D.; Sterba, S.K.; Flake, J.K. r2mlm: R-Squared Measures for Multilevel Models; 2020. Available online: https://github.com/mkshaw/r2mlm (accessed on 27 August 2021).
- Rights, J.D.; Sterba, S.K. Quantifying explained variance in multilevel models: An integrative framework for defining R-squared measures. Psychol. Methods 2019, 24, 309–338. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Soft. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Vergarechea, M.; Alfaro-Sánchez, R.; Cattaneo, N.; Vicente-Serrano, S.M. Tree growth is more limited by drought in rear-edge forests most of the times. For. Ecosyst. 2021, 8, 25. [Google Scholar] [CrossRef]
- Bose, A.K.; Gessler, A.; Bolte, A.; Bottero, A.; Buras, A.; Cailleret, M.; Camarero, J.J.; Haeni, M.; Hereş, A.-M.; Hevia, A.; et al. Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob. Chang. Biol. 2020, 26, 4521–4537. [Google Scholar] [CrossRef]
- Olson, M.E.; Soriano, D.; Rosell, J.A.; Anfodillo, T.; Donoghue, M.J.; Edwards, E.J.; León-Gómez, C.; Dawson, T.; Camarero, J.J.; Castorena, M.; et al. Plant height and hydraulic vulnerability to drought and cold. Proc. Natl. Acad. Sci. USA 2018, 115, 7551–7556. [Google Scholar] [CrossRef] [Green Version]
- Tardif, J.C.; Conciatori, F. Influence of climate on tree rings and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec, Canada. Can. J. For. Res. 2006, 36, 2317–2330. [Google Scholar] [CrossRef] [Green Version]
- Tardif, J.C.; Kames, S.; Nolin, A.F.; Bergeron, Y. Earlywood vessels in black ash (Fraxinus nigra Marsh.) trees show contrasting sensitivity to hydroclimate variables according to flood exposure. Front. Plant Sci. 2021, 12, 754596. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.E.; Anfodillo, T.; Gleason, S.M.; McCulloh, K.A. Tip-to-base xylem conduit widening as an adaptation: Causes, consequences, and empirical priorities. New Phytol. 2021, 229, 1877–1893. [Google Scholar] [CrossRef]
- Herrera, C.M. The ecology of subindividual variability in plants: Patterns, processes, and prospects. Web Ecol. 2017, 17, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Carrer, M.; von Arx, G.; Castagneri, D.; Petit, G. Distilling allometric and environmental information from time series of conduit size: The standardization issue and its relationship to tree hydraulic architecture. Tree Physiol. 2015, 35, 27–33. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pinus halepensis | Pinus sylvestris | Pinus lumholtzii | |||||||
|---|---|---|---|---|---|---|---|---|---|
| TRW | MCA | CWT | TRW | MCA | CWT | TRW | MCA | CWT | |
| No. trees | 5 | 5 | 5 | 5 | 5 | 5 | 4 | 4 | 4 |
| Span | 1970–2012 | 1970–2012 | 1970–2012 | 1950–2010 | 1950–2010 | 1950–2010 | 1970–2013 | 1970–2013 | 1970–2013 |
| Mean | 1.82 | 274.08 | 4.12 | 0.61 | 435.21 | 4.16 | 1.06 | 442.05 | 6.05 |
| SD | 1.04 | 79.29 | 0.33 | 0.36 | 106.09 | 0.38 | 0.45 | 93.99 | 0.81 |
| AR1 | 0.48 | 0.46 | 0.47 | 0.53 | 0.20 | 0.27 | 0.61 | 0.18 | 0.53 |
| MS | 0.38 | 0.20 | 0.04 | 0.44 | 0.24 | 0.08 | 0.26 | 0.20 | 0.09 |
| Rbar | 0.45 | 0.61 | 0.19 | 0.69 | 0.41 | 0.23 | 0.39 | 0.04 | 0.11 |
| EPS | 0.77 | 0.86 | 0.48 | 0.91 | 0.77 | 0.59 | 0.58 | 0.13 | 0.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rita, A.; Camarero, J.J.; Colangelo, M.; de Andrés, E.G.; Pompa-García, M. Wood Anatomical Traits Respond to Climate but More Individualistically as Compared to Radial Growth: Analyze Trees, Not Means. Forests 2022, 13, 956. https://doi.org/10.3390/f13060956
Rita A, Camarero JJ, Colangelo M, de Andrés EG, Pompa-García M. Wood Anatomical Traits Respond to Climate but More Individualistically as Compared to Radial Growth: Analyze Trees, Not Means. Forests. 2022; 13(6):956. https://doi.org/10.3390/f13060956
Chicago/Turabian StyleRita, Angelo, Jesús Julio Camarero, Michele Colangelo, Ester González de Andrés, and Marín Pompa-García. 2022. "Wood Anatomical Traits Respond to Climate but More Individualistically as Compared to Radial Growth: Analyze Trees, Not Means" Forests 13, no. 6: 956. https://doi.org/10.3390/f13060956
APA StyleRita, A., Camarero, J. J., Colangelo, M., de Andrés, E. G., & Pompa-García, M. (2022). Wood Anatomical Traits Respond to Climate but More Individualistically as Compared to Radial Growth: Analyze Trees, Not Means. Forests, 13(6), 956. https://doi.org/10.3390/f13060956