
(Re)Designing Urban Parks to Maximize Urban Heat Island Mitigation by Natural Means



Abstract
:1. Introduction
2. Materials and Methods
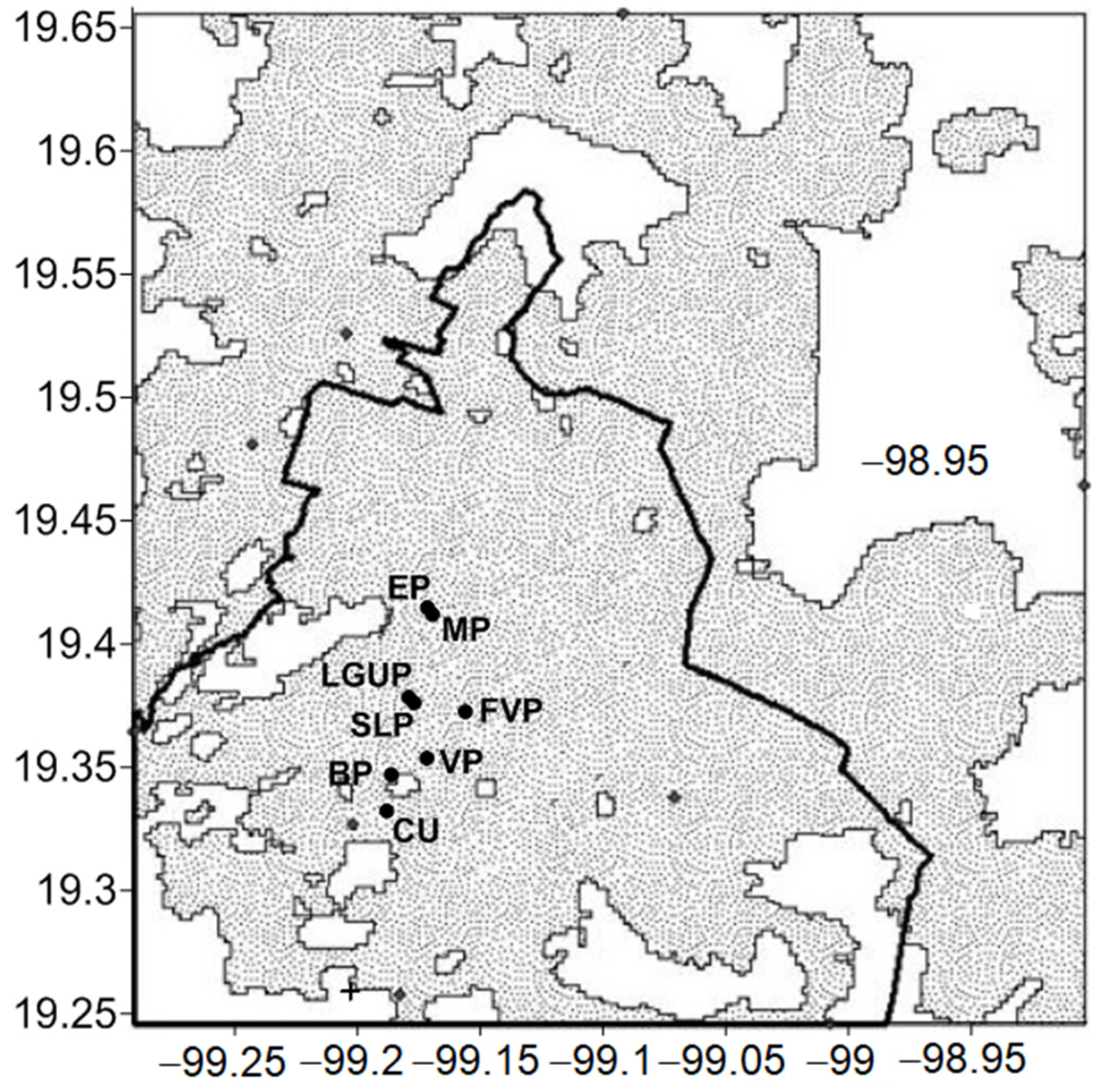
2.1. Study Area
2.2. Species Selection
2.3. Data Collection
2.4. The Thermal UrbaN Environment Energy Balance Model
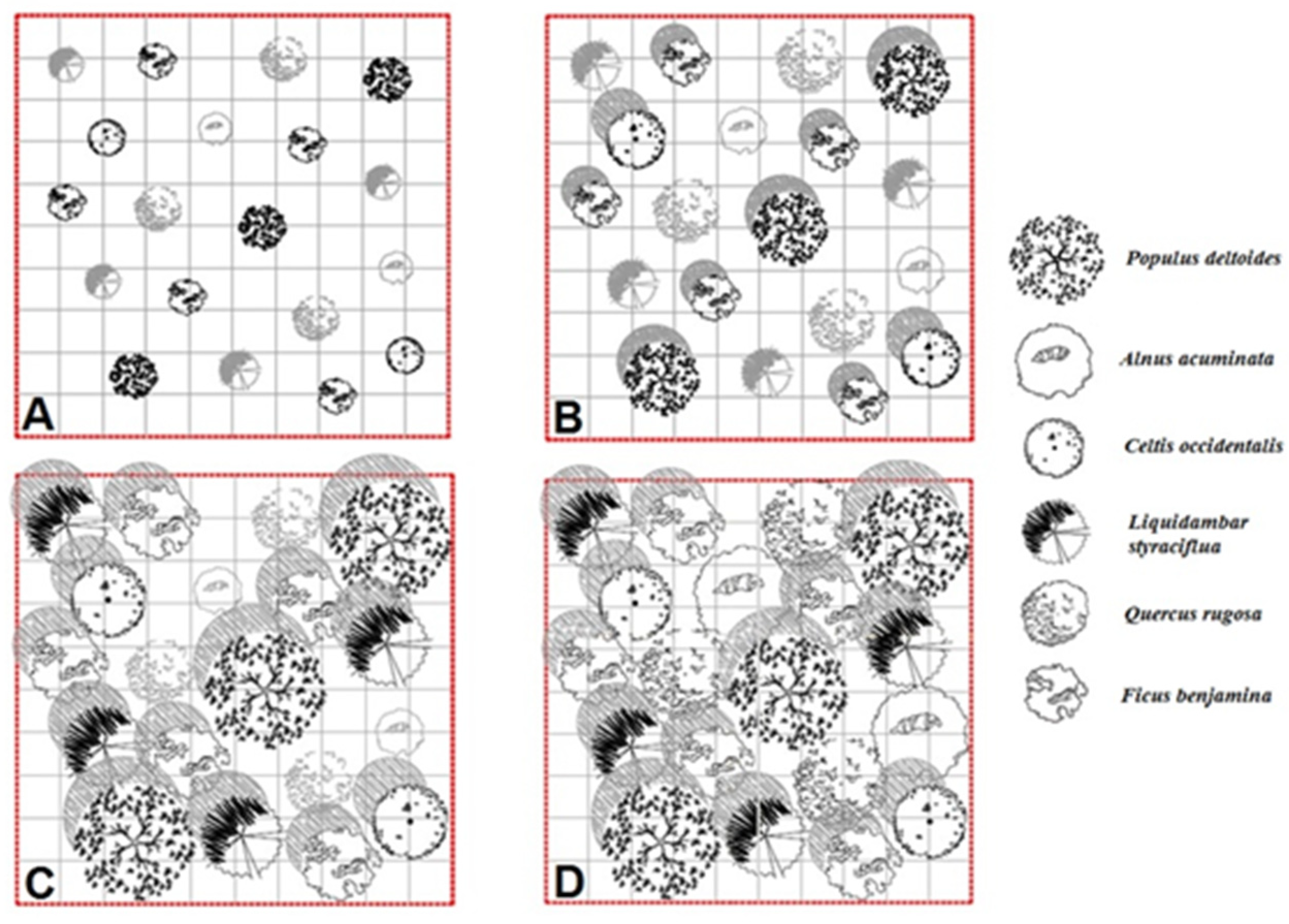
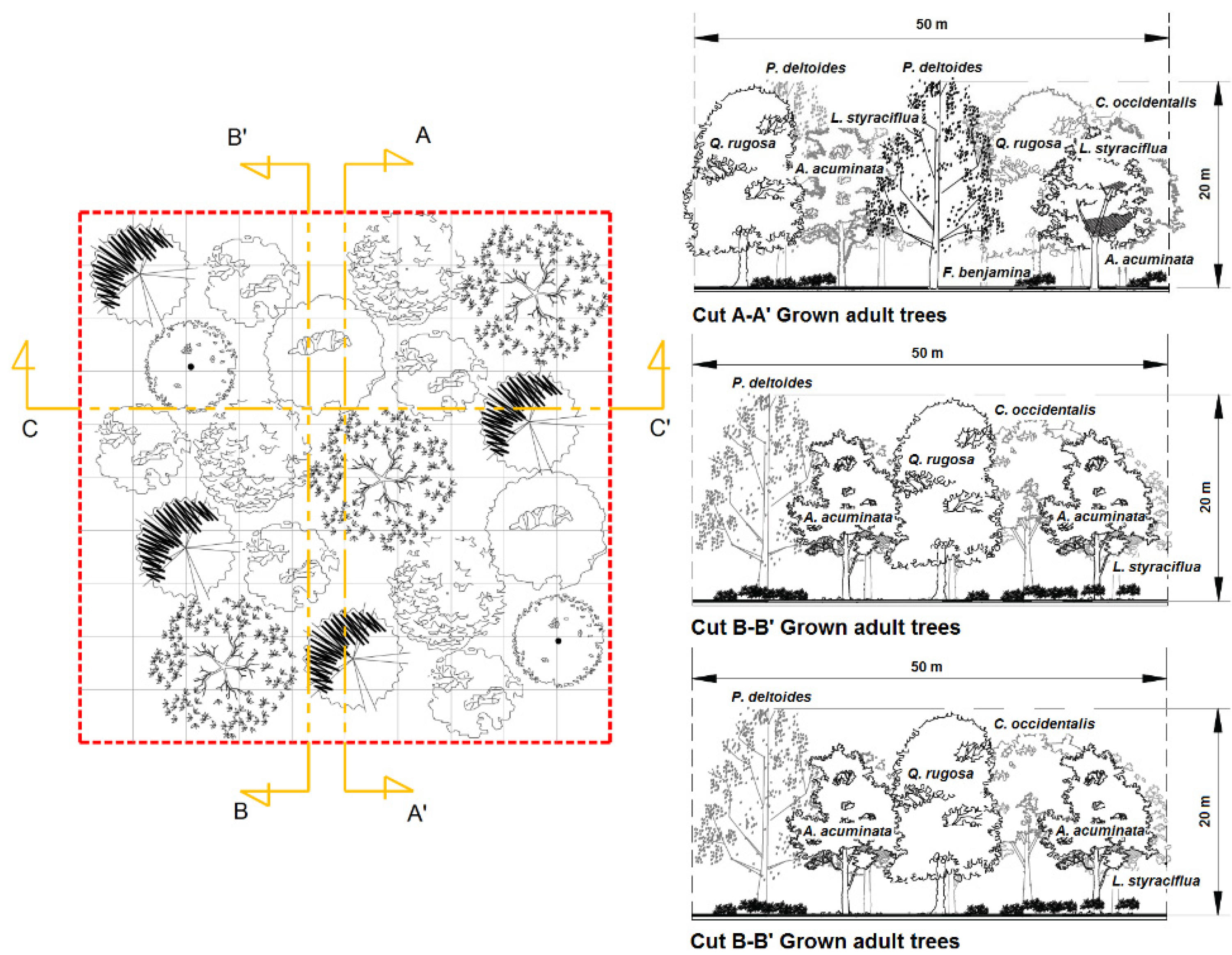
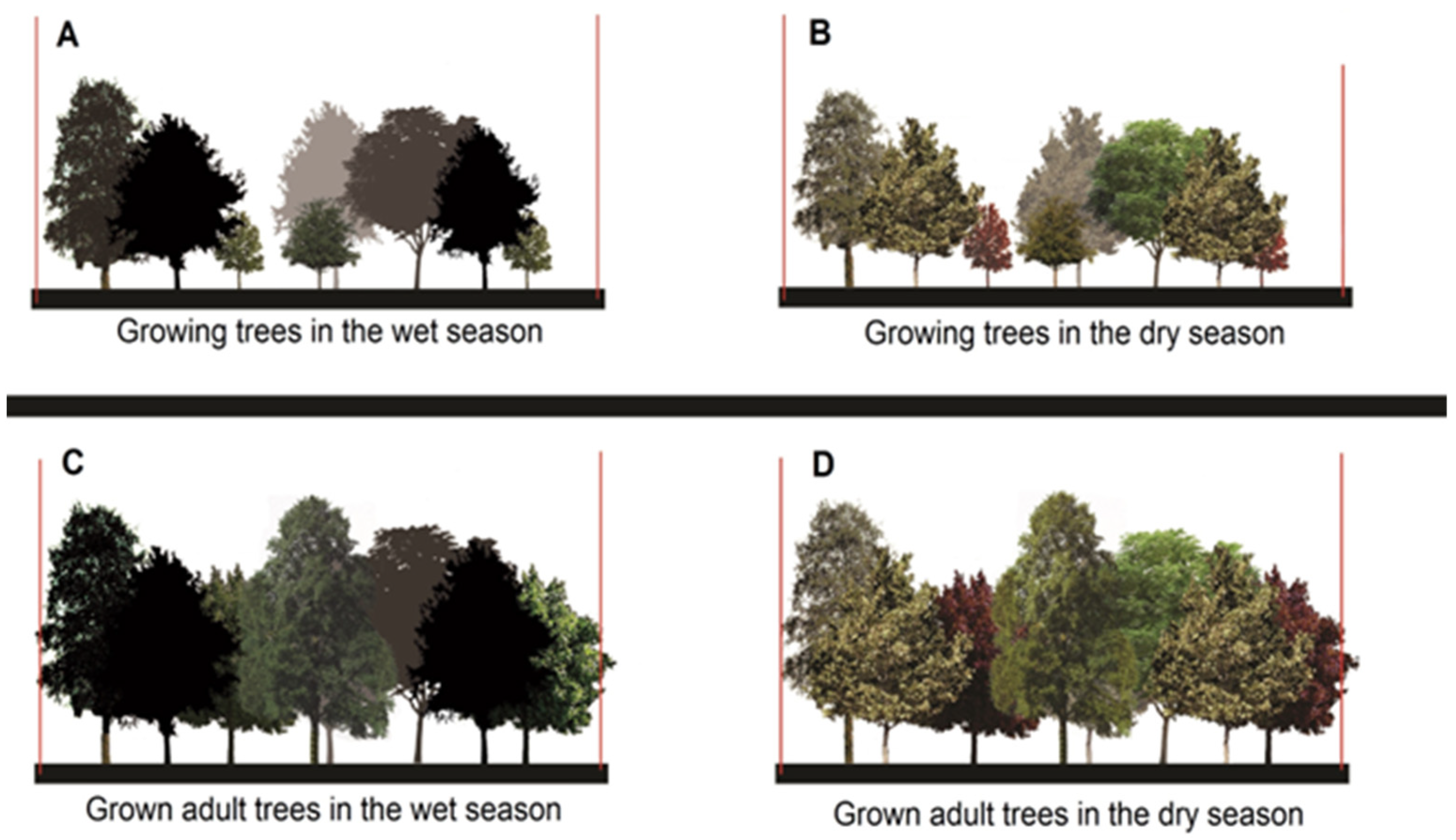
2.5. (Re)Designing Urban Parks
2.6. Statistical Analysis
3. Results
4. Discussion
4.1. Transpiration and Stomatal Conductance
4.2. (Re)Designing Urban Parks
4.3. Additional Management Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oke, T.R. The heat island of the urban boundary layer: Characteristics, causes and effects. Wind Clim. Cities 1995, 277, 81–107. [Google Scholar]
- Williams, N.S.G.; Hahs, A.K.; Vesk, P.A. Urbanisation, plant traits and the composition of urban floras. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 78–86. [Google Scholar] [CrossRef]
- Miller, R.W.; Hauer, R.J.; Werner, L.P. Urban Forestry: Planning and Managing Urban Greenspaces, 3rd ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2015. [Google Scholar]
- Livesley, S.; McPherson, E.G.; Calfapietra, C. The urban forest and ecosystem services: Impacts on Urban water, heat, and pollution cycles at the tree, street, and city scale. J. Environ. Qual. 2016, 45, 119–124. [Google Scholar] [CrossRef]
- Beer, A.R. Developing tools to monitor the effectiveness of development plans. In Proceedings of the PRO/ECE-Workshop on Sustainable Urban Development: Research and Experiments, 1st ed.; Van der Vegt, H., Ter Heide, H., Tjallingii, S., Van Alphen, D., Eds.; Delft University Press: Dordrecht, The Netherlands, 1994; pp. 69–85. [Google Scholar]
- Sayad, B.; Alkama, D.; Ahmad, H.; Baili, J.; Aljahdaly, N.H.; Menni, Y. Nature-based solutions to improve the summer thermal comfort outdoors. Case Stud. Therm. Eng. 2021, 28, 101399. [Google Scholar] [CrossRef]
- Ulrich, R.S. Natural versus urban sciences: Some psycho-physiological effects. Environ. Behav. 1981, 13, 523–556. [Google Scholar] [CrossRef]
- Chiesura, A. The role of urban parks for the sustainable city. Landsc. Urban Plan. 2004, 68, 129–138. [Google Scholar] [CrossRef]
- Donovan, G.H.; Prestemon, J. The Effect of Trees on Crime in Portland, Oregon. Environ. Behav. 2010, 44, 3–30. [Google Scholar] [CrossRef]
- Vandermeulen, V.; Verspecht, A.; Vermeire, B.; Van Huylenbroeck, G.; Gellynck, X. The use of economic valuation to create public support for green infrastructure investments in urban areas. Landsc. Urban Plan. 2011, 103, 198–206. [Google Scholar] [CrossRef]
- Thompson, C.W. Urban open space in the 21st century. Landsc. Urban Plan. 2002, 60, 59–72. [Google Scholar] [CrossRef]
- Farhadi, H.; Faisi, M.; Sanaieian, H. Mitigating the urban heat island in residential area of Tehran: Investigating the role of vegetation, material, and orientation of buildings. Sustain. Cities Soc. 2019, 46, 101448. [Google Scholar] [CrossRef]
- Rakoto, P.Y.; Deilami, K.; Hurley, J.; Amati, M.; Sun, Q.C. Revisiting the cooling effects of urban greening: Planning implications of vegetation types and spatial configuration. Urban For. Urban Green. 2021, 64, 127266. [Google Scholar] [CrossRef]
- Cortes, A.; Rejuso, A.J.; Santos, J.A.; Blanco, A. Evaluating mitigation strategies for urban heat island in Mandaue City using ENVI-met. J. Urban Manag. 2022, 11, 97–106. [Google Scholar] [CrossRef]
- Buyadi, S.N.A.; Mohd, W.M.N.W.; Misni, A. Green spaces growth impact on the urban microclimate. Procedia—Soc. Behav. Sci. 2013, 105, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Gioia, A.; Paolini, L.; Malizia, A.; Oltra-Carrió, R.; Sobrino, J.A. Size matters: Vegetation patch size and surface temperature relationship in foothills cities of northwestern Argentina. Urban Ecosyst. 2014, 17, 1161–1174. [Google Scholar] [CrossRef]
- Alavipanah, S.; Wegmann, M.; Qureshi, S.; Weng, Q.; Koellner, T. The Role of Vegetation in Mitigating Urban Land Surface Temperatures: A Case Study of Munich, Germany during the Warm Season. Sustainability 2015, 7, 4689–4706. [Google Scholar] [CrossRef] [Green Version]
- Gates, D.M. Transpiration and leaf temperature. Annu. Rev. Plant Physiol. 1968, 19, 211–238. [Google Scholar] [CrossRef]
- Ballinas, M.; Barradas, V.L. The urban tree as a Tool to mitigate the urban heat island in Mexico City: A simple phenomenological model. J. Environ. Qual. 2016, 45, 157–166. [Google Scholar] [CrossRef]
- Freeman, S. Biological Science, 3rd ed.; Benjamin Cummings: San Francisco, CA, USA, 2007. [Google Scholar]
- Esau, K. 1977. In Anatomy of Seed Plants, 2nd ed.; Wiley and Sons: Denver, CO, USA, 2007. [Google Scholar]
- García-Sánchez, I.E.; Barradas, V.L.; de León Hill, C.A.P.; Esperón-Rodríguez, M.; Pérez, I.R.; Ballinas, M. Effect of heavy metals and environmental variables on the assimilation of CO2 and stomatal conductance of Ligustrum lucidum, an urban tree from Mexico City. Urban For. Urban Green. 2019, 42, 72–81. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2007; pp. 325–366. ISBN 978-0-12-088765-1. [Google Scholar]
- IPCC. Climate Change 2021: The Physical Science Basis, the Working Group I contribution to the Sixth Assessment Report; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2021. [Google Scholar]
- Seppanen, O.; Fisk, W.J.; Faulkner, D. Control of Temperature for Health and Productivity in Offices; Lawrence Berkeley National Laboratory: Berkeley, CA, USA, 2004. [Google Scholar]
- Tse, W.L.; So, A.T. The Importance of Human Productivity to Air-Conditioning Control in Office Environments. HVACR Res. 2007, 13, 3–21. [Google Scholar] [CrossRef]
- Lowe, S.A. An energy and mortality impact assessment of the urban heat island in the US. Environ. Impact Assess. Rev. 2016, 56, 139–144. [Google Scholar] [CrossRef]
- Ballinas, M. Mitigación de la isla de calor urbana: Estudio de caso de la zona metropolitana de la Ciudad de México. Master’s Theise, Universidad Nacional Autónoma de México, Ciudad de México, Mexico, 2011. [Google Scholar]
- Ramos-Niembro, G.; Heard, C.; Sánchez-Viveros, A. Simulación de escenarios de ahorro y uso eficiente de energía, con medidas de control pasivo. Energía Racion. FIDE 1998, 28, 17–29. [Google Scholar]
- INEGI. Available online: http://cuentame.inegi.org.mx/monografias/informacion/df/territorio/clima.aspx?tema=me&e=09 (accessed on 23 April 2021).
- SMN. Available online: https://smn.conagua.gob.mx/es/climatologia/informacion-climatologica/normales-climatologicas-por-estado (accessed on 23 April 2021).
- Barradas, V.L.; Tejeda-Martínez, A.; Jáuregui, E. Energy balance measurements in a suburban vegetated area in Mexico City. Atmos. Environ. 1999, 33, 4109–4113. [Google Scholar] [CrossRef]
- Oke, T.R.; Sproken-Smith, R.A.; Jáuregui, E.; Grimmond, C.S.B. The energy balance of central Mexico City during the dry season. Atmos. Environ. 1999, 33, 3919–3930. [Google Scholar] [CrossRef]
- Jauregui, E. Mesomicroclima de la Ciudad de México; Instituto de Geografía UNAM: Ciudad de México, Mexico, 1971. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Kumar, R.; Kaushik, S.C. Performance evaluation of green roof and shading for thermal protection of buildings. Build. Environ. 2005, 40, 1505–1511. [Google Scholar] [CrossRef]
- Ballinas, M.; Barradas, V.L. Transpiration and stomatal conductance as potential mechanisms to mitigate the heat load in Mexico City. Urban For. Urban Green. 2016, 20, 152–159. [Google Scholar] [CrossRef]
- Thom, A.S. Momentum, Mass and Heat Exchange of Plant Communities, 1st ed.; Vegetation and the atmosphere Academic Press: London, UK, 1976; pp. 57–109. [Google Scholar]
- Monteith, J.L. Evaporation and environment. Symposium of the Soc. Exp. Biol. 1965, 19, 205–224. [Google Scholar]
- Panagopoulos, T. Linking forestry, sustainability and aesthetics. Ecol. Econ. 2009, 68, 2485–2489. [Google Scholar] [CrossRef]
- Yılmaz, S.; Özgüner, H.; Mumcu, S. An aesthetic approach to planting design in urban parks and greenspaces. Landsc. Res. 2018, 43, 965–983. [Google Scholar] [CrossRef]
- Avolio, M.L.; Pataki, D.E.; Trammell, T.L.; Endter-Wada, J. Biodiverse cities: The nursery industry, homeowners, and neighborhood differences drive urban tree composition. Ecol. Monogr. 2018, 88, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Hovenden, J.M.; Brodribb, T. Altitude of origin influences stomatal conductance and therefore maximum assimilation rate in southern Beech, Nothofagus cunninghamii. Aust. J. Plant Physiol. 2000, 27, 451–456. [Google Scholar] [CrossRef]
- Esperón-Rodríguez, M.; Barradas, V.L. Potential vulnerability to climate change of four tree species from the central mountain region of Veracruz, Mexico. Clim. Res. 2014, 60, 163–174. [Google Scholar] [CrossRef]
- Collatz, G.J.; Ball, J.T.; Grivet, C.; Berry, A.J. Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: A model that includes a laminar boundary layer. Agric. For. Meteorol. 1991, 54, 107–136. [Google Scholar] [CrossRef]
- Bunce, J.A. Does transpiration control stomatal responses to water vapour pressure deficit? Plant Cell Environ. 1997, 20, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Scherer, T.F.; Seelig, B.; Franzen, D. Soil, Water and Plant Characteristics Important to Irrigation; NDSU Extension Service: Fargo, ND, USA; Available online: https://www.ag.ndsu.edu/publications/crops/soil-water-and-plant-characteristics-important-to-irrigation (accessed on 13 April 2022).
- McCulloh, K.A.; Woodruff, D.R. Linking stomatal sensitivity and whole-tree hydraulic architecture. Tree Physiol. 2012, 32, 369–372. [Google Scholar] [CrossRef] [Green Version]
- McCree, K.J. Photosynthetically active radiation. Physiol. Plant Ecol. 1981, 12/A, 41–55. [Google Scholar]
- Jones, H.G. Plant and Microclimate, 3rd ed.; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Chazdon, R.L.; Pearcy, R.W.; Lee, D.W.; Fetcher, N. Photosynthetic responses of tropical forest plants to contrasting light environments. In Tropical Forest Plant Ecophysiology; Mulkey, S.S., Chazdon, R.L., Smith, A.P., Eds.; Springer: Boston, MA, USA, 1996; pp. 5–55. [Google Scholar]
- Zhao, D.; Lei, Q.; Shi, Y.; Wang, M.; Chen, S.; Shah, K.; Ji, W. Role of species and planting configuration on transpiration and microclimate for urban trees. Forests 2020, 11, 825. [Google Scholar] [CrossRef]
- Arriaga, A.; Cruz, G.; Ortiz, G. Relaciones Hídricas en Las Plantas; P&V Editores: Mexico City, Mexico, 1999. [Google Scholar]
- Esperon-Rodriguez, M.; Rymer, P.D.; Power, S.A.; Challis, A.; Marchin, R.M.; Tjoelker, M.G. Functional adaptations and trait plasticity of urban trees along a climatic gradient. Urban For. Urban Green. 2020, 54, 126771. [Google Scholar] [CrossRef]
- McPherson, E.G.; Berry, A.M.; van Doorn, N.S. Performance testing to identify climate-ready trees. Urban For. Urban Green. 2018, 29, 28–39. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pugnaire, F.I. The role of nurse plants in the restoration of degraded environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- González, M.F. Las comunidades Vegetales de México, 2nd ed.; SEMARNAT & INE: Ciudad de México, Mexico, 2004; pp. 56–75. [Google Scholar]
- Braun-Blanquet, J. Fitosociología. Bases Para El Estudio de Las Comunidades Vegetales; H. Blume Ediciones: Madrid, Spain, 1979. [Google Scholar]
- Parker, J. Effects of variations in the root-leaf ratio on transpiration rate. Plant Physiol. 1949, 24, 739–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geller, G.N.; Smith, W.K. Influence of leaf size, orientation, and arrangement on temperature and transpiration in three high-elevation, large-leafed herbs. Oecologia 1982, 53, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.B.; Edwards, G.E.; Tanner, C.B. Effects of light, carbon dioxide, and temperature on photosynthesis, oxygen inhibition of photosynthesis, and transpiration in Solanum tuberosum. Plant Physiol. 1977, 59, 868–872. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.W. Arboriculture, Integrated Management of Landscape Trees, Shrubs and Vines, 2nd ed.; Pentice-Hall Inc.: Hoboken, NJ, USA, 1992; pp. 318–345. [Google Scholar]
- Barradas, V.L. Air temperature and humidity and human comfort index of some city parks of Mexico City. Int. J. Biometeorol. 1991, 35, 24–28. [Google Scholar] [CrossRef]
- Nielsen, A.B.; Van Den Bosch, M.; Maruthaveeran, S.; van den Bosch, C.K. Species richness in urban parks and its drivers: A review of empirical evidence. Urban Ecosyst. 2014, 17, 305–327. [Google Scholar] [CrossRef]
- Stigsdotter, U.K.; Corazon, S.S.; Sidenius, U.; Refshauge, A.D.; Grahn, P. Forest design for mental health promotion—Using perceived sensory dimensions to elicit restorative responses. Landsc. Urban Plan. 2017, 160, 1–15. [Google Scholar] [CrossRef]
- Alvey, A.A. Promoting and preserving biodiversity in the urban forest. Urban For. Urban Green. 2006, 5, 195–201. [Google Scholar] [CrossRef]
- Morgenroth, J.; Östberg, J.; Harðarson, S.B.; Wiström, B.; Nielsen, A.B. Urban Tree Diversity for Sustainable Cities—Policy Brief; Nordic Forest Research: Joensuu, Finland, 2016. [Google Scholar]
- Watkins, H.; Hirons, A.; Sjöman, H.; Cameron, R.; Hitchmough, J.D. Can Trait-Based Schemes Be Used to Select Species in Urban Forestry? Front. Sustain. Cities 2021, 3, 54618. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Flower | Flowering | Fruit | A | DSH | H | CD | LS (l, w) | LH | Location (Park) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acacia longifolia | Fabaceae | Small yellow or golden-yellow | Mainly during winter and early spring | A very elongated pod | 3.5 | 0.48 | 8.0 | 9.3 | 8.5, 1.4 | Evergreen | Ciudad Universitaria |
| Acer negundo | Sapindaceae | Several in a cluster, greenish-yellow | Spring | Samaras 1-seeded, pale yellow | 3.2 | 0.31 | 8.8 | 7.5 | 13.5, 8.6 | Deciduous | Luis G. Urbina |
| Alnus acuminata | Betulaceae | Inflorescences are catkins | Spring and Summer | Long dehiscent, woody brown fruits | 2.5 | 0.29 | 8.5 | 6.5 | 7.6, 3.7 | Evergreen / deciduous | Ciudad Universitaria/ Viveros |
| Buddleja cordata | Loganiaceae | Small fragrant flowers, white, cream, or yellow | Summer and early autumn | Capsule fruit | 2.0 | 0.16 | 4.0 | 3.4 | 12.8, 5.3 | Evergreen | Ciudad Universitaria |
| Celtis occidentalis | Cannabaceae | Gold/Yellow, green, orange, purple/lavender | Spring | Round fleshy berry-like drupes | 4.6 | 0.33 | 8.5 | 9.5 | 11.1, 4.9 | Deciduous | Luis G. Urbina |
| Ficus benjamina | Moraceae | Inflorescences black, green, orange, purple/lavender, red/burgundy | Spring and Summer | Globose to slightly oblong fig | 5.4 | 0.76 | 11.5 | 15.0 | 7.6, 3.4 | Evergreen | Bombilla/ Francisco Villa |
| Fraxinus uhdei | Oleaceae | Small and insignificant, without petals, green, white or yellow | Winter | Brown samaras | 4.4 | 0.25 | 15.0 | 11.0 | 8.1, 3.5 | Deciduous | San Lorenzo/ Luis G. Urbina |
| Lagerstroemia indica | Lythraceae | Numerous and irregular, gold/yellow, pink, purple/lavender, red/burgundy, white | Spring, Summer, and Fall | Brown/copper capsule | 2.1 | 0.15 | 4.3 | 5.2 | 5.5, 3.1 | Evergreen | Ciudad Universitaria |
| Ligustrum lucidum | Oleaceae | Small, perfect, creamy white flowers | Summer | Black, blue druppe | 4.0 | 0.15 | 13.0 | 8.1 | 6.3, 3.1 | Evergreen | Ciudad Universitaria |
| Liquidambar styraciflua | Hamamelidaceae | Yellow-green flowers | Spring and Summer | Brown/copper capsule | 4.6 | 0.27 | 13.6 | 11.0 | 7.5, 6.7 | Deciduous | Ciudad Universitaria/ Viveros |
| Populus alba | Salicaceae | Inflorescence catkin | Spring and Summer | White loculicidal capsules | 2.6 | 0.17 | 6.0 | 4.3 | 3.8, 3.7 | Deciduous | Luis G. Urbina/ Bombilla |
| Populus deltoides | Salicaceae | Inflorescence catkin, green red/burgundy | Spring | Brown/copper, Green or white capsule | 3.8 | 0.47 | 13.3 | 9.3 | 11.1, 8.7 | Evergreen | España/Mexico |
| Quercus rugosa | Fagaceae | Pistillate catkins pubescent | Spring | Acorn | 3.1 | 0.17 | 5.5 | 7.8 | 13.6, 8.6 | Evergreen /deciduous | Viveros/ Francisco Villa |
| Robinia pseudoacacia | Fabaceae | Fragrant wisteria-like white flowers | Spring and Summer | Brown/copper, purple/lavender legume | 3.0 | 0.19 | 7.0 | 8.0 | 7.5, 4.4 | Deciduous | Viveros |
| Ulmus parvifolia | Ulmaceae | Insignificant and reddish green | Spring, Summer, and Fall | Brown/copper samara | 1.5 | 0.23 | 5.0 | 3.0 | 8.0, 3.7 | Deciduous | Viveros |
| Nr | Species | gS | E | CP | QN | VPD | TL | TA |
|---|---|---|---|---|---|---|---|---|
| 1 | Liquidambar styraciflua | 3.81 | 0.0357 | 87.2 | 365.5 | 2.79 | 28.2 | 28.2 |
| 2 | Alnus acuminata | 3.52 | 0.0352 | 86.0 | 119.7 | 2.93 | 29.6 | 29.6 |
| 3 | Quercus rugosa | 3.65 | 0.0348 | 84.9 | 345.0 | 3.38 | 32.5 | 32.4 |
| 4 | Ligustrum lucidum | 2.89 | 0.0324 | 79.2 | 475.0 | 2.73 | 27.8 | 27.7 |
| 5 | Ficus benjamina | 3.01 | 0.0307 | 74.9 | 112.2 | 2.69 | 27.8 | 27.5 |
| 6 | Populus deltoides | 2.17 | 0.0215 | 52.5 | 162.2 | 3.01 | 29.6 | 29.2 |
| 7 | Fraxinus udhei | 2.6 | 0.0194 | 47.5 | 355.3 | 2.90 | 29.5 | 28.9 |
| 8 | Populus alba | 2.57 | 0.0187 | 45.5 | 410.48 | 2.80 | 28.5 | 28.3 |
| 9 | Celtis occidentalis | 2.85 | 0.0166 | 40.5 | 295.13 | 2.81 | 29.0 | 28.2 |
| 10 | Acer negundo | 2.10 | 0.0130 | 31.8 | 37.83 | 2.51 | 26.9 | 25.9 |
| 11 | Acacia longifolia | 1.82 | 0.0117 | 28.6 | 299.83 | 2.56 | 26.8 | 26.7 |
| 12 | Ulmus parvifolia | 1.6 | 0.0108 | 26.4 | 384.15 | 3.01 | 30.1 | 30.1 |
| 13 | Robinia pseudoacacia | 1.62 | 0.0107 | 26.2 | 462.17 | 3.21 | 31.5 | 31.4 |
| 14 | Lagerstroemia indica | 1.26 | 0.0102 | 25.1 | 346.5 | 2.01 | 24.1 | 23.8 |
| 15 | Buddleja cordata | 1.08 | 0.0089 | 21.7 | 482.98 | 2.14 | 24.6 | 24.1 |
| Group | Species | TA | VPD | E |
|---|---|---|---|---|
| 1 | Lagerstroemia indica Buddleia cordata | Low | Low | Low. Air is nearly saturated with water vapor; therefore, the leaf cannot release water through stomata |
| 2 | Alnus acuminata Ulmus parvifolia Quercus rugosa Robinia pseudoacacia | High | Moderate | High. VPD is favorable and leaf temperature is high |
| 3 | Populus deltoides Liquidambar styraciflua Ficus benjamina Fraxinus uhdei Celtis occidentalis Ligustrum lucidum Populus alba Acacia longifolia | High | Moderate | Moderate. VPD is favorable and leaf temperature is high |
| 4 | Acer negundo | Low | High | Low. Stomatal closure could avoid water loss and desiccation of leaves |
| Species | QEP | TAP | ΔT |
|---|---|---|---|
| Populus deltoides | 96.8 | 24.5 | 3.4 |
| Alnus acuminata | 99.6 | 24.4 | 3.6 |
| Celtis occidentalis | 86.4 | 24.9 | 3.1 |
| Liquidambar styraciflua | 185.9 | 21.1 | 6.9 |
| Quercus rugosa | 122.1 | 23.5 | 4.4 |
| Ficus benjamina | 187.7 | 21.0 | 7.0 |
| Weighted average | 142.7 | 22.7 | 5.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barradas, V.L.; Miranda, J.A.; Esperón-Rodríguez, M.; Ballinas, M. (Re)Designing Urban Parks to Maximize Urban Heat Island Mitigation by Natural Means. Forests 2022, 13, 1143. https://doi.org/10.3390/f13071143
Barradas VL, Miranda JA, Esperón-Rodríguez M, Ballinas M. (Re)Designing Urban Parks to Maximize Urban Heat Island Mitigation by Natural Means. Forests. 2022; 13(7):1143. https://doi.org/10.3390/f13071143
Chicago/Turabian StyleBarradas, Victor L., Jennifer A. Miranda, Manuel Esperón-Rodríguez, and Monica Ballinas. 2022. "(Re)Designing Urban Parks to Maximize Urban Heat Island Mitigation by Natural Means" Forests 13, no. 7: 1143. https://doi.org/10.3390/f13071143
APA StyleBarradas, V. L., Miranda, J. A., Esperón-Rodríguez, M., & Ballinas, M. (2022). (Re)Designing Urban Parks to Maximize Urban Heat Island Mitigation by Natural Means. Forests, 13(7), 1143. https://doi.org/10.3390/f13071143