
Ecological Strategy Spectra for Communities of Different Successional Stages in the Tropical Lowland Rainforest of Hainan Island



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Strategy
2.2. Determination of Functional Traits
2.3. Data Analysis
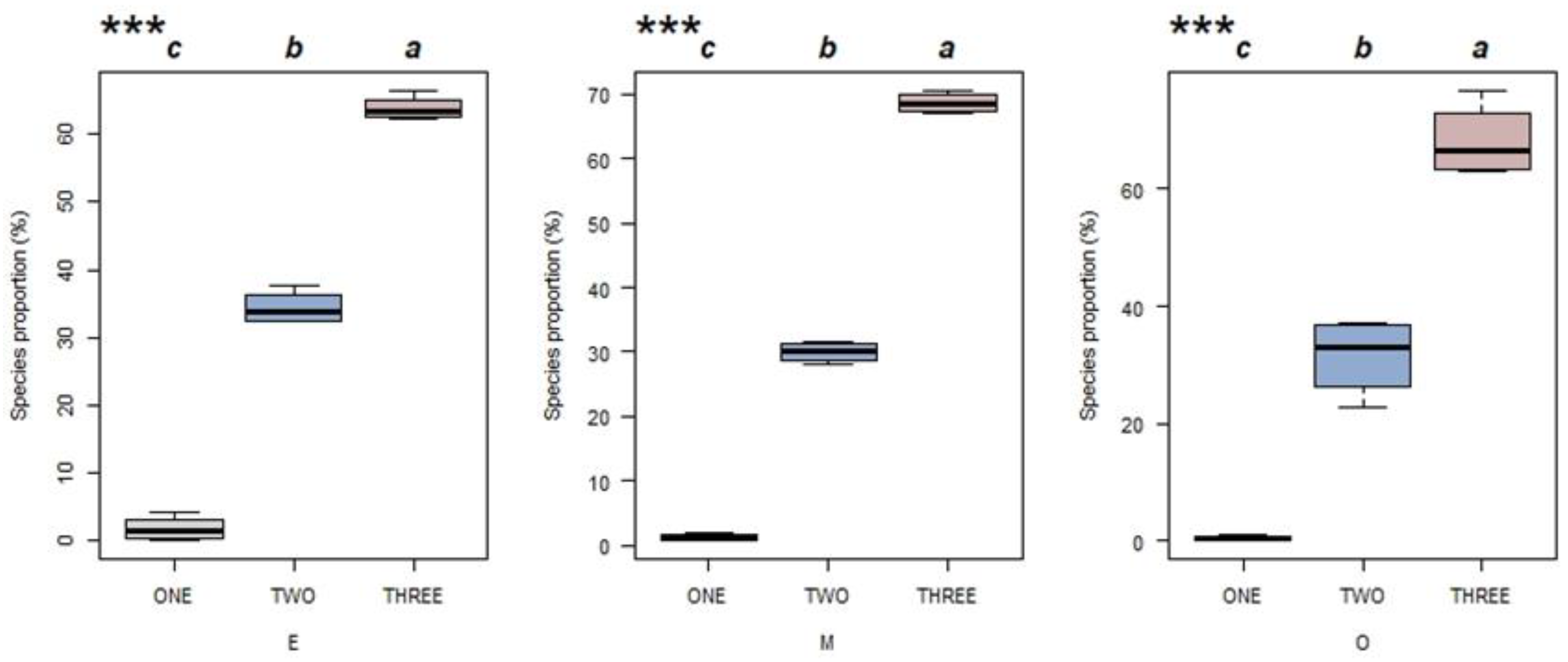
3. Results
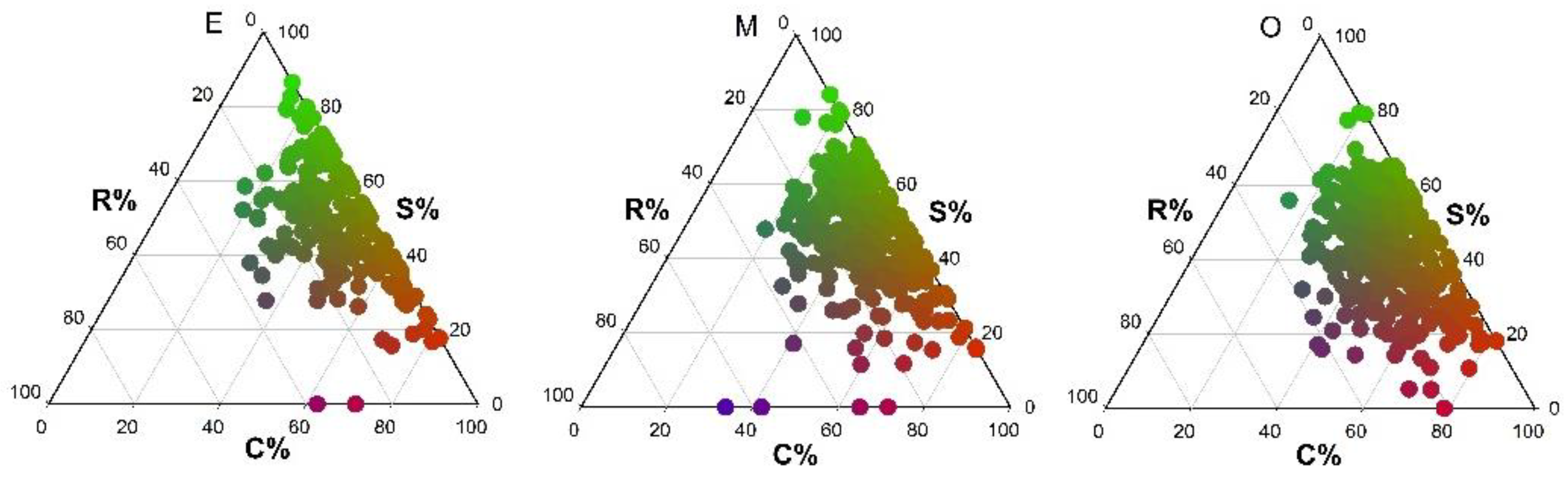
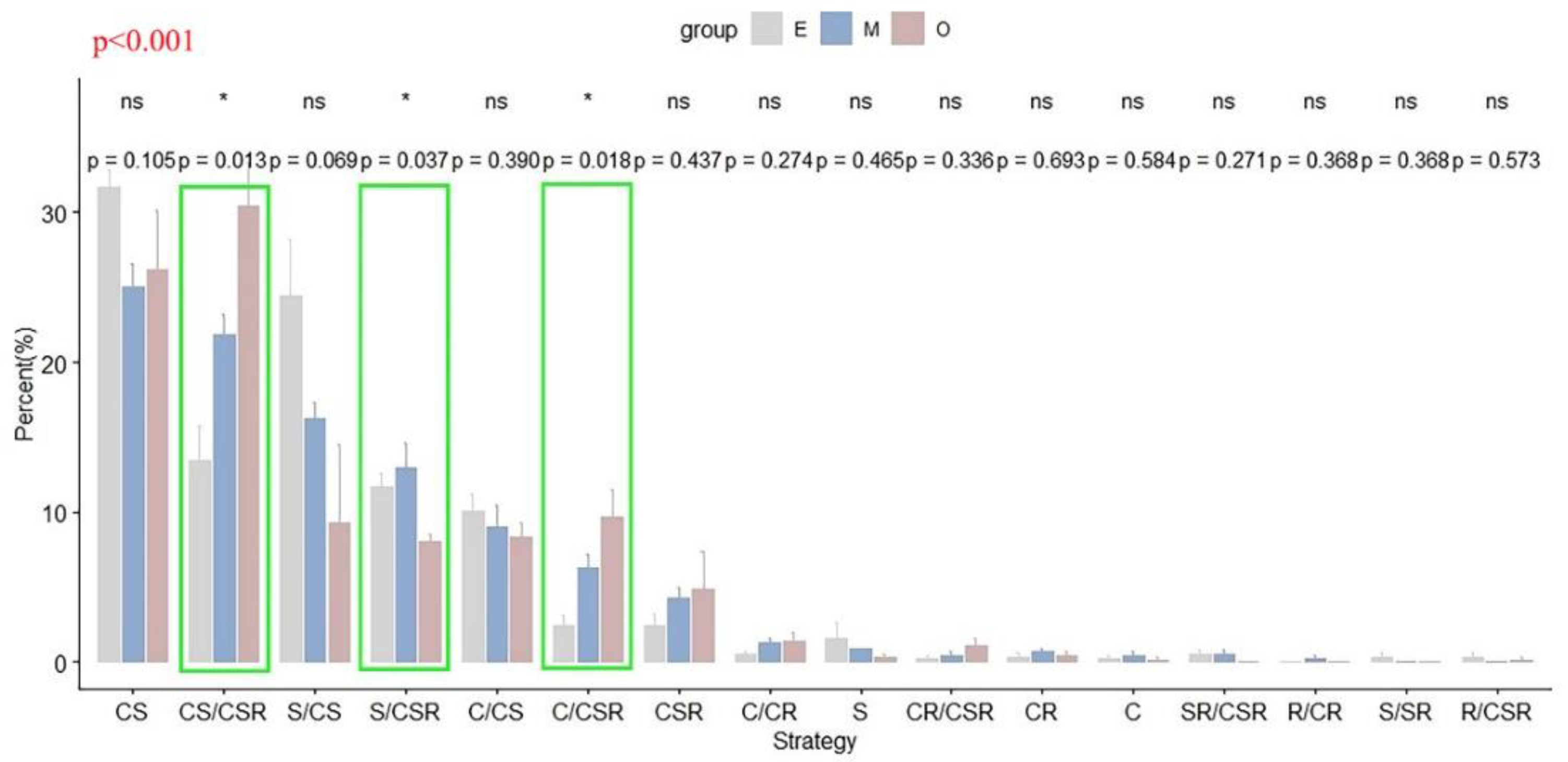
3.1. Types of Community Ecological Strategies in Successional Stages
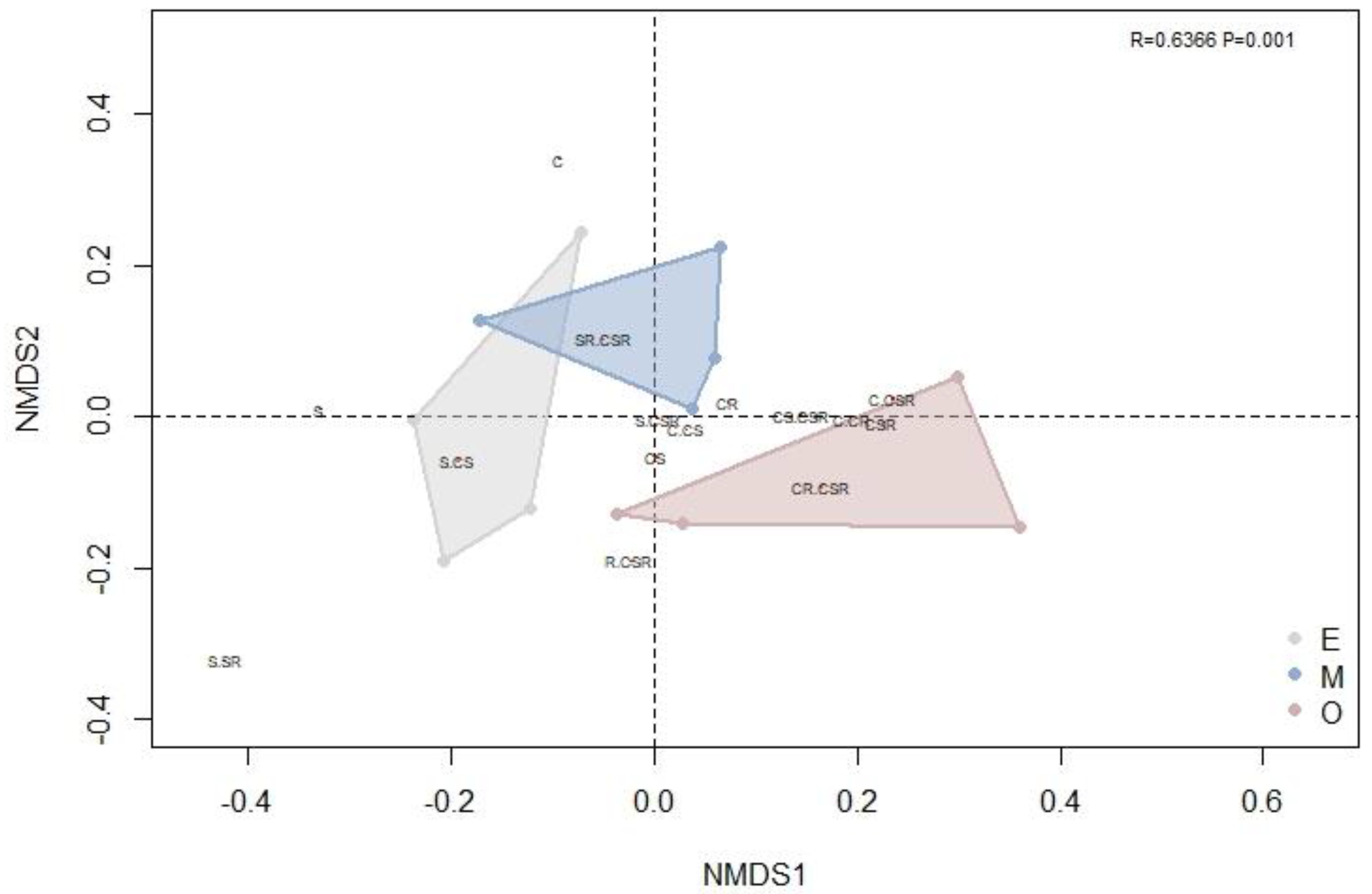
3.2. Successional Dynamics in Community Ecological Strategy Spectra
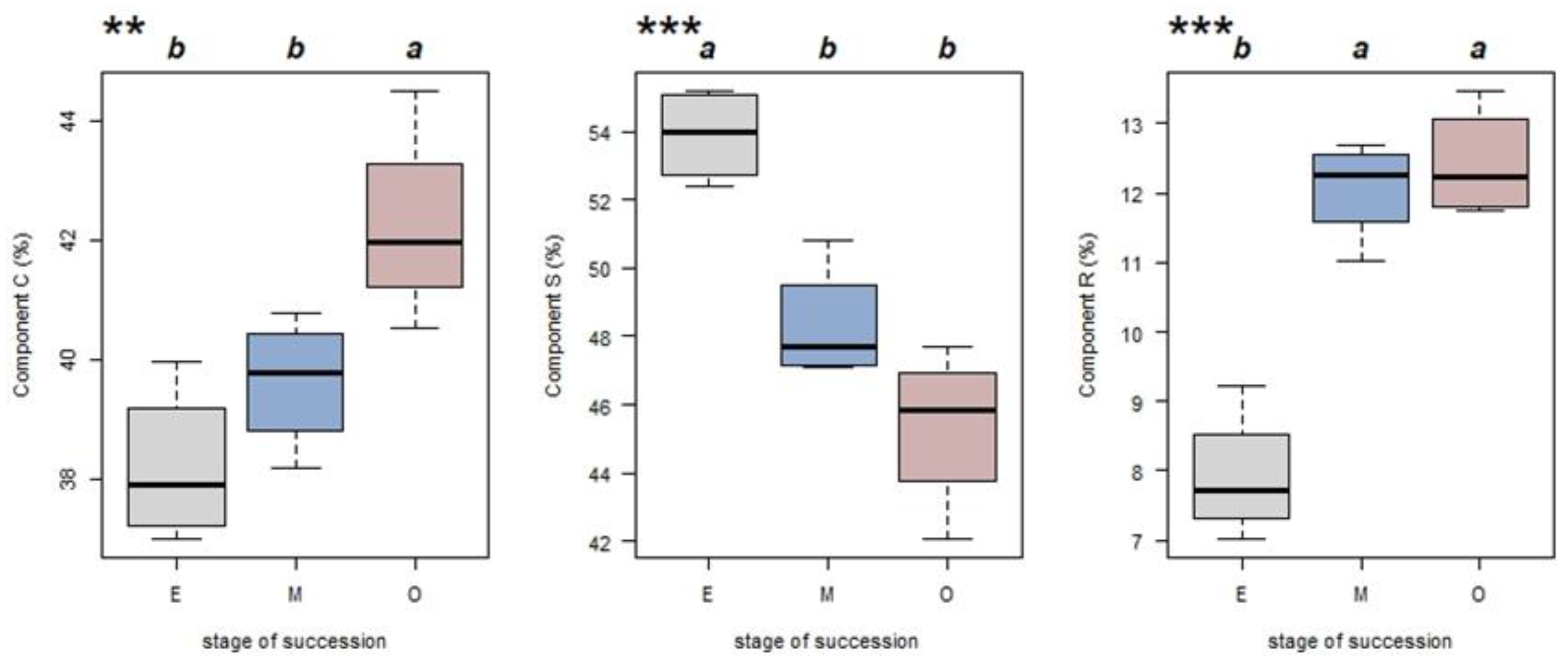
3.3. Effects of Successions on CSR Strategies Composition
4. Discussion
4.1. Dynamic Successional Patterns in Community Ecological Strategy Spectra
4.2. Community-Level Differences in Ecological Strategy Composition with Succession
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Diaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Chauvin, K.M.; Asner, G.P.; Martin, R.E.; Kress, W.J.; Wright, S.J.; Field, C.B. Decoupled dimensions of leaf economic and anti-herbivore defense strategies in a tropical canopy tree community. Oecologia 2018, 186, 765–782. [Google Scholar] [CrossRef] [PubMed]
- DÍAz, S.; Lavorel, S.; McIntyre, S.U.E.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; et al. Plant trait responses to grazing—A global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Guo, W.Y.; van Kleunen, M.; Winter, M.; Weigelt, P.; Stein, A.; Pierce, S.; Pergl, J.; Moser, D.; Maurel, N.; Lenzner, B.; et al. The role of adaptive strategies in plant naturalization. Ecol. Lett. 2018, 21, 1380–1389. [Google Scholar] [CrossRef]
- Niu, S.; Classen, A.T.; Luo, Y. Functional traits along a transect. Funct. Ecol. 2018, 32, 4–9. [Google Scholar] [CrossRef] [Green Version]
- GÓMez-Aparicio, L.; GarcÍA-ValdÉS, R.; RuÍZ-Benito, P.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Chang. Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Cabido, M.R. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties; Wiley: Hoboken, NJ, USA, 2001. [Google Scholar]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant Ecological Strategies: Some Leading Dimensions of Variation between Species. Annu. Rev. Entomol. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Dayrell, R.L.C.; Arruda, A.J.; Pierce, S.; Negreiros, D.; Meyer, P.B.; Lambers, H.; Silveira, F.A.O.; Godoy, O. Ontogenetic shifts in plant ecological strategies. Funct. Ecol. 2018, 32, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the Holy Grail: Using plant functional traits to understand ecological processes. Biol. Rev. Camb Philos. Soc. 2017, 92, 1156–1173. [Google Scholar] [CrossRef] [PubMed]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J.G.; Wilson, P.J.; Hunt, R.; Grime, J.P.; Thompson, K. Allocating C-S-R Plant Functional Types: A Soft Approach to a Hard Problem. Oikos 1999, 85, 282–294. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2016, 529, 204–207. [Google Scholar] [CrossRef] [Green Version]
- Rosado, B.H.P.; Mattos, E.A.; Baltzer, J. On the relative importance of CSR ecological strategies and integrative traits to explain species dominance at local scales. Funct. Ecol. 2017, 31, 1969–1974. [Google Scholar] [CrossRef] [Green Version]
- Negreiros, D.; Le Stradic, S.; Fernandes, G.W.; Rennó, H.C. CSR analysis of plant functional types in highly diverse tropical grasslands of harsh environments. Plant Ecol. 2014, 215, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Cerabolini, B.E.L.; Brusa, G.; Ceriani, R.M.; De Andreis, R.; Luzzaro, A.; Pierce, S. Can CSR classification be generally applied outside Britain? Plant Ecol. 2010, 210, 253–261. [Google Scholar] [CrossRef]
- Kattenborn, T.; Fassnacht, F.E.; Pierce, S.; Lopatin, J.; Grime, J.P.; Schmidtlein, S.; Paruelo, J. Linking plant strategies and plant traits derived by radiative transfer modelling. J. Veg. Sci. 2017, 28, 717–727. [Google Scholar] [CrossRef]
- de Paula, L.F.A.; Negreiros, D.; Azevedo, L.O.; Fernandes, R.L.; Stehmann, J.R.; Silveira, F.A.O. Functional ecology as a missing link for conservation of a resource-limited flora in the Atlantic forest. Biodivers. Conserv. 2015, 24, 2239–2253. [Google Scholar] [CrossRef]
- Hu, G.; Jin, Y.; Liu, J.; Yu, M. Functional diversity versus species diversity: Relationships with habitat heterogeneity at multiple scales in a subtropical evergreen broad-leaved forest. Ecol. Res. 2014, 29, 897–903. [Google Scholar] [CrossRef]
- Huston, M.A.; Smith, T. Plant Succession: Life History and Competition. Am. Nat. 1987, 130, 168–198. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Barba-Escoto, L.; Ponce-Mendoza, A.; García-Romero, A.; Calvillo-Medina, R.P. Plant community strategies responses to recent eruptions of Popocatépetl volcano, Mexico. J. Veg. Sci. 2019, 30, 375–385. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Romero-Pérez, I.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef]
- Malizia, A.; Easdale, T.A.; Grau, H.R. Rapid structural and compositional change in an old-growth subtropical forest: Using plant traits to identify probable drivers. PLoS ONE 2013, 8, e73546. [Google Scholar] [CrossRef] [Green Version]
- Heim, A.; Lundholm, J. Changes in plant community composition and functional plant traits over a four-year period on an extensive green roof. J. Environ. Manag. 2022, 304, 114154. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional traits and niche-based tree community assembly in an amazonian forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodáňová, D. Plant strategies and vegetation processes. Biol. Plant 1981, 23, 254. [Google Scholar] [CrossRef]
- Mouillot, D.; Villeger, S.; Scherer-Lorenzen, M.; Mason, N.W. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS ONE 2011, 6, e17476. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Zang, R.G. Effects of Logging on the Diversity of Lianas in a Lowland Tropical Rain Forest in Hainan Island, South China. Biotropica 2009, 41, 618–624. [Google Scholar] [CrossRef]
- Lu, X.; Zang, R.; Ding, Y.; Huang, J. Changes in biotic and abiotic drivers of seedling species composition during forest recovery following shifting cultivation on Hainan Island, China. Biotropica 2016, 48, 758–769. [Google Scholar] [CrossRef]
- Condit, R. The CTFS and the Standardization of Methodology. In Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer: Berlin/Heidelberg, Germany, 1998; pp. 3–7. [Google Scholar]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Magneville, C.; Loiseau, N.; Albouy, C.; Casajus, N.; Claverie, T.; Escalas, A.; Leprieur, F.; Maire, E.; Mouillot, D.; Villéger, S. mFD: An R package to compute and illustrate the multiple facets of functional diversity. Ecography 2022, 2022. [Google Scholar] [CrossRef]
- Han, X.; Huang, J.; Zang, R. Soil nutrients and climate seasonality drive differentiation of ecological strategies of species in forests across four climatic zones. Plant Soil 2022, 473, 517–531. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Han, X.; Huang, J.; Yao, J.; Xu, Y.; Ding, Y.; Zang, R. Effects of logging on the ecological strategy spectrum of a tropical montane rain forest. Ecol. Indic. 2021, 128, 107812. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Volume 1, pp. 12–21. [Google Scholar]
- Rosenfield, M.F.; Müller, S.C.; Overbeck, G.E. Short gradient, but distinct plant strategies: The CSR scheme applied to subtropical forests. J. Veg. Sci. 2019, 30, 984–993. [Google Scholar] [CrossRef]
- Liu, X.; Swenson, N.G.; Zhang, J.; Ma, K.; Thompson, K. The environment and space, not phylogeny, determine trait dispersion in a subtropical forest. Funct. Ecol. 2013, 27, 264–272. [Google Scholar] [CrossRef]
- Cerabolini, B.E.L.; Pierce, S.; Verginella, A.; Brusa, G.; Ceriani, R.M.; Armiraglio, S. Why are many anthropogenic agroecosystems particularly species-rich? Plant Biosyst. 2014, 150, 550–557. [Google Scholar] [CrossRef]
- Gutiérrez-Girón, A.; Gavilán, R. Plant functional strategies and environmental constraints in Mediterranean high mountain grasslands in central Spain. Plant Ecol. Divers. 2013, 6, 435–446. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Read, Q.D.; Moorhead, L.C.; Swenson, N.G.; Bailey, J.K.; Sanders, N.J.; Fox, C. Convergent effects of elevation on functional leaf traits within and among species. Funct. Ecol. 2014, 28, 37–45. [Google Scholar] [CrossRef]
- Yu, R.; Huang, J.; Xu, Y.; Ding, Y.; Zang, R. Plant Functional Niches in Forests Across Four Climatic Zones: Exploring the Periodic Table of Niches Based on Plant Functional Traits. Front. Plant Sci. 2020, 11, 841. [Google Scholar] [CrossRef]
- Han, X.; Huang, J.; Zang, R. Shifts in ecological strategy spectra of typical forest vegetation types across four climatic zones. Sci. Rep. 2021, 11, 14127. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, N.; Liu, C.; Yang, H.; Li, M.; Yu, G.; Wilcox, K.; Yu, Q.; He, N. C:N:P stoichiometry in China’s forests: From organs to ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Lebrija-Trejos, E.; Perez-Garcia, E.A.; Meave, J.A.; Poorter, L.; Bongers, F. Environmental changes during secondary succession in a tropical dry forest in Mexico. J. Trop. Ecol. 2011, 27, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Lasky, J.R.; Sun, I.F.; Su, S.H.; Chen, Z.S.; Keitt, T.H.; Canham, C. Trait-mediated effects of environmental filtering on tree community dynamics. J. Ecol. 2013, 101, 722–733. [Google Scholar] [CrossRef]
- Dudova, K.V.; Dzhatdoeva, T.M.; Dudov, S.V.; Akhmetzhanova, A.A.; Tekeev, D.K.; Onipchenko, V.G. Competitive Strategy of Subalpine Tall-Grass Species of the Northwestern Caucasus. Mosc. Univ. Biol. Sci. Bull. 2019, 74, 140–146. [Google Scholar] [CrossRef]
- Hou, D.; He, W.; Liu, C.; Qiao, X.; Guo, K. Litter accumulation alters the abiotic environment and drives community successional changes in two fenced grasslands in Inner Mongolia. Econ. Evol. 2019, 9, 9214–9224. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages of Succession | Abbreviation | Interference History | Number of Plots |
|---|---|---|---|
| Early succession | E | 30-year-old secondary forest | 4 |
| Mid-succession | M | 60-year-old secondary forest | 4 |
| Late succession | O | Old growth forest | 4 |
| Early Succession | Mid-Succession | Late Succession |
|---|---|---|
| C C/CR C/CS C/CSR CR CS CS/CSR CSR CR/CSR R/CSR S S/CS S/CSR S/SR SR/CSR | C C/CR C/CS C/CSR CR CR/CSR CS CS/CSR CSR R/CR S S/CS S/CSR SR/CSR | C C/CR C/CS C/CSR CR CR/CSR CS CS/CSR CSR R/CSR S/CS S/CSR S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Wen, Y.; Ji, T.; Zhao, H.; Zang, R.; Lu, X. Ecological Strategy Spectra for Communities of Different Successional Stages in the Tropical Lowland Rainforest of Hainan Island. Forests 2022, 13, 973. https://doi.org/10.3390/f13070973
Chen C, Wen Y, Ji T, Zhao H, Zang R, Lu X. Ecological Strategy Spectra for Communities of Different Successional Stages in the Tropical Lowland Rainforest of Hainan Island. Forests. 2022; 13(7):973. https://doi.org/10.3390/f13070973
Chicago/Turabian StyleChen, Chen, Yabo Wen, Tengyue Ji, Hongxia Zhao, Runguo Zang, and Xinghui Lu. 2022. "Ecological Strategy Spectra for Communities of Different Successional Stages in the Tropical Lowland Rainforest of Hainan Island" Forests 13, no. 7: 973. https://doi.org/10.3390/f13070973
APA StyleChen, C., Wen, Y., Ji, T., Zhao, H., Zang, R., & Lu, X. (2022). Ecological Strategy Spectra for Communities of Different Successional Stages in the Tropical Lowland Rainforest of Hainan Island. Forests, 13(7), 973. https://doi.org/10.3390/f13070973