
Drivers of Native Species Regeneration in the Process of Restoring a Dry Evergreen Forest from Exotic Tree Plantations in Northeastern Thailand


Abstract
:1. Introduction
2. Materials and Methods
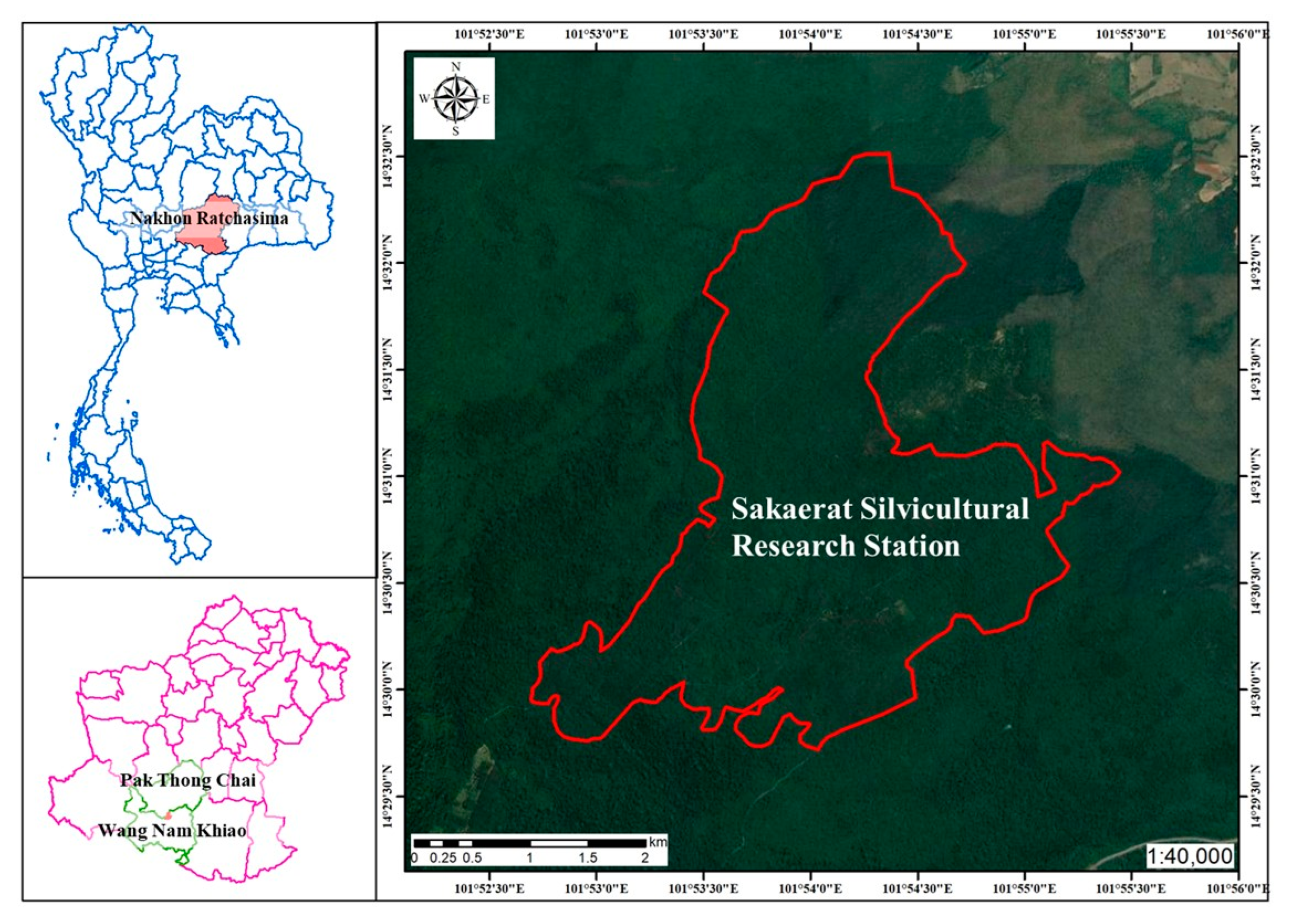
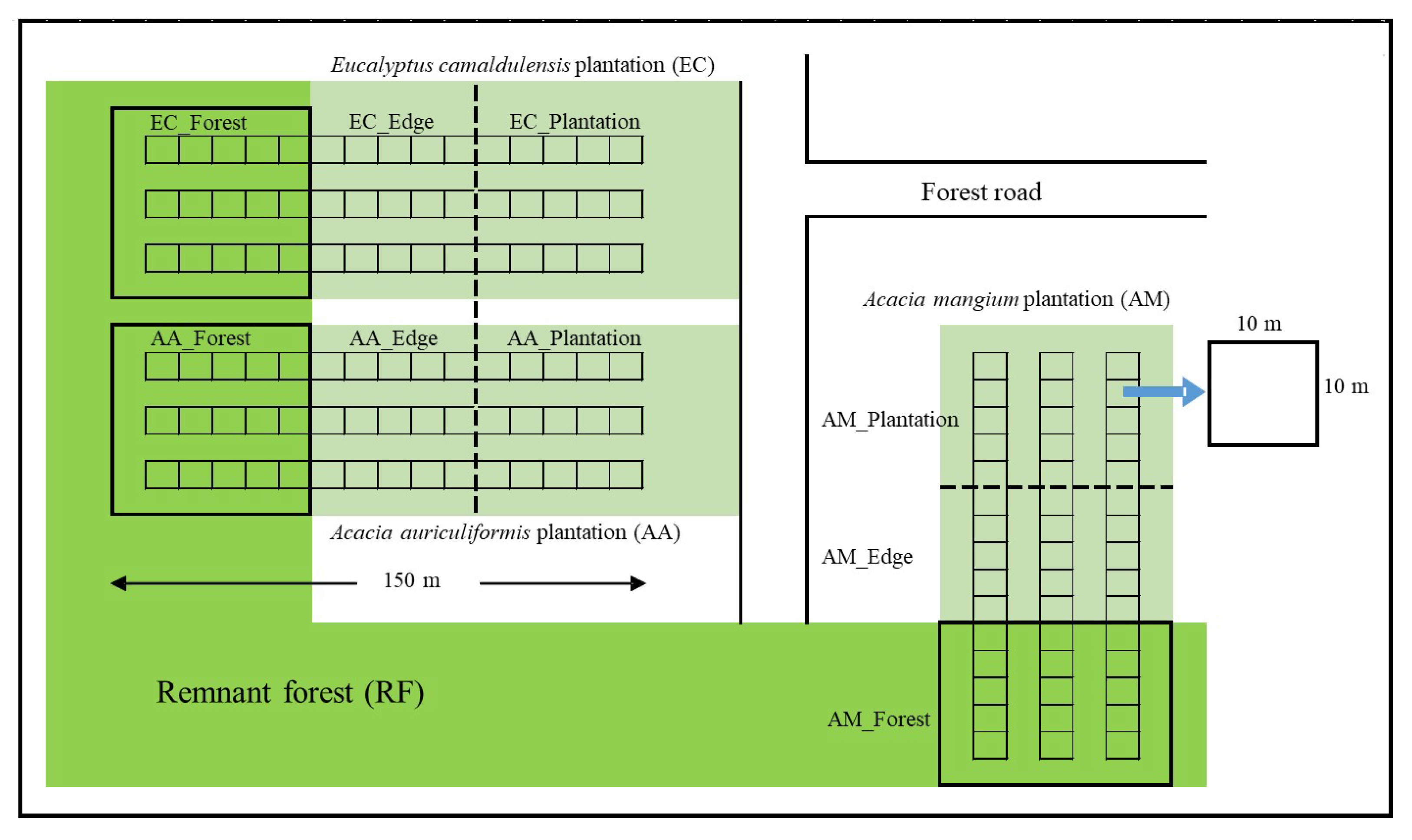
2.1. Study Site
2.2. Environmental Factors
2.3. Data Analysis
3. Results
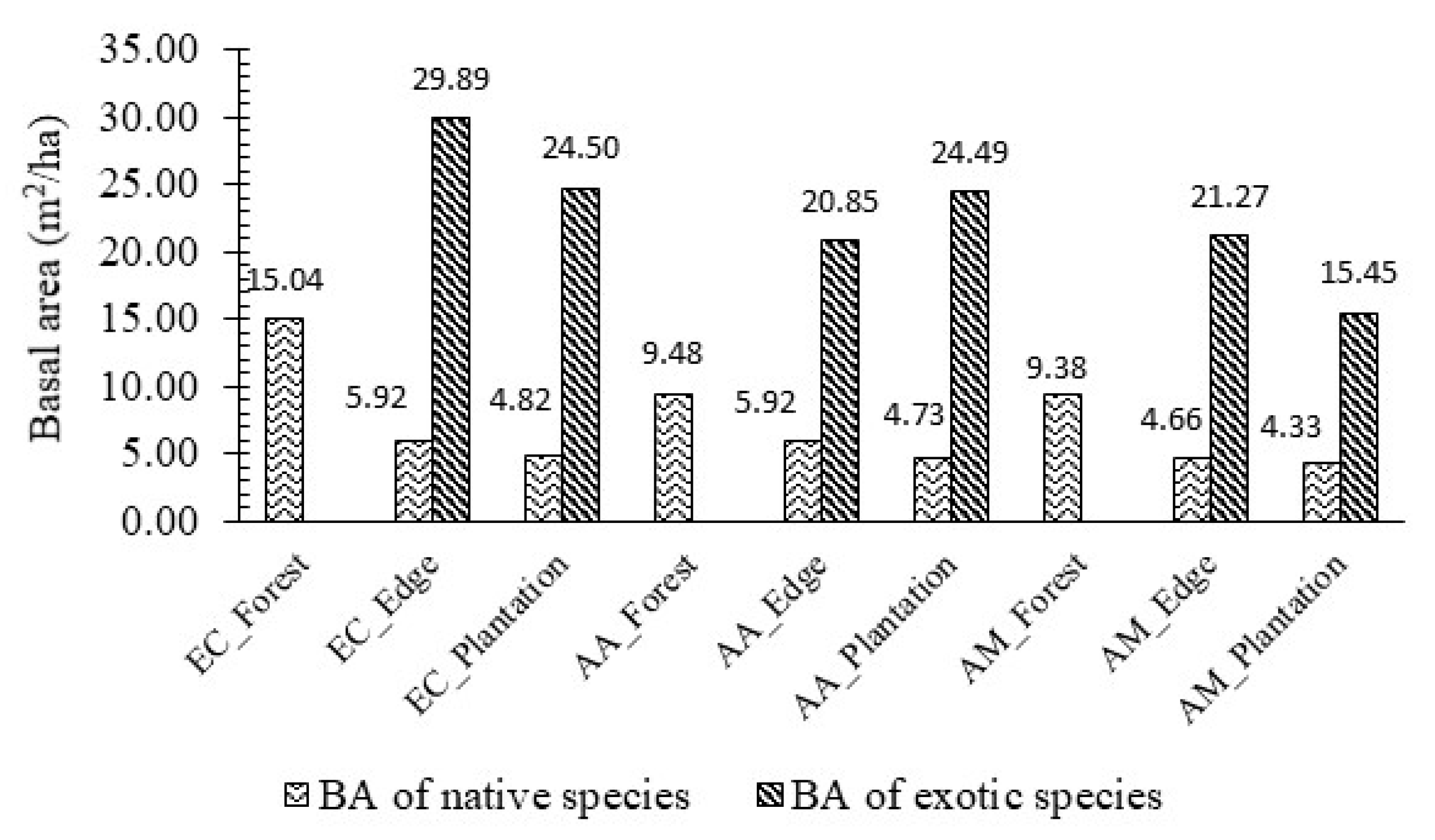
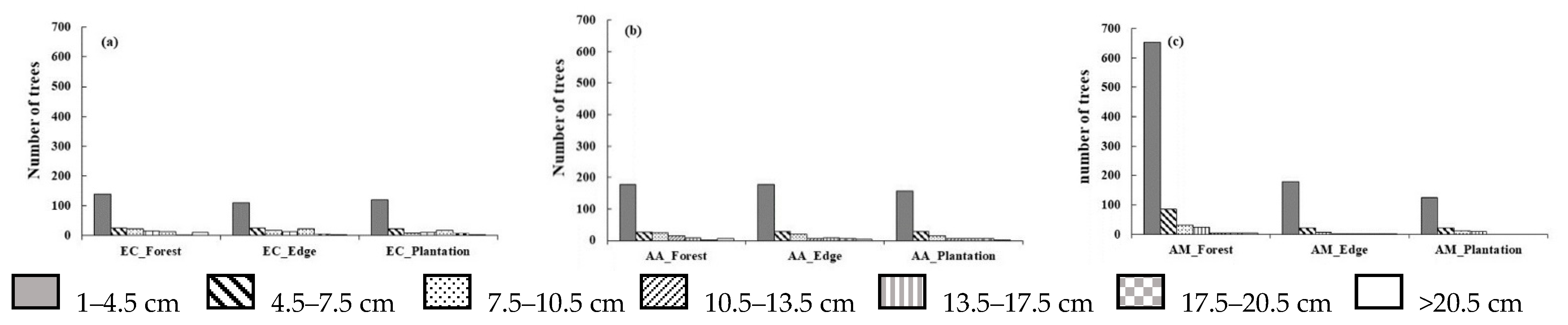
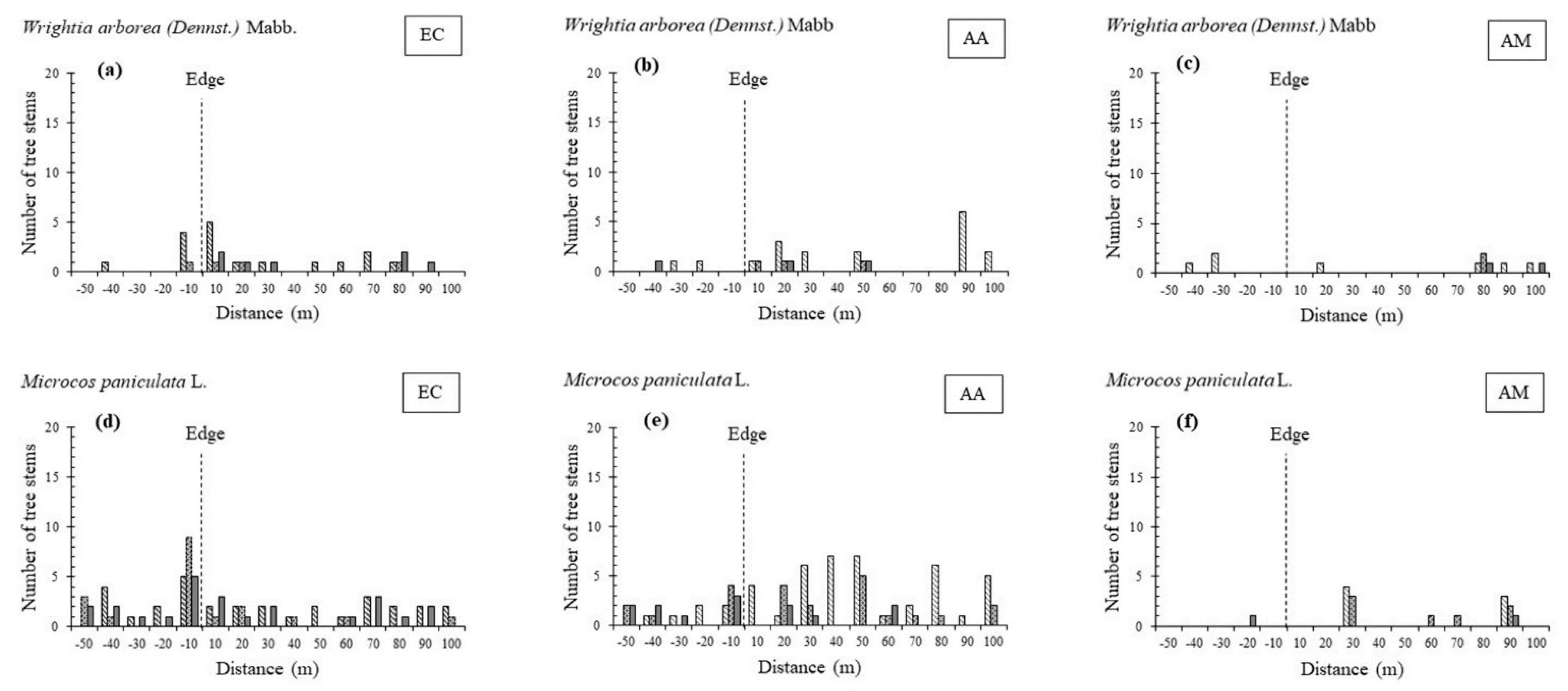
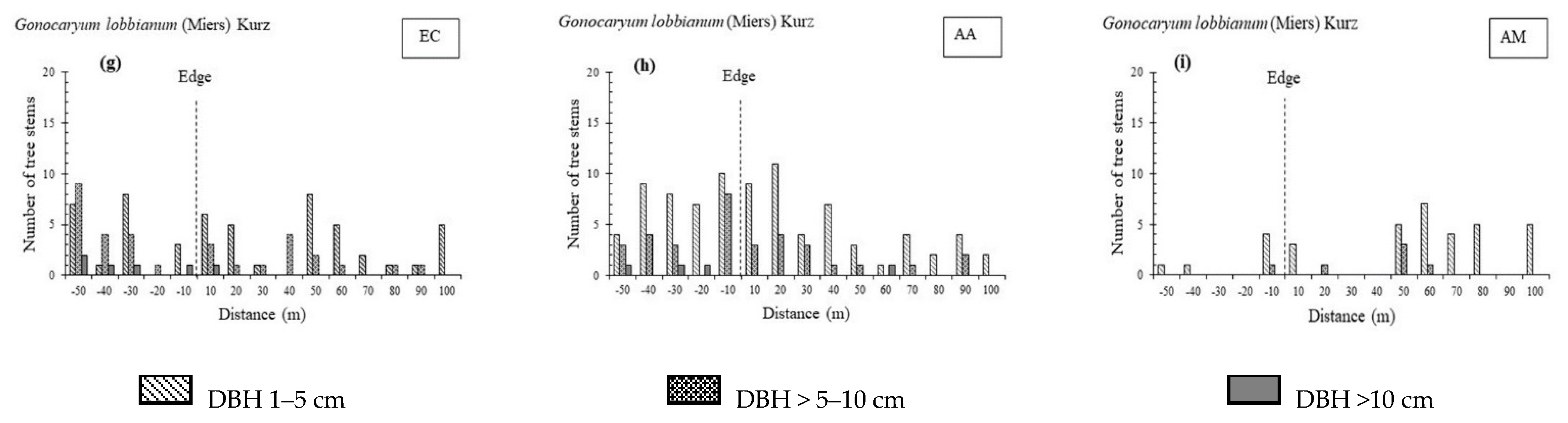
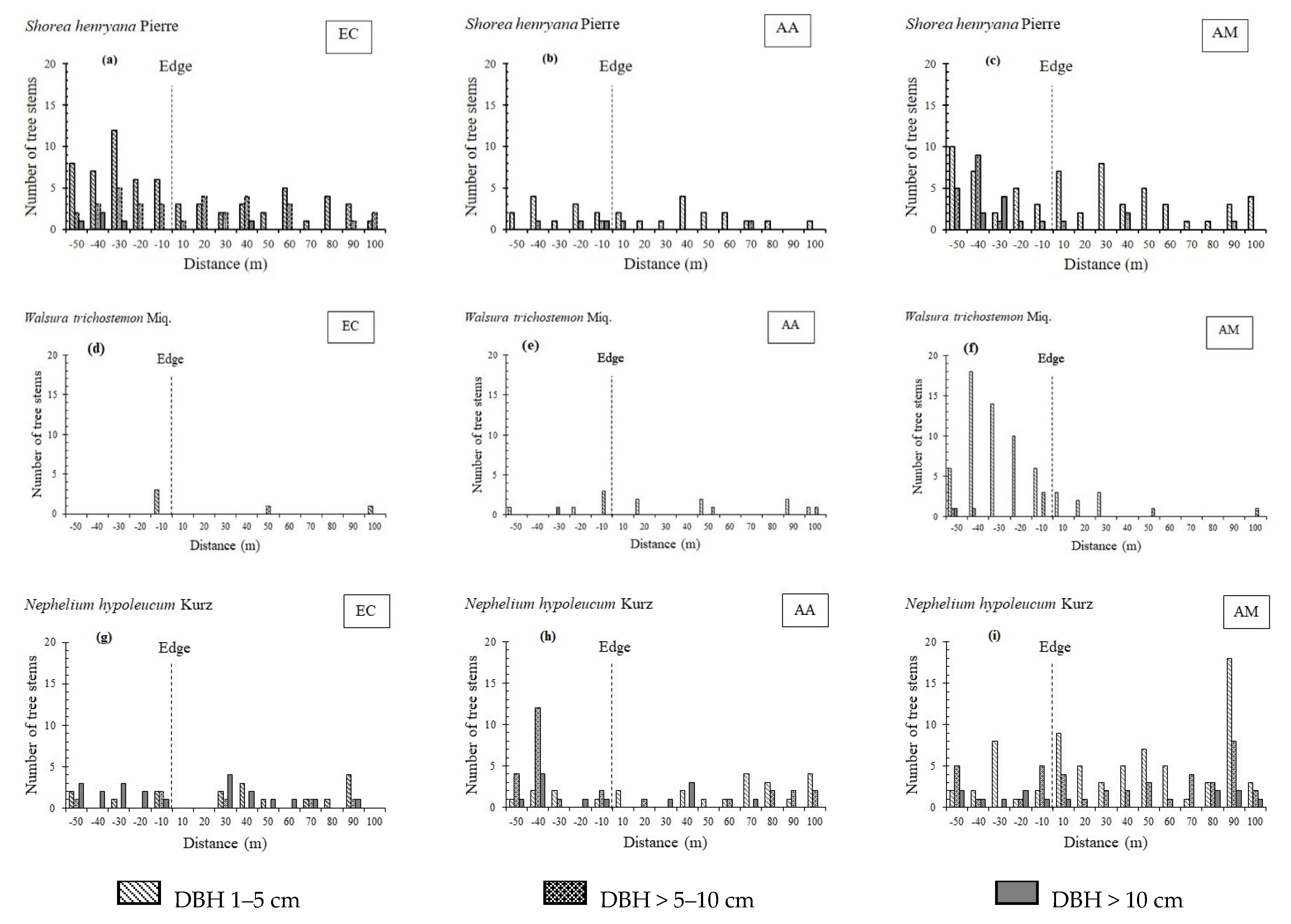
3.1. Stand Structure and Species Composition
3.2. Soil Properties in Exotic Plantation Stands
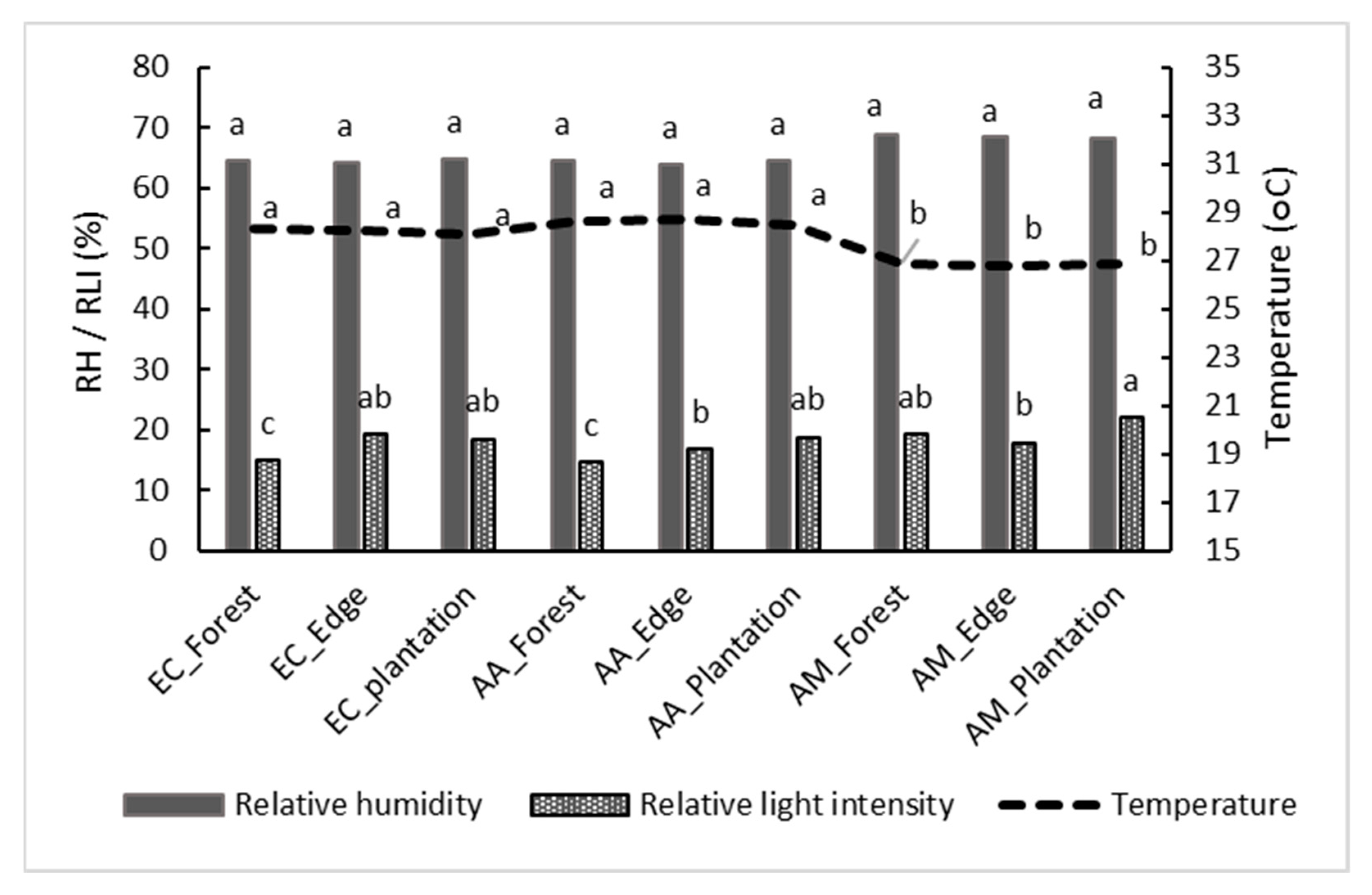
3.3. Environmental Factors in Exotic Plantation Stands
3.4. Relationship between Environmental Factors and Native Tree Distribution
4. Discussion
4.1. Stand Structure and Composition Influencing Understory Vegetation and Environments
4.2. Environmental Factors Driving Natural Regeneration of Native Tree Species
4.3. Natural Regeneration along Forest Remnants to Plantations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | Tree Species | RDo | RF | RD | IVI |
|---|---|---|---|---|---|
| EC_Forest | Lithocarpus elegans (Blume) Hatus. ex Soepadmo. | 30.08 | 5.29 | 3.52 | 38.90 |
| Antidesma acidum Retz. | 5.56 | 7.41 | 21.72 | 34.69 | |
| Shorea henryana Pierre | 7.74 | 7.41 | 12.13 | 27.28 | |
| Nephelium hypoleucum Kurz. | 11.44 | 4.76 | 3.72 | 19.92 | |
| Gonocaryum lobbianum (Miers) Kurz. | 5.90 | 5.82 | 7.83 | 19.54 | |
| EC_Edge | Eucalyptus camaldulensis Dehnh. | 83.45 | 9.26 | 18.63 | 111.35 |
| Clausena guillauminii Tanaka. | 0.82 | 7.41 | 12.22 | 20.44 | |
| Microdesmis caseariifolia Planch. ex Hook. | 0.30 | 8.02 | 8.49 | 16.82 | |
| Antidesma acidum Retz. | 0.66 | 8.02 | 7.25 | 15.93 | |
| Ixora cibdela Craib | 0.15 | 6.17 | 8.28 | 14.60 | |
| EC_Plantation | Eucalyptus camaldulensis Dehnh. | 83.69 | 10.42 | 16.80 | 110.90 |
| Clausena guillauminii Tanaka. | 1.45 | 9.72 | 22.67 | 33.84 | |
| Ixora cibdela Craib | 0.11 | 9.03 | 10.13 | 19.27 | |
| Microcos tomentosa Sm. | 4.23 | 7.64 | 2.67 | 14.54 | |
| Gonocaryum lobbianum (Miers) Kurz | 0.43 | 7.64 | 4,53 | 12.60 | |
| AA_Forest | Gonocaryum lobbianum (Miers) Kurz. | 9.93 | 5.80 | 10.59 | 26.32 |
| Nephelium hypoleucum Kurz. | 12.95 | 4.46 | 5.38 | 22.79 | |
| Antidesma acidum Retz. | 6.23 | 6.70 | 9.41 | 22.34 | |
| Microcos tomentosa Sm. | 12.24 | 5.36 | 3.53 | 21.12 | |
| Microdesmis caseariifolia Planch. ex Hook. | 1.94 | 6.70 | 10.76 | 19.39 | |
| AA_Edge | Acacia auriculiformis A. Cunn. ex Benth. | 76.99 | 6.38 | 5.51 | 88.88 |
| Clausena guillauminii Tanaka. | 4.29 | 7.45 | 23.81 | 35.54 | |
| Ixora cibdela Craib | 0.39 | 7.98 | 11.76 | 20.13 | |
| Antidesma acidum Retz. | 1.58 | 6.91 | 8.78 | 17.28 | |
| Microcos tomentosa Sm. | 3.17 | 6.91 | 5.80 | 15.89 | |
| AA_Plantation | Acacia auriculiformis A. Cunn. ex Benth. | 83.82 | 9.74 | 8.63 | 102.18 |
| Clausena guillauminii Tanaka. | 4.96 | 9.74 | 32.57 | 47.27 | |
| Hopea odorata Roxb. | 1.69 | 9.74 | 13.38 | 24.82 | |
| Microdesmis caseariifolia Planch. ex Hook. | 0.59 | 7.14 | 4.75 | 12.48 | |
| Antidesma acidum Retz. | 0.47 | 7.14 | 4.58 | 12.19 | |
| AM_Forest | Alchornea rugosa Muell. Arg. | 1.77 | 5.93 | 15.88 | 23.59 |
| Shorea henryana Pierre | 11.60 | 5.53 | 6.02 | 23.15 | |
| Dipterocarpus turbinatus C. F. Gaertn. | 9.64 | 5.14 | 7.34 | 22.12 | |
| Pterocymbium tinctorium (Blanco) Merr. | 4.88 | 5.53 | 10.35 | 20.76 | |
| Nephelium hypoleucum Kurz | 10.35 | 4.74 | 4.21 | 19.31 | |
| AM_Edge | Acacia mangium Willd. | 81.86 | 4.72 | 5.04 | 91.61 |
| Alchornea rugosa Muell. Arg. | 0.60 | 5.91 | 18.48 | 24.99 | |
| Clausena guillauminii Tanaka. | 2.34 | 5.91 | 8.91 | 17.16 | |
| Dipterocarpus turbinatus C. F. Gaertn. | 1.59 | 3.15 | 8.01 | 12.75 | |
| Nephelium hypoleucum Kurz | 2.28 | 4.33 | 5.43 | 12.04 | |
| AM_Plantation | Acacia mangium Willd. | 78.14 | 6.09 | 4.08 | 88.31 |
| Alchornea rugosa Muell. Arg. | 0.79 | 5.65 | 19.97 | 26.1 | |
| Nephelium hypoleucum Kurz. | 4.66 | 4.72 | 8.02 | 17.46 | |
| Clausena guillauminii Tanaka | 1.64 | 5.65 | 7.41 | 14.71 | |
| Mallotus philippensis Mull. Arg. | 2.85 | 3.48 | 4.69 | 11.01 |
References
- FAO. Global Forest Resources Assessment 2020 Key Findings; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Montagnini, F.; Jordan, C.F. Tropical Forest Ecology: The Basis for Conservation and Management; Springer: Berlin, Germany, 2005. [Google Scholar]
- Parrotta, J.A. Productivity, nutrient cycling, and succession in single and mixed species plantations of Casuarina equisetifolia, Eucalyptus robusta, and Leucaena leucocephala in Puerto Rico. For. Ecol. Manag. 1999, 124, 45–77. [Google Scholar] [CrossRef]
- Tang, J.W.; Cao, M.; Zhang, J.H.; Li, M.H. Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: A 10 year study. Plant Soil 2010, 335, 271–288. [Google Scholar] [CrossRef]
- Bohre, P.; Chaubey, O.P. Restoration of degraded lands through plantation forests. Glob. J. Sci. Front. Res. 2014, 14, 19–27. [Google Scholar]
- Silva, L.N.; Freer-Smith, P.; Madsen, P. Production, restoration, mitigation: A new generation of plantations. New For. 2019, 50, 153–168. [Google Scholar] [CrossRef]
- Feyera, S.; Beck, E.; Lüttge, U. Exotic trees as nurse-trees for the regeneration of natural tropical forests. Trees 2002, 16, 245–249. [Google Scholar] [CrossRef]
- Martpalakorn, M. Tree Species Trials on Mined Spoils at Amphoe Takuapa, Changwat Phangnga. Master’s Thesis, Kasetsart University, Bangkok, Thailand, 16 March 1990. [Google Scholar]
- Parrotta, J.A. The role of plantation forests in rehabilitating degraded tropical ecosystems. Agric. Ecosyst. Environ. 1992, 41, 115–133. [Google Scholar] [CrossRef]
- Sugimoto, M.; Ohta, S.; Ansori, S.; Arisman, H. Nutrient dynamics via litterfall and litter decomposition on the forest floor of an Acacia mangium Willd. stand in Sumatra. Tropics 2013, 22, 67–81. [Google Scholar] [CrossRef]
- Kremer, K.N.; Bauhus, J. Drivers of native species regeneration in the process of restoring natural forests from mono-specific, even-aged tree plantations: A quantitative review. Restor. Ecol. 2020, 28, 1074–1086. [Google Scholar] [CrossRef]
- Parrotta, J.A. Influence of overstory composition on understory colonization by native species in plantations on a degraded tropical site. J. Veg. Sci. 1995, 6, 627–636. [Google Scholar] [CrossRef]
- Loumeto, J.J.; Huttel, C. Understory vegetation in fast-growing tree plantations on savanna soils in Congo. For. Ecol. Manag. 1997, 99, 65–81. [Google Scholar] [CrossRef]
- Senbeta, F.; Teketay, D.; Naslund, B. Native woody species regeneration in exotic tree plantations at Munessa-Shashemene Forest, southern Ethiopia. New For. 2002, 24, 131–145. [Google Scholar] [CrossRef]
- Forbes, A.S.; Norton, D.A.; Carswell, F.E. Opportunities and limitations of exotic Pinus radiata as a facilitative nurse for New Zealand indigenous forest restoration. N. Z. J. For. Sci. 2019, 49, 6. [Google Scholar] [CrossRef]
- Keenan, R.; Lamb, D.; Woldring, O.; Irvine, T.; Jensen, R. Restoration of plant biodiversity beneath tropical tree plantations in Northern Australia. For. Ecol. Manag. 1997, 99, 117–131. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.; Xia, H.; Zou, B.; Li, N.; Liu, J.; Zhu, W. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci. Plant Nutr. 2010, 56, 297–306. [Google Scholar] [CrossRef]
- Martins, K.G.; Marques, M.M.C.; dos Santos, E.; Marques, R. Effects of soil conditions on the diversity of tropical forests across a successional gradient. For. Ecol. Manag. 2015, 349, 4–11. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Q.R.; Fan, W.; Song, G.H. The relationship between secondary forest and environmental factors in the southern Taihang Mountains. Sci. Rep. 2017, 7, 16431. [Google Scholar] [CrossRef]
- Vahdati, F.B.; Mehrvarz, S.S.; Dey, D.C.; Naqinezhad, A. Environmental factors–ecological species group relationships in the Surash lowland-mountain forests in northern Iran. Nord. J. Bot. 2016, 35, 240–250. [Google Scholar] [CrossRef]
- Thammanu, S.; Marod, D.; Han, H.; Bhusal, N.; Asanok, L.; Ketdee, P.; Gaewsingha, N.; Lee, S. The influence of environmental factors on species composition and distribution in a community forest in Northern Thailand. J. For. Res. 2021, 32, 649–662. [Google Scholar] [CrossRef]
- Duan, W.; Ren, H.; Fu, S.; Wang, J.; Zhang, J.; Yang, L.; Huang, C. Community Comparison and Determinant Analysis of Understory Vegetation in Six Plantations in South China. Restor. Ecol. 2010, 18, 206–214. [Google Scholar] [CrossRef]
- Asanok, L.; Taweesuk, R.; Papakian, K. Woody species colonization along edge-interior gradients of deciduous forest remnants in the Mae Khum Mee Watershed Northern Thailand. Int. J. For. Res. 2020, 2020, 5867376. [Google Scholar] [CrossRef]
- Van, D.T.; Lee, D.K.; Van, T.H. Rehabilitation of native tree species in the forest plantations and denuded hills of Namlau commune in Sonla Province, Vietnam. For. Sci. Technol. 2005, 1, 51–58. [Google Scholar] [CrossRef]
- Sakai, A.; Visaratana, T.; Vacharangkura, T.; Ishizuka, M.; Nakamura, S. Growth performances of three indigenous tree species planted in a mature Acacia mangium plantation with different canopy openness under a tropical monsoon climate. Jpn. Agric. Res. Q. JARQ 2011, 45, 317–326. [Google Scholar] [CrossRef]
- Sakurai, K.; Kozasa, S.; Yuasa, T.; Puriyakorn, B.; Preechapanya, P.; Tanpibal, V.; Muangnil, K.; Prachaiyo, B. Changes in soil properties after land degradation associated with various human activities in Thailand. Soil Sci. Plant Nutr. 1996, 42, 81–92. [Google Scholar] [CrossRef]
- Kongdam, P.; Pimprasit, S.; Wachrinrat, C.; Marod, D. Forest structure and species composition in restoration by Teak plantation at Jedkhod—Pongkhonsao Natural Study and Ecotourism Center, Kheang Khoi District, Saraburi Province. Thai J. For. 2016, 35, 11–23. [Google Scholar]
- Hermhuk, S.; Sungpalee, W.; Sri-Ngernyuang, K.; Satienperakul, K. Natural regeneration of native plant species in restoration forest by Eucalyptus camaldulensis at Khun Han Plantation, Si Sa Ket province. Thai J. For. 2019, 38, 66–80. [Google Scholar]
- Phumphuang, W.; Marod, D.; Sungkaew, S.; Thinkapaeng, S. Forest dynamics and tree distribution patterns in dry evergreen forest, northeastern, Thailand. Nat. Resour. Environ. 2018, 16, 58–67. [Google Scholar]
- Bullock, J. Plants. In Ecological Census Techniques: A Handbook; Sutherland, W.J., Ed.; Cambridge University Press: Cambridge, UK, 1996; pp. 111–138. [Google Scholar]
- Bunyavejchewin, S. Structure and dynamics in seasonal dry evergreen forest in northeastern Thailand. J. Veg. Sci. 1999, 10, 787–792. [Google Scholar] [CrossRef]
- Frazer, G.W.; Trofymow, J.A.; Lertzman, K.P. A Method for Estimating Canopy Openness, Effective Leaf Area Index, and Photosynthetically Active Photon Flux Density Using Hemispherical Photography and Computerized Image Analysis Techniques; The Pacific Forestry Centre: Victoria, BC, Canada, 1997. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis-Advanced Course; Parallel Press: Madison, WI, USA, 1965. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–37. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis, Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America, Inc. and American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. The Interrelations of Certain Analytic and Synthetic Phytosociological Characters. Ecology 1950, 31, 434–455. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community ecology package. R package version 2.2-1, 2016. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 12 October 2020).
- McCune, B.; Mefford, M.J. PC-ORD Version 5.10: Multivariate Analysis of Ecological Data; MjM Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Lameira, L.P.; Ferreira, F.C.G.; Filardi, R.A.E.; Queiroz, J.M.; Sansevero, J.B.B. Plant-canopy effects on natural regeneration in sites under restoration: Do tree species matter? Floresta e Ambiente 2019, 26, e20180398. [Google Scholar] [CrossRef]
- Fisher, R.F.; Binkley, D. Ecology and Management of Forest Soils, 3rd ed.; John Wiley and Sons: New York, NY, USA, 2000. [Google Scholar]
- Wongprom, J.; Poolsiri, R.; Diloksumpun, S.; Ngernsaengsaruay, C. Soil properties and tree composition in a 27-year-old Acacia mangium Willd. Plantation on abandoned mining area at Phangnga Forestry Research Station. BIOTROPIA 2020, 27, 125–133. [Google Scholar]
- Niamrat, W.; Marod, D. Seedling establishment of climax species under the Eucalyptus plantations and open areas. Thai J. For. 2005, 24, 35–47. [Google Scholar]
- Ali, A.; Dai, D.; Akhtar, K.; Teng, M.; Yan, Z.; Urbina-Cardona, N.; Mullerova, J.; Zhou, Z. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 2019, 20, e00775. [Google Scholar] [CrossRef]
- Wongprom, J.; Poolsiri, R.; Diloksumpun, S.; Ngernsaengsaruay, C.; Tansakul, S.; Chandaeng, W. Litterfall, litter decomposition and nutrient return of rehabilitated mining areas and natural forest in Phangnga forestry research station, southern Thailand. BIOTROPIA 2022, 29, 74–85. [Google Scholar]
- Urairak, K.; Poolsiri, R.; Kaitpraneet, S. Soil properties below exotic tree plantations at the Saithong Silvicultural Research Station in Prachuap Khirikhan province, Thailand. BIOTROPIA 2020, 27, 171–178. [Google Scholar]
- Zhao, Z.; Shahrour, I.Z.; Bai, Z.; Fan, W.; Feng, L.; Li, H. Soil development in opencast coal mine spoils reclaimed for 1–13 years in the West-Northern Loess Plateau of China. Eur. J. Soil Biol. 2013, 55, 40–46. [Google Scholar] [CrossRef]
- Bernhard-Reversat, F. Nitrogen cycling in tree plantations grown on a poor sandy savanna soil in Congo. Appl. Soil Ecol. 1996, 4, 161–172. [Google Scholar] [CrossRef]
- Zahid, D.M.; Shah, F.R.; Majeed, A. Planting Eucalyptus camaldulensis in arid environment is it useful species under water deficit system. Pak. J. Bot. 2010, 42, 1733–1744. [Google Scholar]
- Goosem, S.; Tuckerm, N.I.J. Repairing the Rainforest, 2nd ed.; Wet Tropics Management Authority and Biotropica Australia Pty Ltd.: Cairns, QLD, Australia, 2013. [Google Scholar]
- Sringernyuang, K.; Chai-Udom, K.; Sungpalee, W.; Kanzaki, M.; Itoh, A. The seedling survivorship of two climax Lauraceae species in tropical montane, Thailand. In Proceedings of the Thai Forest Ecology Research Network: Ecology Knowledge for Restoration, Chiang Mai, Thailand, 24–26 January 2013; pp. 25–38. [Google Scholar]
- Marod, D.; Panmongkol, A.; Sangkaew, S.; Jingjai, A. Influences of environmental factors on tree distribution of lower montane evergreen forest at Doi Sutep-Pui Natural Park, Chiang Mai Province. Thai J. For. 2014, 33, 22–33. [Google Scholar]
- Marod, D.; Kutintara, U.; Tanaka, H.; Nakashizuka, T. The effects of drought and fire in seed and seedling dynamics in a tropical seasonal forest in Thailand. Plant Ecol. 2002, 161, 41–57. [Google Scholar] [CrossRef]
- Asanok, L.; Marod, D.; Duengkae, P.; Pranmongkol, U.; Kurokawa, H.; Aiba, M.; Katabuchi, M.; Nakashizuka, T. Relationships between functional traits and the ability of forest tree species to reestablish in secondary forest and enrichment plantations in the uplands of northern Thailand. For. Ecol. Manag. 2013, 296, 9–23. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Fensham, R.; Buckley, Y.M. Restoration thinning accelerates structural development and carbon sequestration in an endangered Australian ecosystem. J. Appl. Ecol. 2010, 47, 681–691. [Google Scholar] [CrossRef]
- Wongprom, J. Growth performance of dipterocarp species planted on abandoned mining area in southern Thailand. BIOTROPIA 2020, 27, 115–124. [Google Scholar]
- Ruiz-Jaen, M.C.; Aide, T.M. Restoration success: How is it being measured? Restor. Ecol. 2005, 13, 569–577. [Google Scholar] [CrossRef]
- McConkey, K.R.; Prasad, S.; Corlett, R.T.; Campos-Arceiz, A.; Brodie, J.F.; Rogers, H.; Santamaria, L. Seed dispersal in changing landscapes. Biol. Conserv. 2012, 146, 1–13. [Google Scholar] [CrossRef]
- Sansevero, J.B.B.; Prieto, P.V.; de Moraes, L.F.D.; Rodrigues, P.J.F.P. Natural regeneration in plantations of native trees in lowland Brazilian Atlantic forest: Community structure, diversity, and dispersal syndromes. Restor. Ecol. 2011, 19, 379–389. [Google Scholar] [CrossRef]
- Koonkhunthod, N.; Sakurai, K.; Tanaka, S. Composition and diversity of woody regeneration in a 37-year-old teak (Tectona grandis L.) plantation in Northern Thailand. For. Ecol. Manag. 2007, 247, 246–254. [Google Scholar] [CrossRef]
- Hörnberg, G.; Ohlson, M.; Zackrisson, O. Stand dynamics, regeneration patterns and long-term continuity in boreal old-growth Picea abies swamp-forests. J. Veg. Sci. 1995, 6, 291–298. [Google Scholar] [CrossRef]









| Stand | Number of Species | Basal Area (m2/ha) | Tree Density (stem/ha) | Shannon–Wiener Index | Sorensen Similarity Index |
|---|---|---|---|---|---|
| EC_Forest | 27.00 ± 1.73 bcde | 15.04 ± 3.81 c | 3406.67 ± 81.92 cd | 2.69 ± 0.03 abc | 100.00 a |
| EC_Edge | 21.33 ± 0.33 de | 35.81 ± 3.20 a | 3220.00 ± 264.58 cd | 2.53 ± 0.06 bcd | 64.26 ± 1.24 cd |
| EC_Plantation | 20.00 ± 3.79 e | 29.55 ± 4.04 ab | 2500.00 ± 726.91 d | 2.35 ± 0.19 cd | 58.28 ± 4.79 d |
| AA_Forest | 28.67 ± 1.45 abcd | 9.48 ± 1.73 c | 4013.33 ± 544.10 bc | 2.84 ± 0.13 ab | 100.00 a |
| AA_Edge | 24.33 ± 1.86 cde | 26.78 ± 6.20 ab | 4480.00 ± 242.49 abc | 2.55 ± 0.10 bcd | 67.55 ± 0.45 bc |
| AA_Plantation | 22.33 ± 2.33 de | 29.23 ± 1.48 ab | 3786.67 ± 363.75 bcd | 2.23 ± 0.08 d | 68.10 ± 3.84 bc |
| AM_Forest | 35.33 ± 1.20 a | 9.38 ± 0.40 c | 5566.67 ± 335.13 a | 2.92 ± 0.03 a | 100.00 a |
| AM_Edge | 34.00 ± 2.08 ab | 25.92 ± 2.92 ab | 5113.33 ± 81.10 ab | 2.87 ± 0.09 ab | 73.66 ± 3.02 b |
| AM_Plantation | 32.00 ± 4.16 abc | 19.78 ± 3.50 bc | 4406.67 ± 666.57 abc | 2.86 ± 0.18 ab | 65.97 ± 0.68 c |
| F-value (Species) | 19.37 ** | 15.80 ** | 16.28 * | 4.54 ns | 1.03 ns |
| F-value (Stand within species) | 3.73 * | 27.29 ** | 2.60 ns | 10.88 ** | 1.79 ns |
| F-value (Species * Stand) | 2.23 ns | 0.84 ns | 0.36 ns | 2.38 ns | 0.99 ns |
| Stand | Soil Texture | Soil Particle (%) | Bulk Density (g m−3) | ||
|---|---|---|---|---|---|
| Sand | Silt | Clay | |||
| Topsoil (Soil depth 0–10 cm) | |||||
| EC_Forest | Sandy loam | 61.07 a | 19.67 b | 19.27 e | 2.33 |
| EC_Edge | Sand clay loam | 59.73 a | 16.33 b | 23.93 cde | 2.15 |
| EC_Plantation | Sand clay loam | 56.57 a | 18.00 b | 25.43 bc | 2.27 |
| AA_Forest | Sandy clay loam | 54.40 a | 20.33 b | 25.27 bc | 2.34 |
| AA_Edge | Sandy clay loam | 60.40 a | 16.33 b | 23.27 cde | 2.39 |
| AA_Plantation | Sandy clay loam | 63.40 a | 15.50 b | 21.10 de | 2.40 |
| AM_Forest | Clay loam | 42.40 b | 29.33 a | 28.27 bc | 2.34 |
| AM_Edge | Clay loam | 39.73 b | 30.67 a | 29.60 ab | 2.36 |
| AM_Plantation | Clay loam | 35.90 b | 30.67 a | 33.43 a | 2.35 |
| F-value (Species) | 373.14 ** | 39.72 ** | 25.19 ** | 4.04 ns | |
| F-value (Stand within species) | 0.27 ns | 0.37 ns | 1.67 ns | 0.36 ns | |
| F-value (Species * Stand) | 1.27 ns | 0.47 ns | 4.24 * | 0.54 ns | |
| Subsoil (Soil depth 10–30 cm) | |||||
| EC_Forest | Sandy clay loam | 55.73 a | 18.00 c | 19.27 e | 2.43 |
| EC_Edge | Sandy clay loam | 59.40 a | 15.00 c | 23.93 cde | 2.26 |
| EC_Plantation | Sandy clay loam | 54.07 a | 16.83 c | 25.43 bcd | 2.32 |
| AA_Forest | Sandy clay loam | 51.07 a | 16.67 c | 25.27 bcd | 2.31 |
| AA_Edge | Sandy clay loam | 55.73 a | 17.67 c | 23.27 cde | 2.35 |
| AA_Plantation | Sandy clay loam | 58.07 a | 18.50 c | 21.10 de | 2.41 |
| AM_Forest | Clay loam | 42.07 b | 24.67 b | 28.27 bc | 2.40 |
| AM_Edge | Clay loam | 35.73 bc | 28.00 ab | 29.60 ab | 2.31 |
| AM_Plantation | Clay loam | 31.57 c | 30.67 a | 33.43 a | 2.26 |
| F-value (Species) | 59.38 ** | 73.90 ** | 22.44 ** | 3.51 ns | |
| F-value (Stand within species) | 2.42 ns | 0.82 ns | 0.35 ns | 0.13 ns | |
| F-value (Species * Stand) | 0.98 ns | 1.23 ns | 1.51 ns | 2.20 ns | |
| Stand | pH | Exchangeable (mg kg−1) | Available P (mg kg−1) | Total N (%) | OM (%) | ||
|---|---|---|---|---|---|---|---|
| K | Ca | Mg | |||||
| Topsoil (Soil depth 0–10 cm) | |||||||
| EC_Forest | 4.1 | 63.47 ab | 165.34 | 73.94 ab | 12.18 | 0.16 | 3.29 c |
| EC_Edge | 3.9 | 58.54 b | 111.68 | 46.51 ab | 11.63 | 0.13 | 3.14 c |
| EC_Plantation | 3.8 | 71.27 ab | 134.10 | 47.22 ab | 13.70 | 0.14 | 3.94 c |
| AA_Forest | 3.8 | 70.31 ab | 94.03 | 46.91 ab | 10.76 | 0.16 | 8.45 ab |
| AA_Edge | 3.7 | 50.11 b | 63.51 | 22.83 b | 13.96 | 0.14 | 8.97 a |
| AA_Plantation | 3.7 | 55.05 b | 47.65 | 19.77 b | 10.00 | 0.13 | 8.07 ab |
| AM_Forest | 4.1 | 56.28 b | 214.91 | 44.91 ab | 10.58 | 0.17 | 3.70 c |
| AM_Edge | 3.6 | 99.61 ab | 131.53 | 80.41 a | 10.07 | 0.15 | 8.81 a |
| AM_Plantation | 3.8 | 109.5 a | 167.24 | 95.22 a | 11.07 | 0.17 | 6.41 b |
| F-value (Species) | 3.59 ns | 2.29 ns | 3.90 ns | 10.37 * | 0.41 ns | 1.82 ns | 28.32 ** |
| F-value (Stand within species) | 1.55 ns | 0.25 ns | 0.59 ns | 0.06 ns | 0.97 ns | 2.50 ns | 5.15 * |
| F-value (Species * Stand) | 0.14 ns | 1.39 ns | 0.95 ns | 0.91 ns | 0.71 ns | 0.68 ns | 3.77 * |
| Subsoil (Soil depth 10–30 cm) | |||||||
| EC_Forest | 3.9 bcd | 28.45 b | 41.63 b | 28.16 b | 9.64 ab | 0.09 | 2.15 b |
| EC_Edge | 3.8 bcd | 30.72 b | 17.57 b | 10.85 b | 9.05 abc | 0.09 | 1.84 b |
| EC_Plantation | 3.8 bcd | 40.92 ab | 22.88 b | 18.17 b | 10.65 a | 0.09 | 1.81 b |
| AA_Forest | 3.7 d | 31.77 b | 20.79 b | 12.73 b | 6.69 bc | 0.10 | 7.25 a |
| AA_Edge | 3.7 d | 37.04 ab | 29.59 b | 13.41 b | 6.85 bc | 0.12 | 7.33 a |
| AA_Plantation | 3.8 cd | 29.77 b | 15.82 b | 7.76 b | 7.67 abc | 0.10 | 6.82 a |
| AM_Forest | 4.1 a | 34.78 b | 58.04 b | 71.49 a | 5.78 c | 0.10 | 1.49 b |
| AM_Edge | 4.0 a | 72.40 a | 116.68 a | 90.68 a | 5.86 c | 0.10 | 6.77 a |
| AM_Plantation | 3.9 abc | 72.58 a | 68.26 b | 74.71 a | 6.83 bc | 0.13 | 5.60 a |
| F-value (Species) | 4.41 * | 8.60 * | 14.73 ** | 21.56 ** | 3.82 * | 2.20 ns | 40.11 ** |
| F-value (Stand within species) | 0.30 ns | 0.24 ns | 0.76 ns | 0.63 ns | 0.21 ns | 0.10 ns | 0.77 ns |
| F-value (Species * Stand) | 0.94 ns | 0.98 ns | 0.71 ns | 0.44 ns | 0.26 ns | 1.36 ns | 0.31 ns |
| CCA Results | Axis 1 | Axis 2 | Axis 3 |
|---|---|---|---|
| Eigenvalues | 0.298 | 0.142 | 0.084 |
| Variance in species data % of variance explained | 41.40 | 19.70 | 11.70 |
| Cumulative % explained | 41.40 | 61.10 | 72.80 |
| Pearson correlation (species–environment) | 0.984 | 0.998 | 0.944 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staporn, D.; Marod, D.; Wongprom, J.; Diloksumpun, S. Drivers of Native Species Regeneration in the Process of Restoring a Dry Evergreen Forest from Exotic Tree Plantations in Northeastern Thailand. Forests 2022, 13, 1321. https://doi.org/10.3390/f13081321
Staporn D, Marod D, Wongprom J, Diloksumpun S. Drivers of Native Species Regeneration in the Process of Restoring a Dry Evergreen Forest from Exotic Tree Plantations in Northeastern Thailand. Forests. 2022; 13(8):1321. https://doi.org/10.3390/f13081321
Chicago/Turabian StyleStaporn, Duriya, Dokrak Marod, Jetsada Wongprom, and Sapit Diloksumpun. 2022. "Drivers of Native Species Regeneration in the Process of Restoring a Dry Evergreen Forest from Exotic Tree Plantations in Northeastern Thailand" Forests 13, no. 8: 1321. https://doi.org/10.3390/f13081321
APA StyleStaporn, D., Marod, D., Wongprom, J., & Diloksumpun, S. (2022). Drivers of Native Species Regeneration in the Process of Restoring a Dry Evergreen Forest from Exotic Tree Plantations in Northeastern Thailand. Forests, 13(8), 1321. https://doi.org/10.3390/f13081321