
Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe

 , ,
, ,
Abstract
:1. Introduction
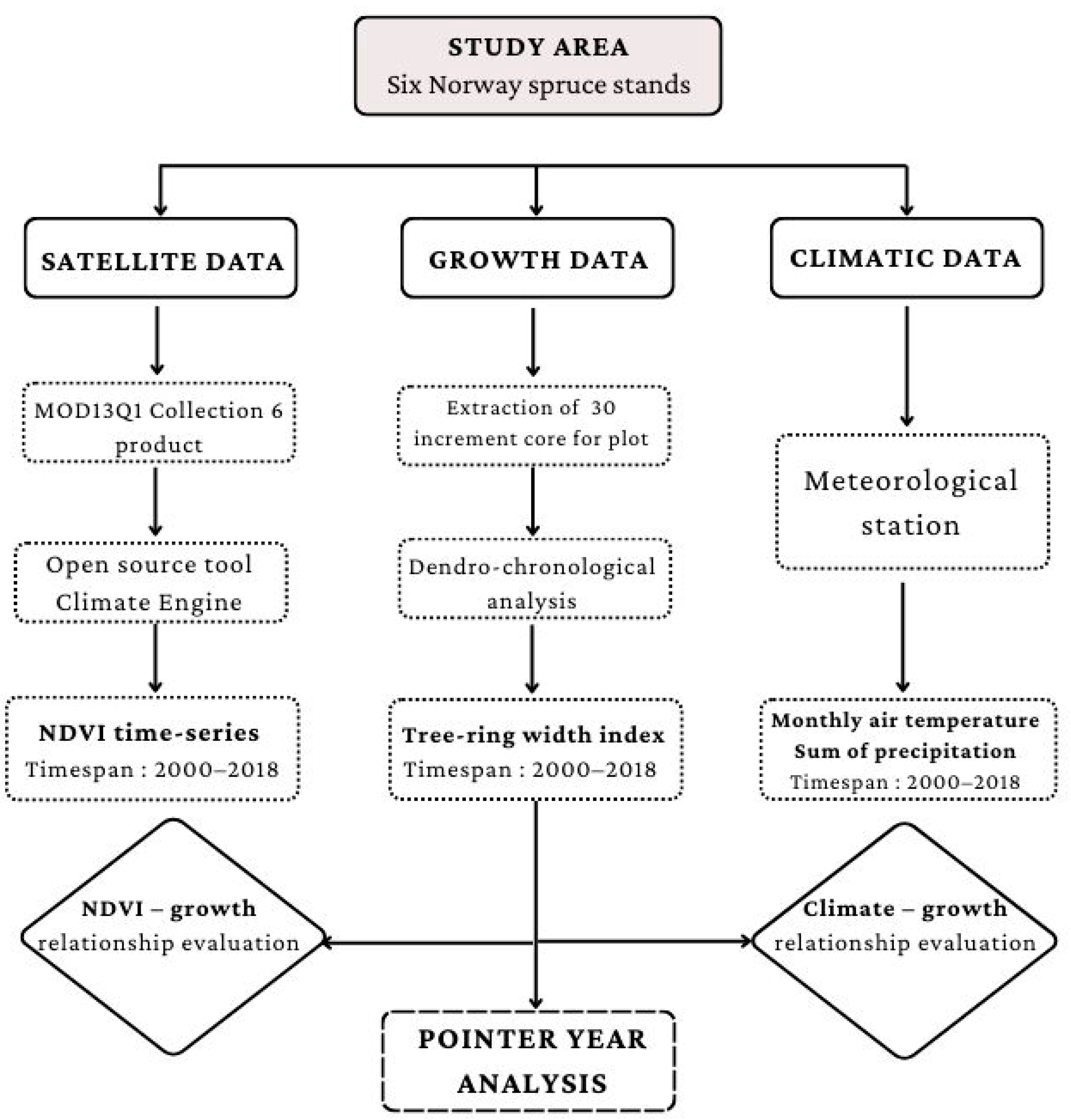
2. Material and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
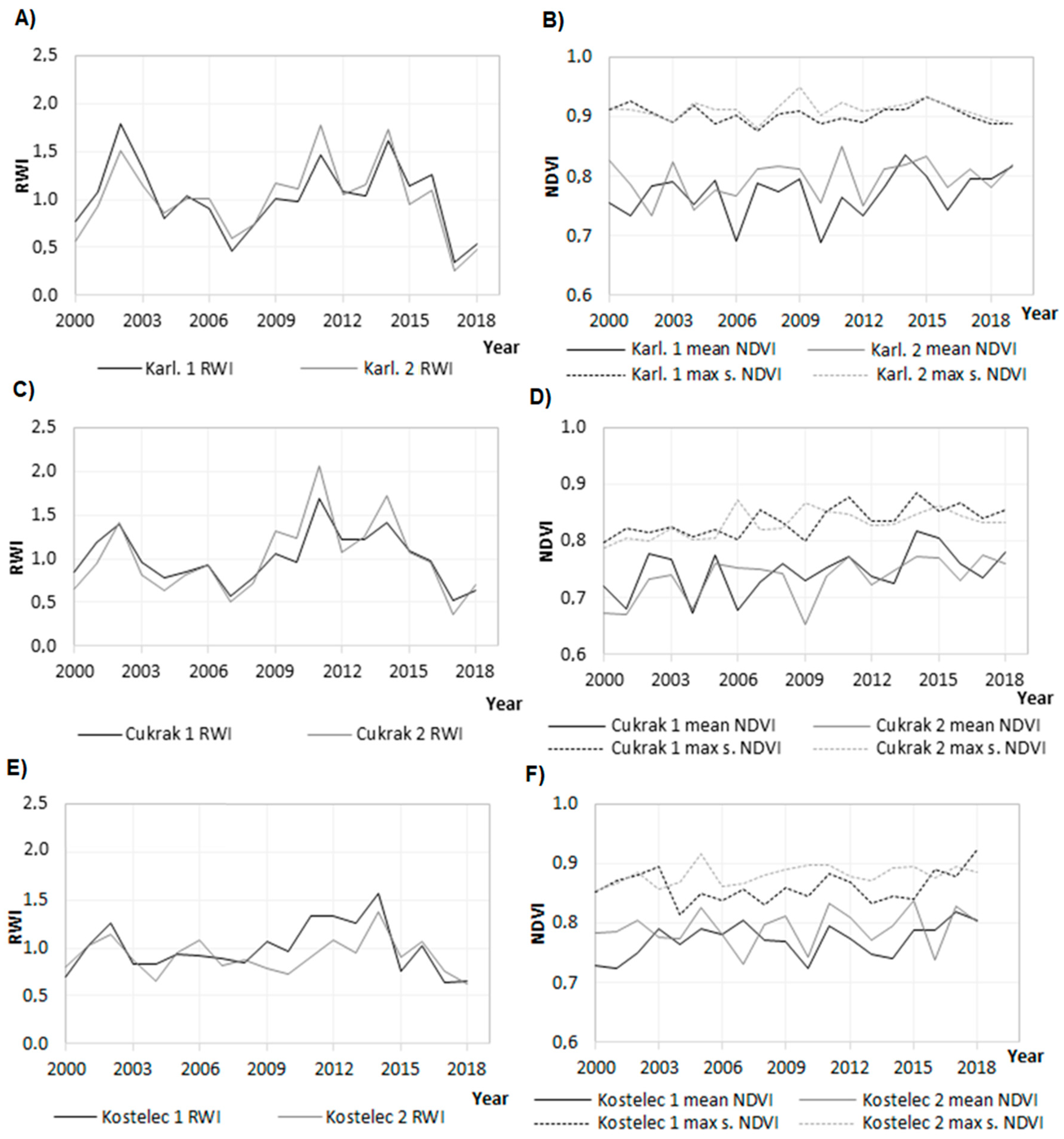
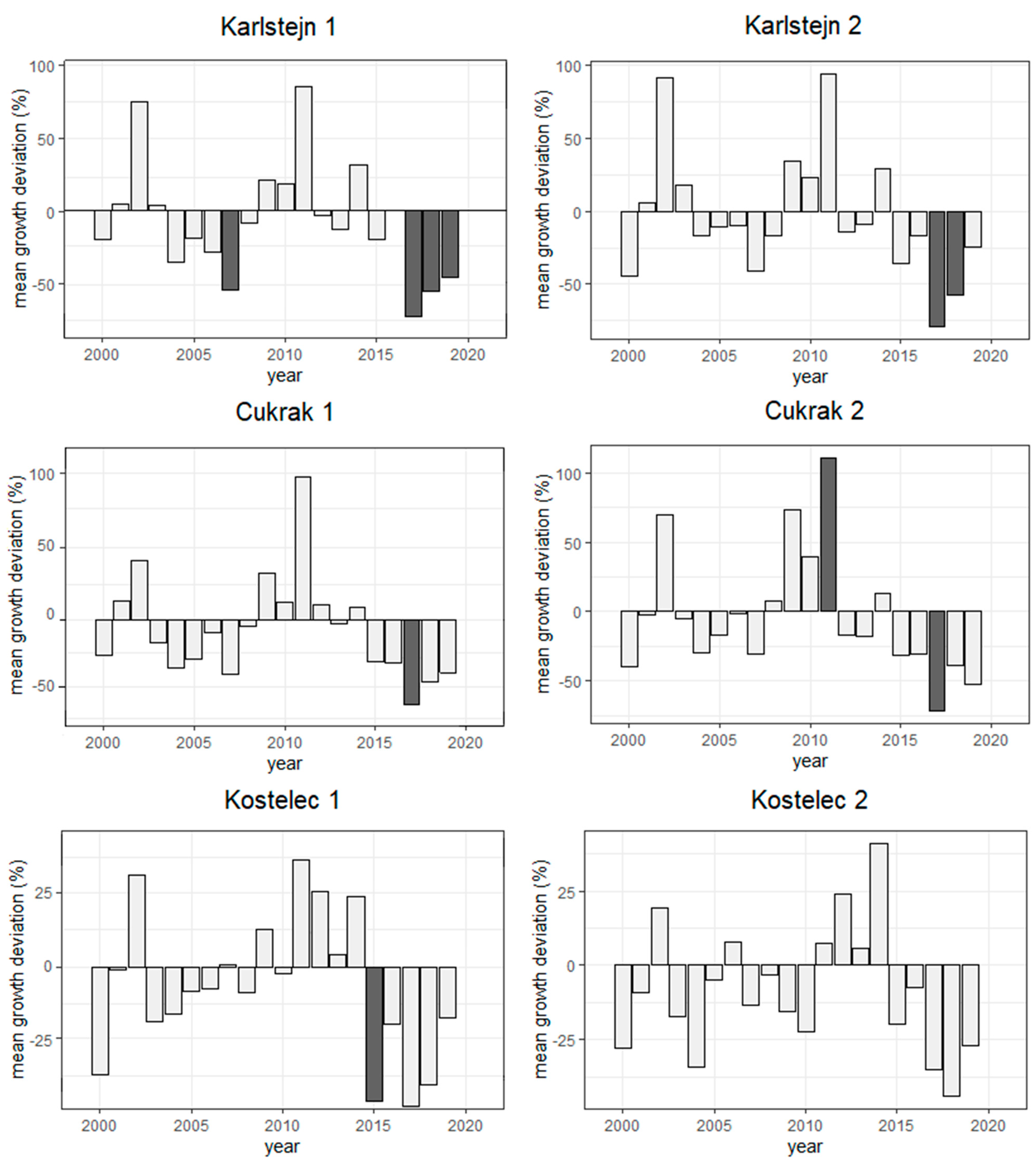
3.1. Trend of Radial Growth
3.2. Relationships between Tree-Ring Growth and NDVI
3.3. Effect of Climate on Radial Growth
4. Discussion
4.1. Trend in Radial Growth
4.2. Low Similarity between Tree-Ring Growth and NDVI
4.3. Radial Growth and Climatic Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castellaneta, M.; Rita, A.; Camarero, J.J.; Colangelo, M.; Ripullone, F. Declines in canopy greenness and tree growth are caused by combined climate extremes during drought-induced dieback. Sci. Total Environ. 2022, 813, 152666. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Vescovo, L.; Gianelle, D. Using the MIR bands in vegetation indices for the estimation of grassland biophysical parameters from satellite remote sensing in the Alps region of Trentino (Italy). Adv. Space Res. 2008, 41, 1764–1772. [Google Scholar] [CrossRef]
- Pal, S.C.; Chakrabortty, R.; Malik, S.; Das, B. Application of forest canopy density model for forest cover mapping using LISS-IV satellite data: A case study of Sali watershed, West Bengal. Model. Earth Syst. Environ. 2018, 4, 853–865. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, A.; Kumar, A.; Singh, G. Feature Extraction using Normalized Difference Vegetation Index (NDVI): A Case Study of Jabalpur City. Procedia Technol. 2012, 6, 612–621. [Google Scholar] [CrossRef]
- Goward, S.N.; Tucker, C.J.; Dye, D.G. North American vegetation patterns observed with the NOAA-7 advanced very high resolution radiometer. Vegetatio 1985, 64, 3–14. [Google Scholar] [CrossRef]
- Achard, F.; Estreguil, C. Forest classification of Southeast Asia using NOAA AVHRR data. Remote Sens. Environ. 1995, 54, 198–208. [Google Scholar] [CrossRef]
- Wang, S.; Li, R.; Wu, Y.; Zhao, S. Effects of multi-temporal scale drought on vegetation dynamics in Inner Mongolia from 1982 to 2015, China. Ecol. Indic. 2022, 136, 108666. [Google Scholar] [CrossRef]
- Morante-Carballo, F.; Bravo-Montero, L.; Carrión-Mero, P.; Velastegui-Montoya, A.; Berrezueta, E. Forest Fire Assessment Using Remote Sensing to Support the Development of an Action Plan Proposal in Ecuador. Remote Sens. 2022, 14, 1783. [Google Scholar] [CrossRef]
- Verma, S.; Sharma, A.; Yadava, P.K.; Gupta, P.; Singh, J.; Payra, S. Rapid flash flood calamity in Chamoli, Uttarakhand region during Feb 2021: An analysis based on satellite data. Nat. Hazards 2022, 112, 1379–1393. [Google Scholar] [CrossRef]
- Bojórquez, A.; Martínez-Yrízar, A.; Álvarez-Yépiz, J.C. A landscape assessment of frost damage in the northmost Neotropical dry forest. Agric. For. Meteorol. 2021, 308, 108562. [Google Scholar] [CrossRef]
- Myneni, R.; Hall, F.; Sellers, P.; Marshak, A. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remote Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jedrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen successes in animal ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- Hasenauer, H.; Petritsch, R.; Zhao, M.; Boisvenue, C.; Running, S.W. Reconciling satellite with ground data to estimate forest productivity at national scales. For. Ecol. Manag. 2012, 276, 196–208. [Google Scholar] [CrossRef]
- Brehaut, L.; Danby, R.K. Inconsistent relationships between annual tree ring-widths and satellite-measured NDVI in a mountainous subarctic environment. Ecol. Indic. 2018, 91, 698–711. [Google Scholar] [CrossRef]
- Karnieli, A.; Agam, N.; Pinker, R.T.; Anderson, M.; Imhoff, M.L.; Gutman, G.G.; Panov, N.; Goldberg, A. Use of NDVI and Land Surface Temperature for Drought Assessment: Merits and Limitations. J. Clim. 2010, 23, 618–633. [Google Scholar] [CrossRef]
- Kaufmann, R.K.; D’Arrigo, R.D.; Paletta, L.F.; Tian, H.Q.; Jolly, W.M.; Myneni, R.B. Identifying Climatic Controls on Ring Width: The Timing of Correlations between Tree Rings and NDVI. Earth Interact. 2008, 12, 1–14. [Google Scholar] [CrossRef]
- Nanzad, L.; Zhang, J.; Tuvdendorj, B.; Nabil, M.; Zhang, S.; Bai, Y. NDVI anomaly for drought monitoring and its correlation with climate factors over Mongolia from 2000 to 2016. J. Arid Environ. 2019, 164, 69–77. [Google Scholar] [CrossRef]
- Wang, Z.; Lyu, L.; Liu, W.; Liang, H.; Huang, J.; Zhang, Q.-B. Topographic patterns of forest decline as detected from tree rings and NDVI. Catena 2021, 198, 105011. [Google Scholar] [CrossRef]
- Bumann, E.; Awada, T.; Wardlow, B.; Hayes, M.; Okalebo, J.; Helzer, C.; Mazis, A.; Hiller, J.; Cherubini, P. Assessing responses of Betula papyrifera to climate variability in a remnant population along the Niobrara River Valley in Nebraska, U.S.A., through dendroecological and remote-sensing techniques. Can. J. For. Res. 2019, 49, 423–433. [Google Scholar] [CrossRef]
- Decuyper, M.; Chávez, R.O.; Čufar, K.; Estay, S.A.; Clevers, J.G.; Prislan, P.; Gričar, J.; Črepinšek, Z.; Merela, M.; de Luis, M.; et al. Spatio-temporal assessment of beech growth in relation to climate extremes in Slovenia—An integrated approach using remote sensing and tree-ring data. Agric. For. Meteorol. 2020, 287, 107925. [Google Scholar] [CrossRef]
- Wen, Y.; Jiang, Y.; Jiao, L.; Hou, C.; Xu, H. Inconsistent relationships between tree ring width and normalized difference vegetation index in montane evergreen coniferous forests in arid regions. Trees 2021, 36, 379–391. [Google Scholar] [CrossRef]
- Toth, D.; Maitah, M.; Maitah, K.; Jarolínová, V. The Impacts of Calamity Logging on the Development of Spruce Wood Prices in Czech Forestry. Forests 2020, 11, 283. [Google Scholar] [CrossRef]
- Knoke, T.; Gosling, E.; Thom, D.; Chreptun, C.; Rammig, A.; Seidl, R. Economic losses from natural disturbances in Norway spruce forests—A quantification using Monte-Carlo simulations. Ecol. Econ. 2021, 185, 107046. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Mielikäinen, K. Climatic signal in annual growth variation in damaged and healthy stands of Norway spruce [Picea abies (L.) Karst.] in Southern Finland. Trees 2001, 15, 177–185. [Google Scholar] [CrossRef]
- Vitas, A. Tree Rings of Norway Spruce (Picea Abies (L.) Karsten) in Lithuania as Drought Indicators: Dendroecological Approach. Pol. J. Ecol. 2004, 52, 201–210. [Google Scholar]
- Aakala, T.; Kuuluvainen, T. Summer droughts depress radial growth of Picea abies in pristine taiga of the Arkhangelsk province, northwestern Russia. Dendrochronologia 2011, 29, 67–75. [Google Scholar] [CrossRef]
- Rybníček, M.; Kolář, T.; Čermák, P.; Tomáš, Ž.I.D.; Hadaš, P. Dendrochronological Analysis and Habitual Stress Diagnostic Assessment of Norway Spruce (Picea abies) Stands in the Drahany Highlands. Wood Res. 2012, 57, 189–206. [Google Scholar]
- Putalová, T.; Vacek, Z.; Vacek, S.; Štefančík, I.; Bulušek, D.; Král, J. Tree-ring widths as an indicator of air pollution stress and climate conditions in different Norway spruce forest stands in the Krkonoše Mts. Cent. Eur. For. J. 2019, 65, 21–33. [Google Scholar] [CrossRef]
- Šimůnek, V.; Vacek, Z.; Vacek, S. Solar Cycles in Salvage Logging: National Data from the Czech Republic Confirm Significant Correlation. Forests 2020, 11, 973. [Google Scholar] [CrossRef]
- Gomez, D.; Ritger, H.; Pearce, C.; Eickwort, J.; Hulcr, J. Ability of Remote Sensing Systems to Detect Bark Beetle Spots in the Southeastern US. Forests 2020, 11, 1167. [Google Scholar] [CrossRef]
- Ge, Z.-M.; Kellomäki, S.; Peltola, H.; Zhou, X.; Wang, K.-Y.; Väisänen, H. Impacts of changing climate on the productivity of Norway spruce dominant stands with a mixture of Scots pine and birch in relation to water availability in southern and northern Finland. Tree Physiol. 2011, 31, 323–338. [Google Scholar] [CrossRef]
- Bergh, J.; Linder, S.; Bergström, J. Potential production of Norway spruce in Sweden. For. Ecol. Manag. 2005, 204, 1–10. [Google Scholar] [CrossRef]
- Koprowski, M.; Zielski, A. Dendrochronology of Norway spruce (Picea abies (L.) Karst.) from two range centres in lowland Poland. Trees 2006, 20, 383–390. [Google Scholar] [CrossRef]
- Savva, Y.; Oleksyn, J.; Reich, P.B.; Tjoelker, M.G.; Vaganov, E.A.; Modrzynski, J. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees 2006, 20, 735–746. [Google Scholar] [CrossRef]
- Rybníček, M.; Cermak, P.; Kolář, T.; Přemyslovská, E.; Žid, T. Influence of temperatures and precipitation on radial increment of Orlické hory Mts. spruce stands at altitudes over 800 m a.s.l. J. For. Sci. 2009, 55, 257–263. [Google Scholar] [CrossRef]
- Gryc, V.; Vavrčík, H.; Vichrová, G. Monitoring of Xylem Formation in Norway Spruce in the Czech Republic 2009. Wood Res. 2011, 56, 467–478. [Google Scholar]
- Vacek, Z.; Vacek, S.; Prokůpková, A.; Bulušek, D.; Podrázský, V.; Hůnová, I.; Putalová, T.; Král, J. Long-term effect of climate and air pollution on health status and growth of Picea abies (L.) Karst. peaty forests in the Black Triangle region. Dendrobiology 2020, 83, 1–19. [Google Scholar] [CrossRef]
- Němeček, J.; Macků, J.; Vokoun, J.; Vavříček, D.; Novák, P. Taxonomický Klasifikační Systém Půd České Republiky; ČZU: Prague, Czech Republic, 2001. [Google Scholar]
- Viewegh, J.; Kusbach, A.; Mikeska, M. Czech Forest Ecosystem Classification. J. For. Sci. 2003, 49, 74–82. [Google Scholar] [CrossRef]
- Köppen, W. Das Geographische System Der Klimate, Handbuch Der Klimatologie; Gebrüder Borntraeger: Berlin, Germany, 1936. [Google Scholar]
- Pekařová, K. Dyinamics of Threatened Species of the Family Ranunculaceae on the Steppe Localities of Doutnáč Hill in the National Natural Reserve. Sci. Agric. Bohem. 2007, 3, 24–33. [Google Scholar]
- Bílek, L.; Remeš, J.; Zahradník, D. Natural Regeneration of Senescent Even-Aged Beech (Fagus Sylvatica L) Stands under the Conditions of Central Bohemia. J. For. Sci. Sci. 2009, 55, 145–155. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Ulbrichová, I.; Simon, J.; Bulušek, D. Impact of applied silvicultural systems on spatial pattern of hornbeam-oak forests. Cent. Eur. For. J. 2018, 64, 33–45. [Google Scholar] [CrossRef] [Green Version]
- CHMU Czech Hydrometeorological Institute. Available online: http://portal.chmi.cz/historicka-data/pocasi/uzemni-srazky (accessed on 25 June 2020).
- Remes, J.; Podrázský, V. Fertilization of spruce monocultures in the territory of Training Forest Enterprise in Kostelec nad Černými lesy. J. For. Sci. 2006, 52, S73–S78. [Google Scholar] [CrossRef]
- Huntington, J.L.; Hegewisch, K.C.; Daudert, B.; Morton, C.G.; Abatzoglou, J.T.; McEvoy, D.J.; Erickson, T. Climate Engine: Cloud Computing and Visualization of Climate and Remote Sensing Data for Advanced Natural Resource Monitoring and Process Understanding. Bull. Am. Meteorol. Soc. 2017, 98, 2397–2410. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Schweingrub, F.H.; Eckstein, D.; Serre-Bachet, F.; Braker, O.U. Identification, Presentation and Interpretation of Event Years and Pointer Years in Dendrochronology. Dendrochronologia 1990, 8, 9–38. [Google Scholar]
- Kraft, G. Beiträge Zur Lehre von Den Durchforstungen, Schlagstellungen und Lichtungshieben; Klindworth: Hannover, Germany, 1884. [Google Scholar]
- Remes, J.; Bílek, L.; Novák, J.; Vacek, Z.; Vacek, S.; Putalová, T.; Koubek, L. Diameter increment of beech in relation to social position of trees, climate characteristics and thinning intensity. J. For. Sci. 2015, 61, 456–464. [Google Scholar] [CrossRef]
- Larsson, L. CooRecorder and Cdendro Programs of the Coorecorder/Cdendropackage Version 7.6. 2013. Available online: http://www.cybis.se/forfun/dendro (accessed on 20 June 2019).
- Rinntech. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and Related Applications; Rinntech: Heidelberg, Germany, 2003; Available online: http://www.rimatech.com (accessed on 20 June 2019).
- Zang, C.; Buras, A.; Cecile, J.; Mudelsee, M.; Schulz, M.; Pucha-cofrep, D. Package ‘dplR’ R, Dendrochronology Program Library in R Version 2018. Available online: https://r-forge.r-project.org/projects/dplr/ (accessed on 25 June 2020).
- Shumilov, O.; Timonen, M.; Kanatjev, A.G. Palaeovolcanos, Solar Activity and Pine Tree-Rings from the Kola Peninsula (Northwestern Russia) over the Last 560 Years Palaeovolcanos. Int. J. Environ. Res. 2011, 5, 855–864. [Google Scholar]
- Bunn, A.; Mikko, K. Chronology Building in DplR; R Foundation for Statistical Computing: Vienna, Austria, 2018; pp. 1–13. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- StatSoft. Statistica Electronic Manual; Statsoft: Tulsa, OK, USA, 2013. [Google Scholar]
- Lim, C.Y.; Stein, M. Properties of spatial cross-periodograms using fixed-domain asymptotics. J. Multivar. Anal. 2008, 99, 1962–1984. [Google Scholar] [CrossRef]
- Hájek, V.; Vacek, S.; Vacek, Z.; Cukor, J.; Šimůnek, V.; Šimková, M.; Prokůpková, A.; Králíček, I.; Bulušek, D. Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic). Forests 2021, 12, 1127. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Boratyński, A.; Bugala, W. Biology and Ecology of Norway Spruce; Springer Science & Business Media: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea Abies in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Bugmann, H.; Brang, P. Toward Quantitative Scenarios of Climate Change Impacts in Switzerland; Swiss Federal Institute for Forest: Bern, Switzerland, 2014. [Google Scholar]
- Rita, A.; Camarero, J.J.; Nolè, A.; Borghetti, M.; Brunetti, M.; Pergola, N.; Serio, C.; Vicente-Serrano, S.M.; Tramutoli, V.; Ripullone, F. The impact of drought spells on forests depends on site conditions: The case of 2017 summer heat wave in southern Europe. Glob. Change Biol. 2019, 26, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Schär, C.; Vidale, P.L.; Lüthi, D.; Frei, C.; Häberli, C.; Liniger, M.A.; Appenzeller, C. The role of increasing temperature variability in European summer heatwaves. Nature 2004, 427, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- García-Herrera, R.; Díaz, J.; Trigo, R.M.; Luterbacher, J.; Fischer, E.M. A Review of the European Summer Heat Wave of 2003. Crit. Rev. Environ. Sci. Technol. 2010, 40, 267–306. [Google Scholar] [CrossRef]
- Rybníček, M.; Čermák, P.; Kolář, T.; Žid, T. Growth responses of Norway spruce (Picea abies (L.) Karst.) to the climate in the south-eastern part of the Českomoravská Upland (Czech Republic). Geochronometria 2012, 39, 149–157. [Google Scholar] [CrossRef]
- Andreu-Hayles, L.; D’Arrigo, R.; Anchukaitis, K.J.; Beck, P.S.A.; Frank, D.; Goetz, S. Varying boreal forest response to Arctic environmental change at the Firth River, Alaska. Environ. Res. Lett. 2011, 6, 045503. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Camarero, J.J.; Olano, J.M.; Martín-Hernández, N.; Peña-Gallardo, M.; Tomás-Burguera, M.; Gazol, A.; Azorin-Molina, C.; Bhuyan, U.; El Kenawy, A. Diverse relationships between forest growth and the Normalized Difference Vegetation Index at a global scale. Remote Sens. Environ. 2016, 187, 14–29. [Google Scholar] [CrossRef]
- D’Arrigo, R.D.; Malmstrom, C.M.; Jacoby, G.C.; Los, S.O.; Bunker, D.E. Correlation between maximum latewood density of annual tree rings and NDVI based estimates of forest productivity. Int. J. Remote Sens. 2000, 21, 2329–2336. [Google Scholar] [CrossRef]
- Lawrence, G.B.; Lapenis, A.G.; Berggren, D.; Aparin, B.F.; Smith, K.T.; Shortle, W.C.; Bailey, S.W.; Varlyguin, D.L.; Babikov, B. Climate Dependency of Tree Growth Suppressed by Acid Deposition Effects on Soils in Northwest Russia. Environ. Sci. Technol. 2005, 39, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Beck, P.S.; Andreu-Hayles, L.; D’Arrigo, R.; Anchukaitis, K.J.; Tucker, C.J.; Pinzón, J.E.; Goetz, S.J. A large-scale coherent signal of canopy status in maximum latewood density of tree rings at arctic treeline in North America. Glob. Planet. Chang. 2013, 100, 109–118. [Google Scholar] [CrossRef]
- Berner, L.T.; Beck, P.S.A.; Bunn, A.G.; Goetz, S.J. Plant response to climate change along the forest-tundra ecotone in northeastern Siberia. Glob. Chang. Biol. 2013, 19, 3449–3462. [Google Scholar] [CrossRef] [PubMed]
- Soudani, K.; le Maire, G.; Dufrêne, E.; François, C.; Delpierre, N.; Ulrich, E.; Cecchini, S. Evaluation of the onset of green-up in temperate deciduous broadleaf forests derived from Moderate Resolution Imaging Spectroradiometer (MODIS) data. Remote Sens. Environ. 2008, 112, 2643–2655. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.-Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.R.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Testa, S.; Mondino, E.C.B.; Pedroli, C. Correcting MODIS 16-day composite NDVI time-series with actual acquisition dates. Eur. J. Remote Sens. 2014, 47, 285–305. [Google Scholar] [CrossRef]
- Bhuyan, U.; Zang, C.; Vicente-Serrano, S.M.; Menzel, A. Exploring Relationships among Tree-Ring Growth, Climate Variability, and Seasonal Leaf Activity on Varying Timescales and Spatial Resolutions. Remote Sens. 2017, 9, 526. [Google Scholar] [CrossRef]
- Fritts, H.C. Growth-Rings of Trees: Their Correlation with Climate. Science 1966, 154, 973–979. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Kahle, H.-P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Králíček, I.; Vacek, Z.; Vacek, S.; Remeš, J.; Bulušek, D.; Král, J.; Štefančík, I.; Putalová, T. Dynamics and structure of mountain autochthonous spruce-beech forests: Impact of hilltop phenomenon, air pollutants and climate. Dendrobiology 2017, 77, 119–137. [Google Scholar] [CrossRef]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- D’Arrigo, R.D.; Kaufmann, R.K.; Davi, N.; Jacoby, G.C.; Laskowski, C.; Myneni, R.B.; Cherubini, P. Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory, Canada. Glob. Biogeochem. Cycles 2004, 18, GB3021. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Griesbauer, H.P.; Green, D.S. Growth responses of three coexisting conifer species to climate across wide geographic and climate ranges in Yukon and British Columbia. For. Ecol. Manag. 2010, 259, 514–523. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Grabařová, S.; Martinková, M. Changes of Norway Spruce (Picea Abies [L.] Karst.) Growth Characteristics under the Impact of Drought. Ekológia 2000, 19, 81–103. [Google Scholar]
- Vacek, Z.; Řeháček, D.; Cukor, J.; Vacek, S.; Khel, T.; Sharma, R.P.; Kučera, J.; Král, J.; Papaj, V. Windbreak Efficiency in Agricultural Landscape of the Central Europe: Multiple Approaches to Wind Erosion Control. Environ. Manag. 2018, 62, 942–954. [Google Scholar] [CrossRef]
- Kolář, T.; Čermák, P.; Trnka, M.; Koňasová, E.; Sochová, I.; Rybníček, M. Dendroclimatic study of a mixed spruce-fir-beech forest in the Czech Republic. Les/Wood 2020, 69, 21–32. [Google Scholar] [CrossRef]
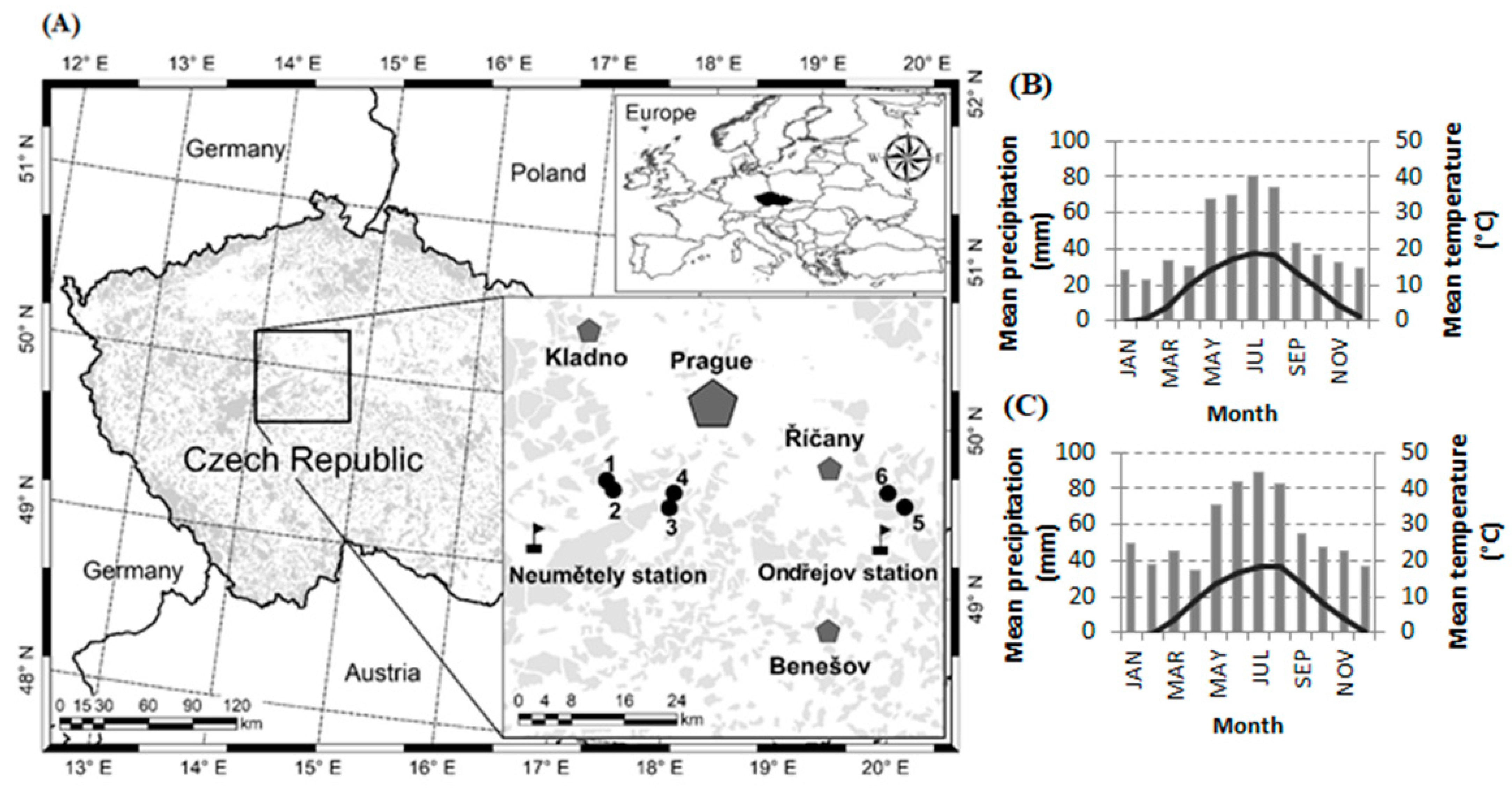
 ) (A); pentagons mean big cities; the shadowed areas are forested areas; the map was made in ArcGIS 10 software (Esri). Climogram for Karlstejn and Cukrak research plots (B); Climogram for Kostelec research plots (C).
) (A); pentagons mean big cities; the shadowed areas are forested areas; the map was made in ArcGIS 10 software (Esri). Climogram for Karlstejn and Cukrak research plots (B); Climogram for Kostelec research plots (C).
) (A); pentagons mean big cities; the shadowed areas are forested areas; the map was made in ArcGIS 10 software (Esri). Climogram for Karlstejn and Cukrak research plots (B); Climogram for Kostelec research plots (C).
) (A); pentagons mean big cities; the shadowed areas are forested areas; the map was made in ArcGIS 10 software (Esri). Climogram for Karlstejn and Cukrak research plots (B); Climogram for Kostelec research plots (C).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | GPS | Altitude | Exposure | Slope | Age of Tree Layers | Height | DBH | Forest |
|---|---|---|---|---|---|---|---|---|
| Coordinates | [m] | [%] | [years] | [m] | [cm] | Type * | ||
| Karlstejn 1 | 49°56′51.3′′ N, 14°12′05.6′′ E | 422 | N–W | 5–10 | 93 | 26 | 36 | 3B |
| Karlstejn 2 | 49°56′41.6′′ N, 14°12′19.6′′ E | 406 | W | <5 | 83 | 24 | 32 | 2H |
| Cukrak 1 | 49°56′14.2′′ N, 14°21′13.4′′ E | 402 | N–W | 20–25 | 83 | 23 | 29 | 3K |
| Cukrak 2 | 49°56′51.3′′ N, 14°21′25.1′′ E | 319 | N–W | 5-10 | 80 | 26 | 30 | 3I |
| Kostelec 1 | 49°57′55.9′′ N, 14°48′58.9′′ E | 423 | N–W | <5 | 96 | 31 | 46 | 3S |
| Kostelec 2 | 49°58′33.5′′ N, 14°47′20.9′′ E | 425 | N–E | 10-15 | 129 | 30 | 35 | 3I |
| Plot Name | No. Trees | Mean RW (mm) | Mean Min—Max RW (mm) | Age (Years) | Std. (mm) | ar1 | R-Bar | ESP | SNR |
|---|---|---|---|---|---|---|---|---|---|
| Karlstejn 1 | 27 | 1.79 | 1.29–2.58 | 93 | 0.87 | 0.55 | 0.59 | 0.97 | 32.75 |
| Karlstejn 2 | 30 | 1.61 | 0.99–2.25 | 83 | 0.94 | 0.66 | 0.53 | 0.97 | 28.90 |
| Cukrak 1 | 31 | 1.65 | 1.09–2.63 | 83 | 0.82 | 0.60 | 0.46 | 0.96 | 22.14 |
| Cukrak 2 | 27 | 1.94 | 1.15–3.20 | 80 | 1.11 | 0.57 | 0.62 | 0.98 | 40.14 |
| Kostelec 1 | 28 | 2.26 | 1.22–3.34 | 96 | 0.96 | 0.65 | 0.47 | 0.96 | 21.29 |
| Kostelec 2 | 30 | 1.45 | 1.03–2.17 | 129 | 0.69 | 0.73 | 0.39 | 0.95 | 17.68 |
| Plot Name | Mean NDVI | Mean Seasonal NDVI | Max Seasonal NDVI | Temperature | Seasonal Temperature | Precipitation | Seasonal Precipitation |
|---|---|---|---|---|---|---|---|
| Karlstejn 1 RWI | 0.36 | 0.10 | 0.29 | −0.11 | −0.28 | 0.40 | 0.44 |
| p-value | 0.13 | 0.68 | 0.23 | 0.64 | 0.25 | 0.09 | 0.06 |
| Karlstejn 2 RWI | 0.01 | 0.10 | 0.33 | −0.24 | −0.40 | 0.49 | 0.59 |
| p-value | 0.95 | 0.68 | 0.17 | 0.33 | 0.09 | 0.03 | 0.08 |
| Cukrak 1 RWI | 0.29 | 0.18 | 0.26 | −0.13 | −0.30 | 0.55 | 0.62 |
| p-value | 0.23 | 0.45 | 0.29 | 0.61 | 0.22 | 0.02 | 0.01 |
| Cukrak 2 RWI | 0.16 | 0.15 | 0.39 | −0.13 | −0.30 | 0.57 | 0.67 |
| p-value | 0.55 | 0.54 | 0.10 | 0.60 | 0.22 | 0.01 | 0.01 |
| Kostelec 1 RWI | −0.36 | −0.26 | −0.08 | −0.30 | −0.53 | 0.50 | 0.59 |
| p-value | 0.13 | 0.28 | 0.75 | 0.21 | 0.02 | 0.03 | 0.01 |
| Kostelec 2 RWI | 0.06 | 0.22 | 0.01 | 0.04 | −0.15 | 0.43 | 0.50 |
| p-value | 0.80 | 0.37 | 0.95 | 0.87 | 0.55 | 0.07 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Andrea, G.; Šimůnek, V.; Castellaneta, M.; Vacek, Z.; Vacek, S.; Pericolo, O.; Zito, R.G.; Ripullone, F. Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe. Forests 2022, 13, 1417. https://doi.org/10.3390/f13091417
D’Andrea G, Šimůnek V, Castellaneta M, Vacek Z, Vacek S, Pericolo O, Zito RG, Ripullone F. Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe. Forests. 2022; 13(9):1417. https://doi.org/10.3390/f13091417
Chicago/Turabian StyleD’Andrea, Giuseppe, Václav Šimůnek, Maria Castellaneta, Zdeněk Vacek, Stanislav Vacek, Osvaldo Pericolo, Rosa Giada Zito, and Francesco Ripullone. 2022. "Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe" Forests 13, no. 9: 1417. https://doi.org/10.3390/f13091417
APA StyleD’Andrea, G., Šimůnek, V., Castellaneta, M., Vacek, Z., Vacek, S., Pericolo, O., Zito, R. G., & Ripullone, F. (2022). Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe. Forests, 13(9), 1417. https://doi.org/10.3390/f13091417