
Effect of Provenance and Environmental Factors on Tree Growth and Tree Water Status of Norway Spruce

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Plant Material
2.2. Environmental Data
2.3. Band Dendrometer Records (BDR)
2.4. Data Analysis and Statistical Evaluation
- Random forest (RF) is a decision tree nonparametric algorithm for classification or regression [47]. The algorithm selects a random number of samples from training datasets, what is called a bootstrap aggregation [48]. Afterwards, the randomly chosen samples are used to develop a decision tree based on the most important variables. For each prediction, multiple decisions trees are constructed, and the average value is outputted.
- Gradient boosting machine (GBM) is also based on decision trees. The main difference to RF is that the random forest uses averaging, while the gradient boosting provides additive (ensamble) modelling [49]. Moreover, the random forest combines results at the end of the process, while the gradient boosting combines the results along the way.
- Support vector machines (SVM) are used both in classification and regression. In support vector regression, the space that is required to fit the data is referred to as a hyperplane [50]. A hyperplane is a subspace with the number of dimensions equal to that of the original space minus one. The best fit is the hyperplane that contains the maximum number of points within threshold values. This separates SVM from other regression models which minimise errors between real and predicted values. The hyperplane is determined using a kernel, which is a set of mathematical functions that takes data as input and transforms it into the required form.
- Neural networks (NN), also known as artificial neural networks (ANNs), are composed at least of three layers: input, output layers and one or more hidden layers. Each neuron has a specific weight and a threshold value and is connected to other neurons. If the output of a neuron exceeds the threshold value, the neuron is activated and sends the signal (data) to the next layer of the network [51]. Deep neural network (DNN) refers to the situation when more than one hidden layer is applied. The deeper the DNN, the more complex patterns the network can learn [52].
3. Results
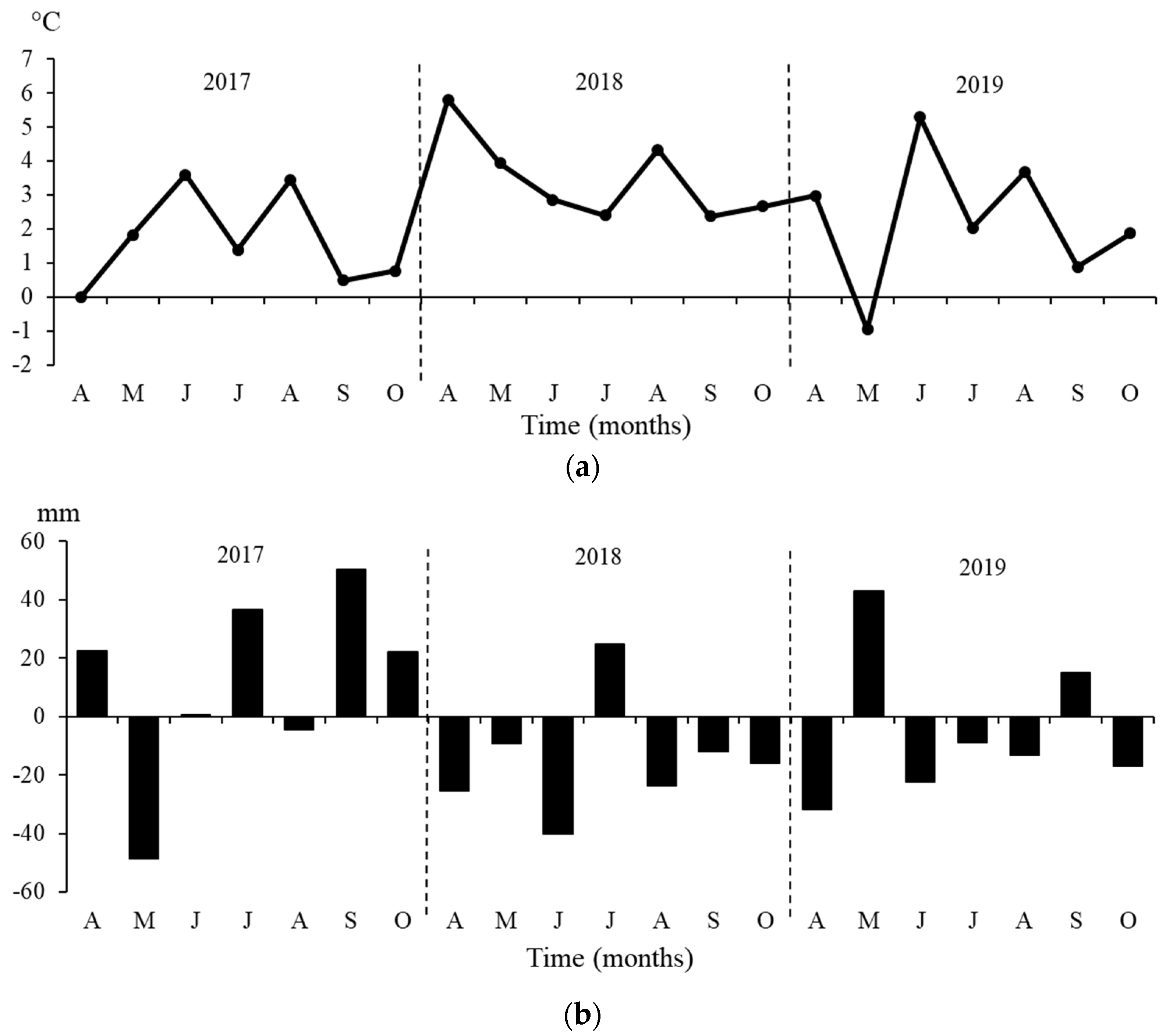
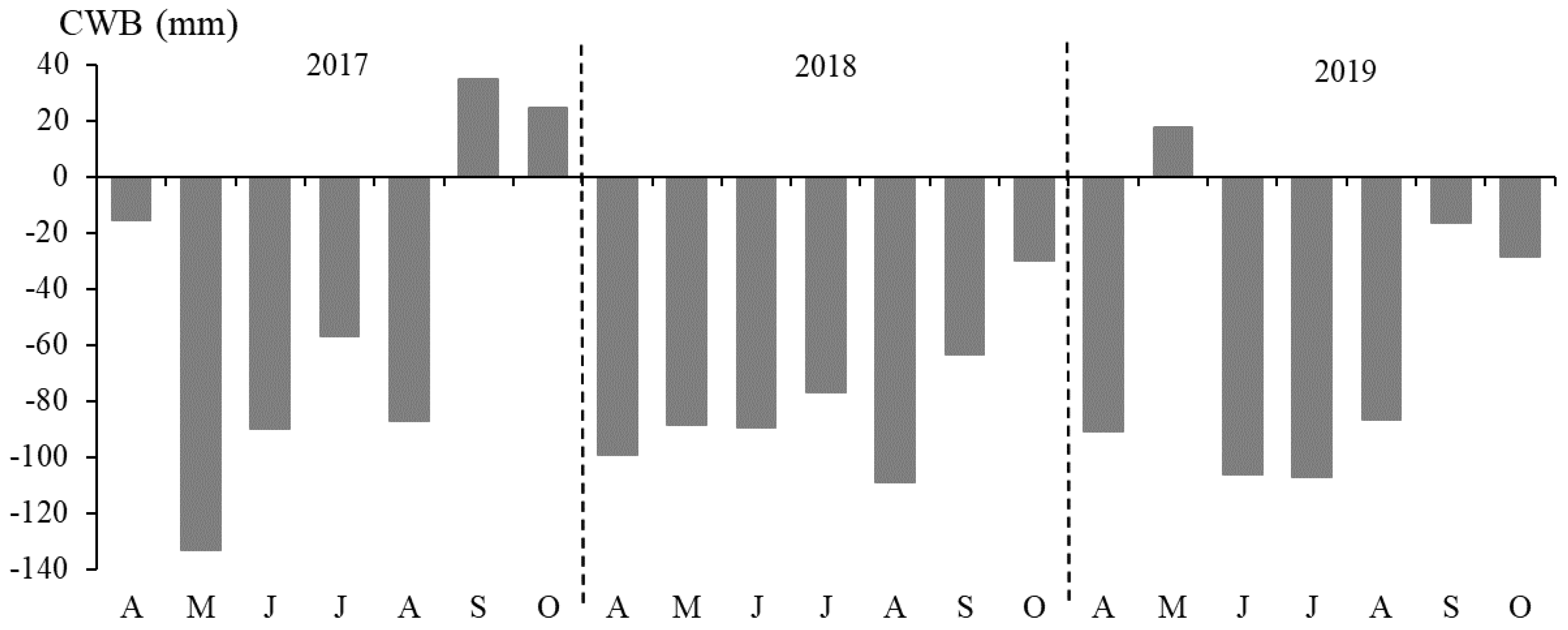
3.1. Environmental Variables during the Growing Seasons of the Years 2017–2019
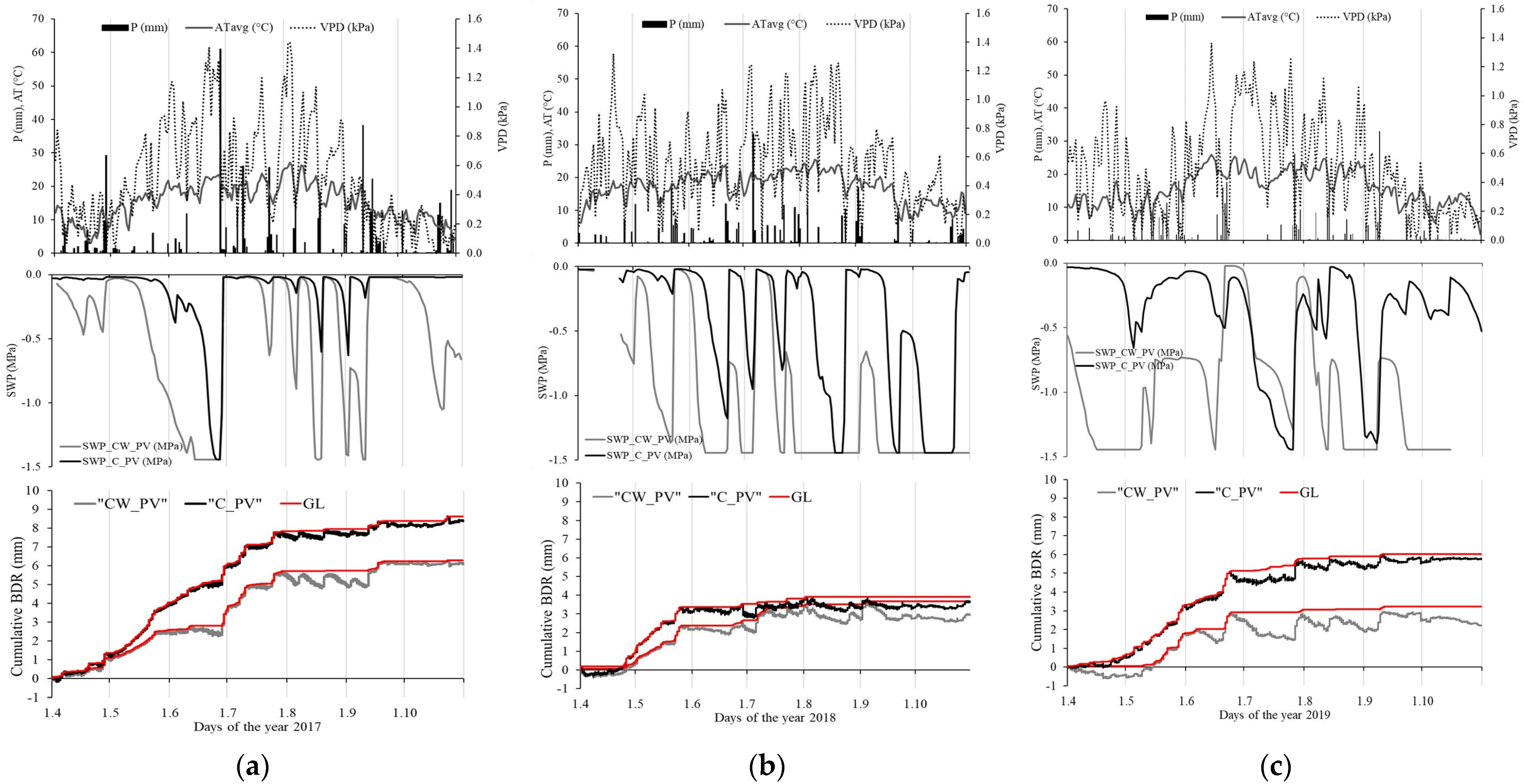
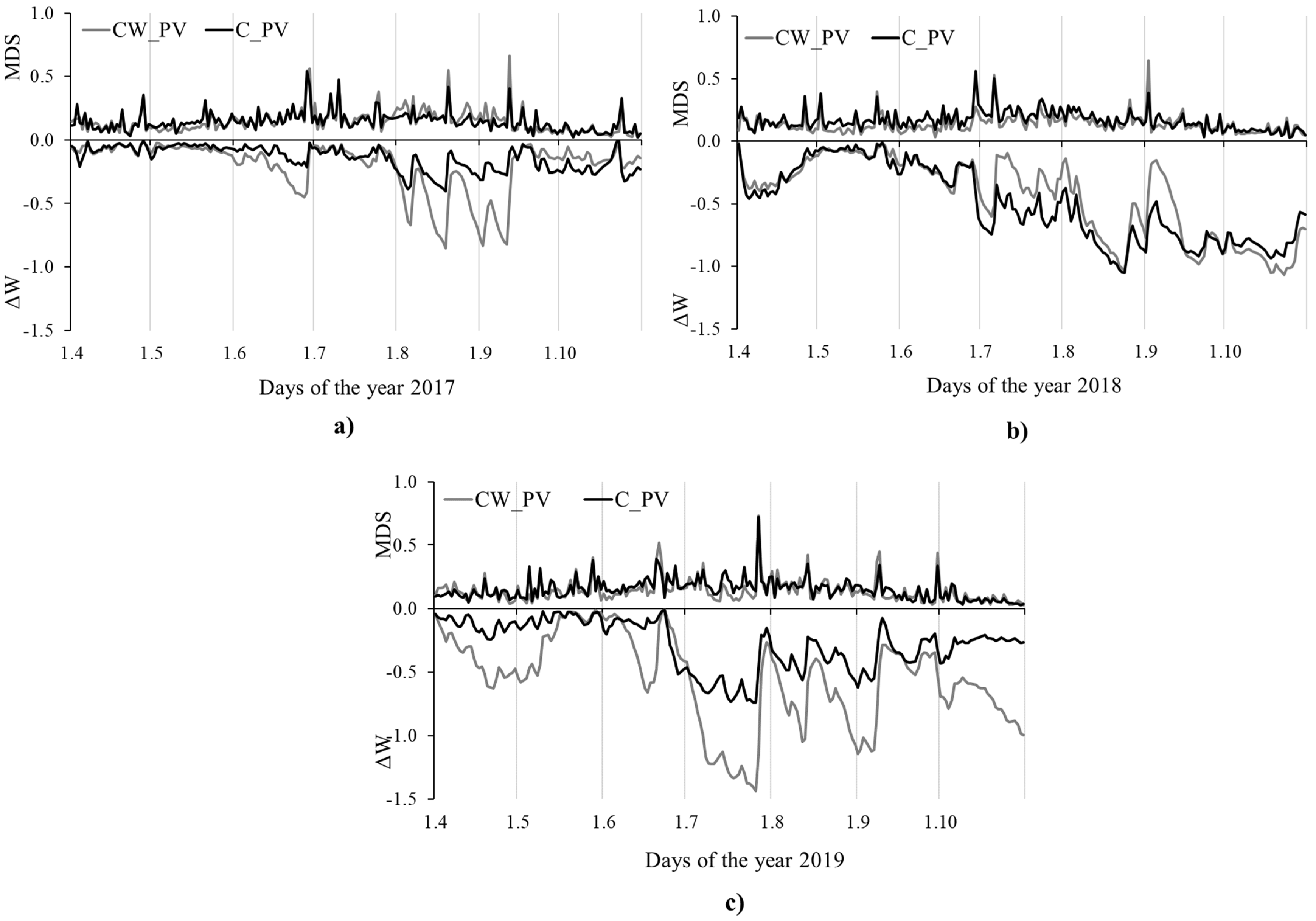
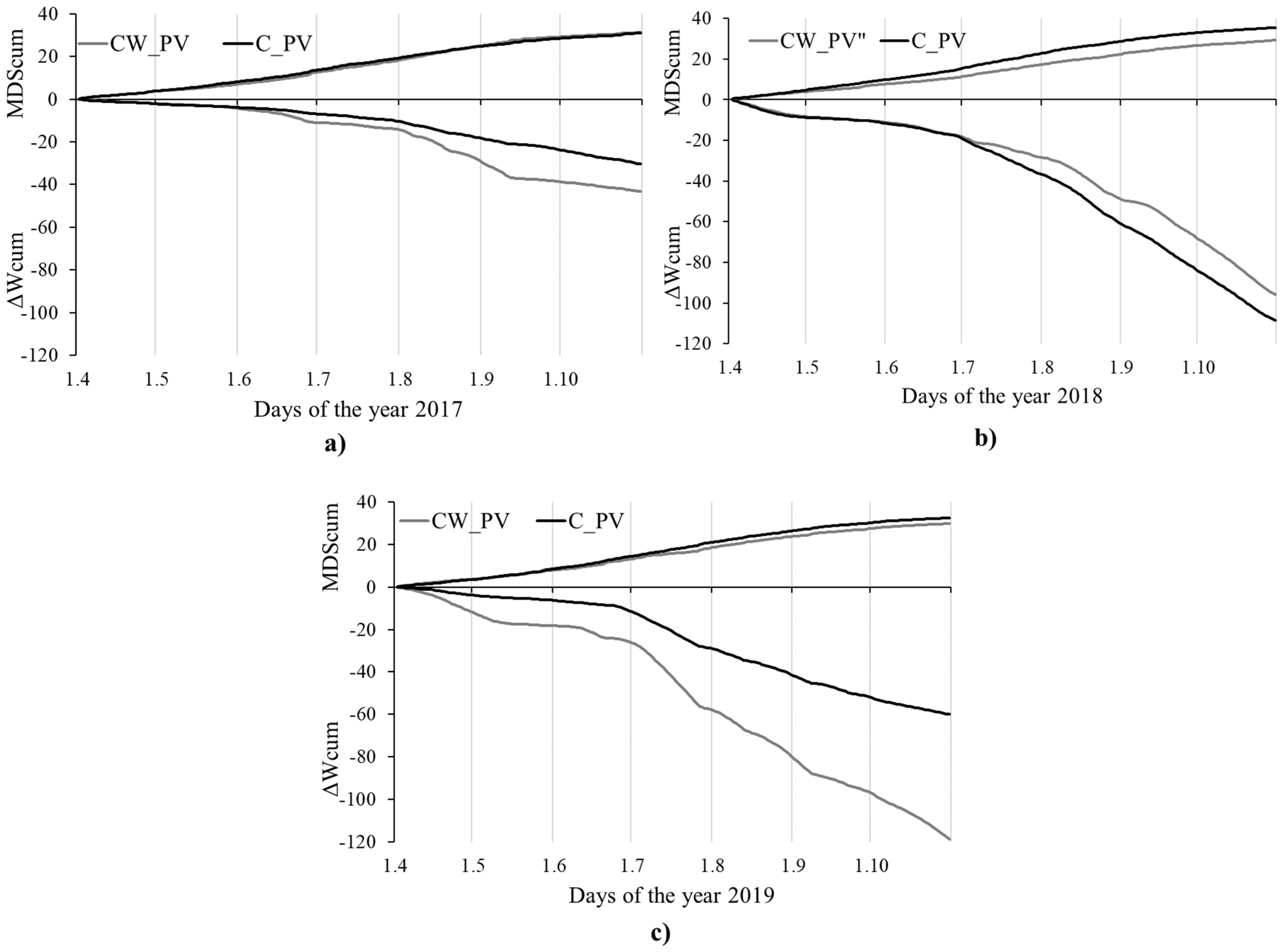
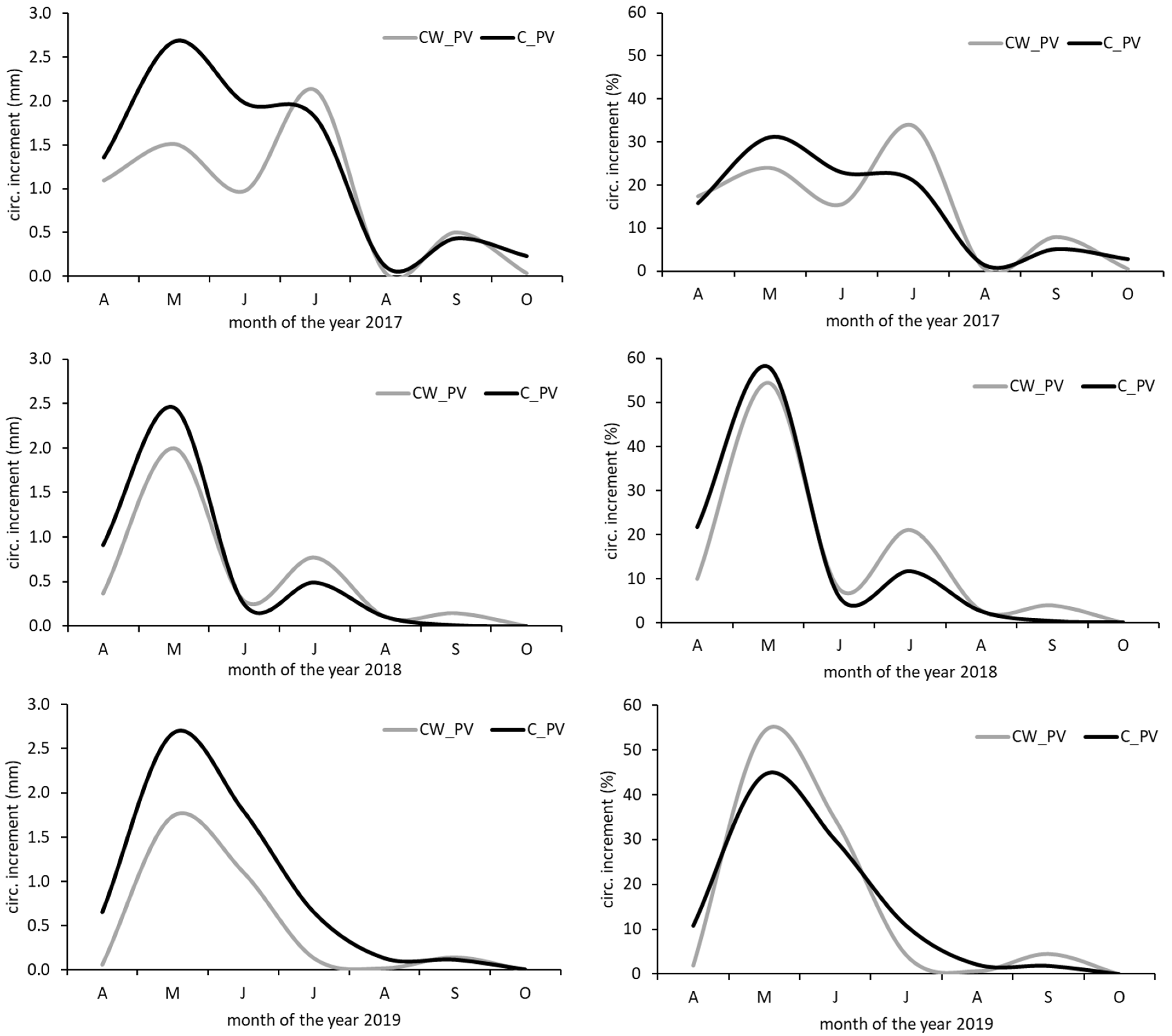
3.2. Stem Radius Changes and Stem Water Status
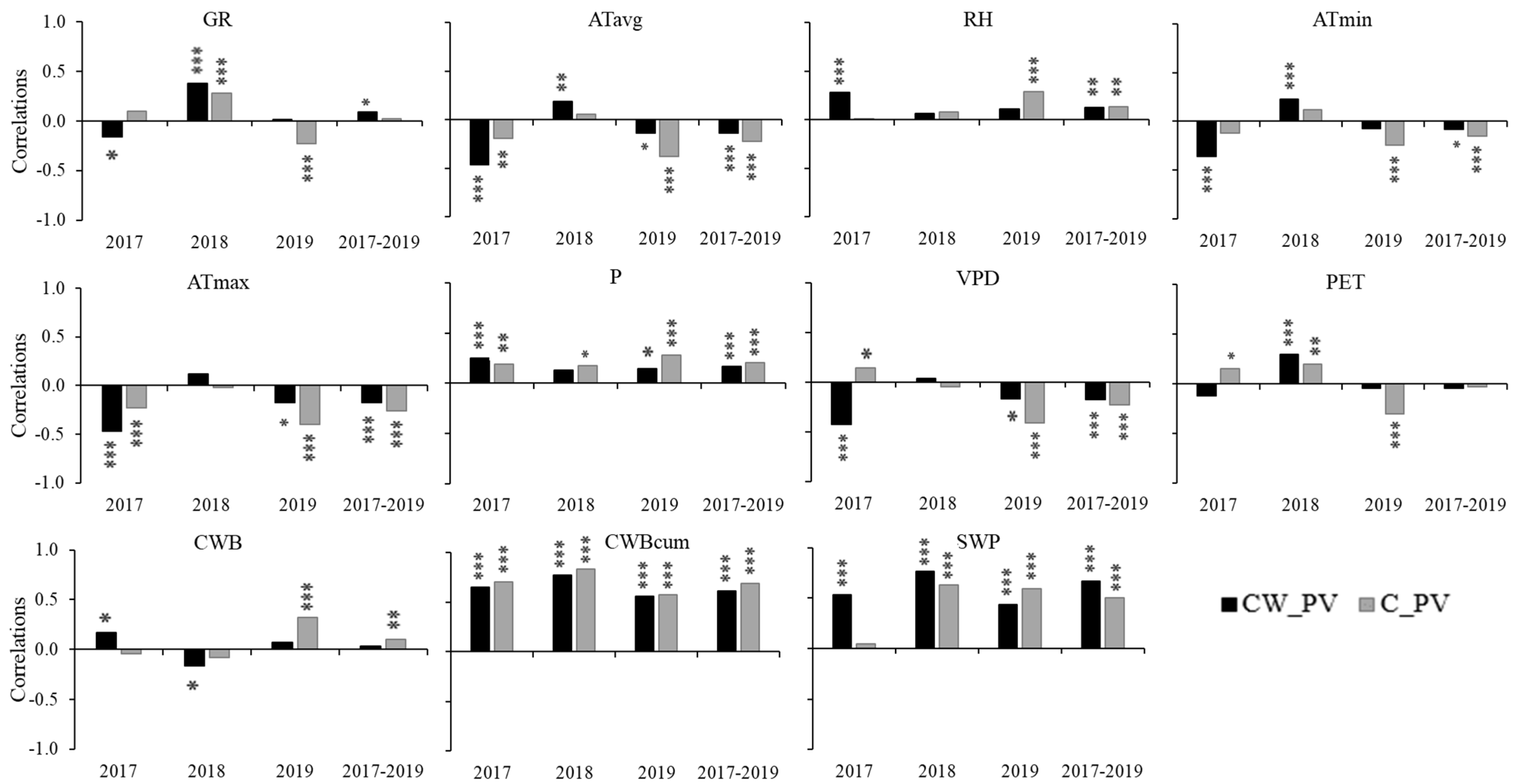
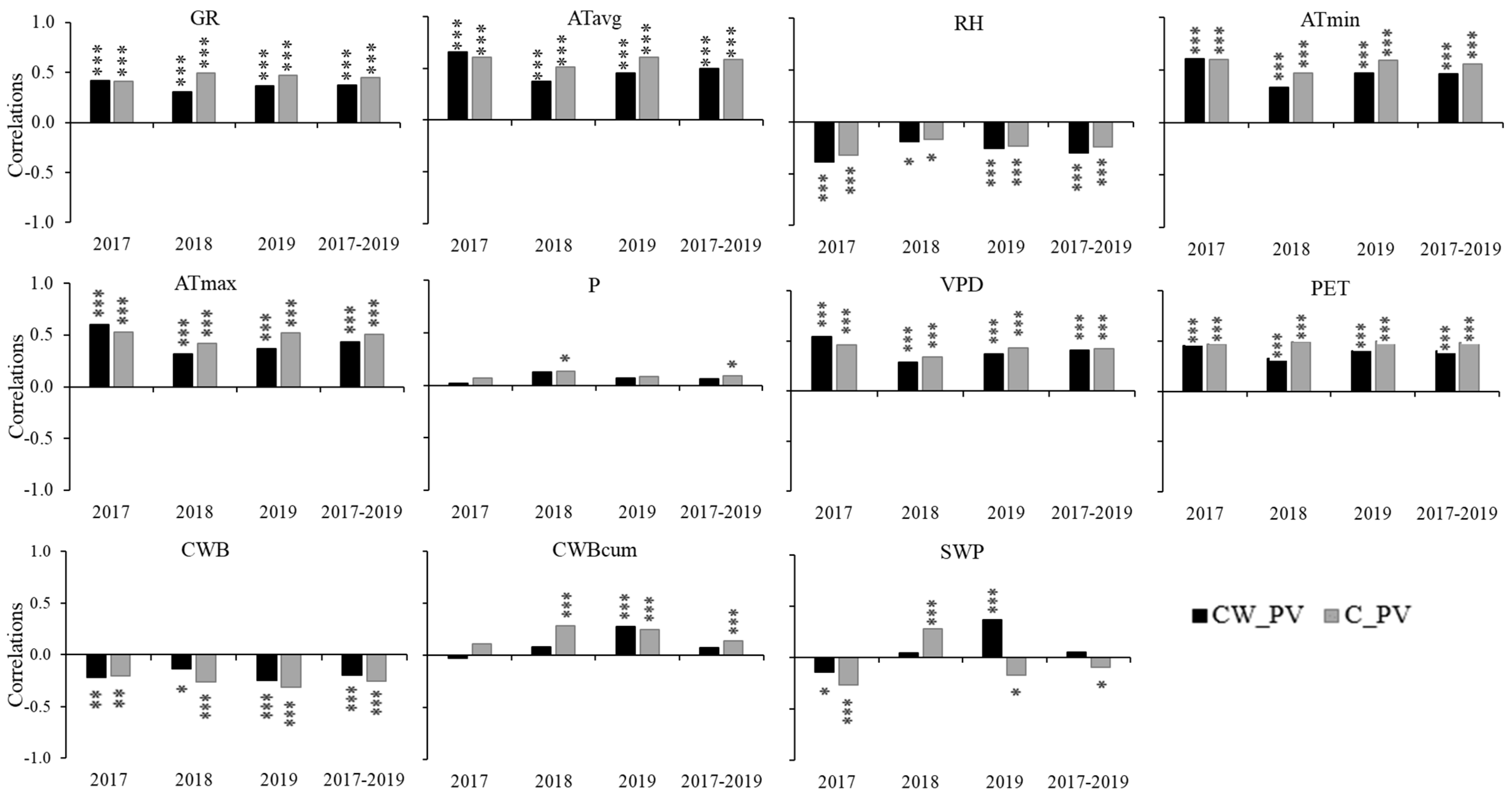
3.3. Influence of Environmental Variables on Growth and Tree Water Status
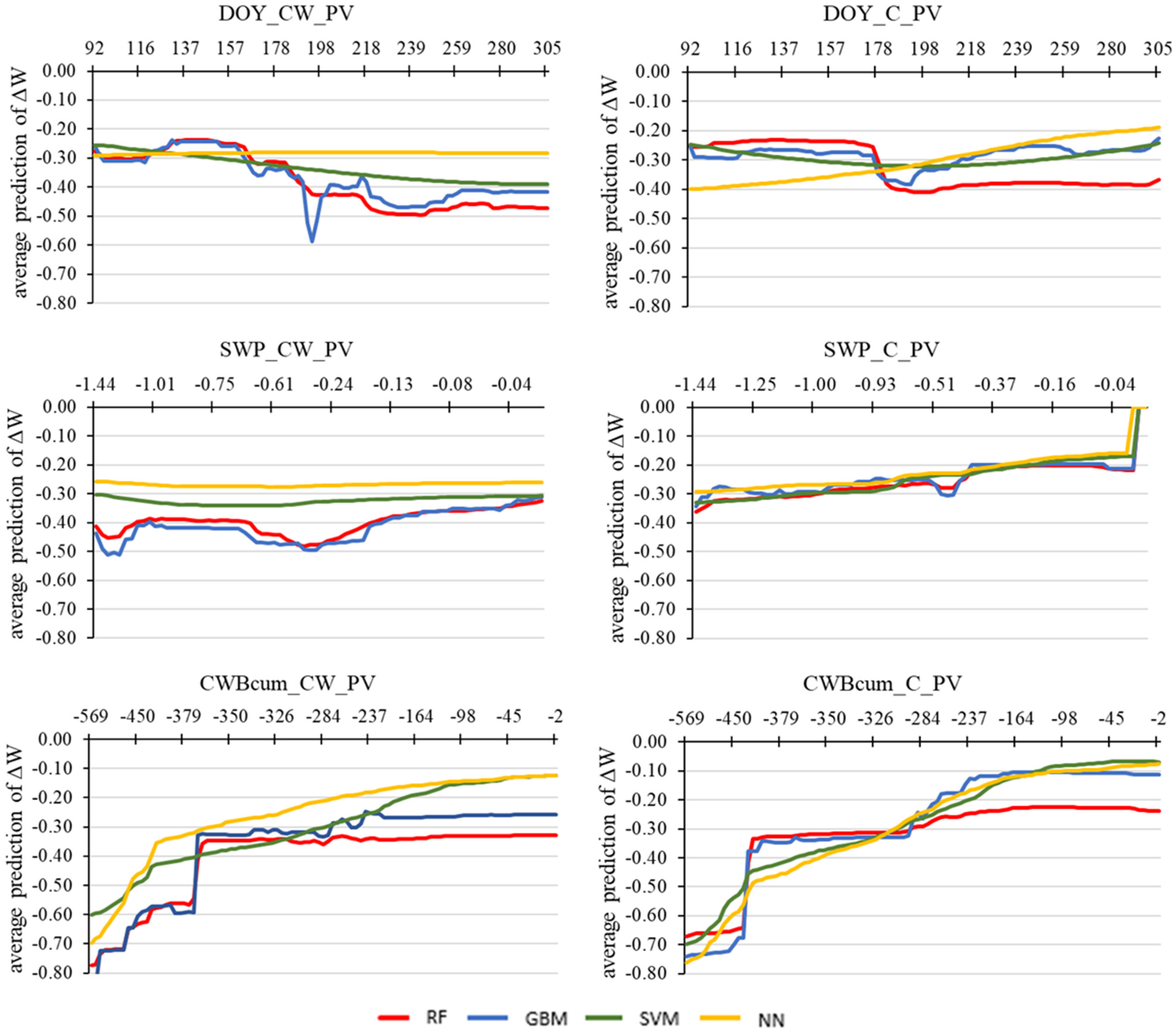
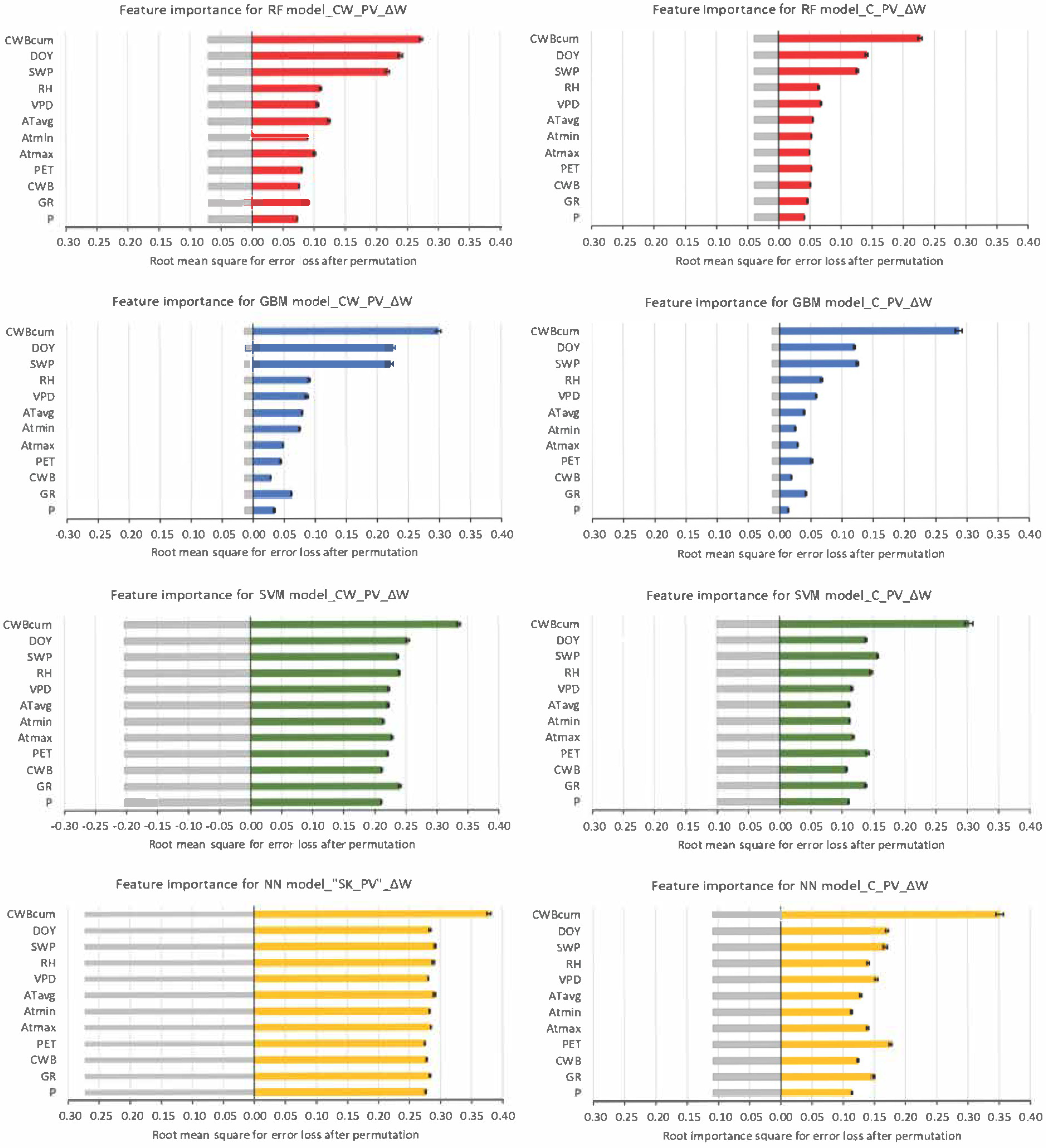
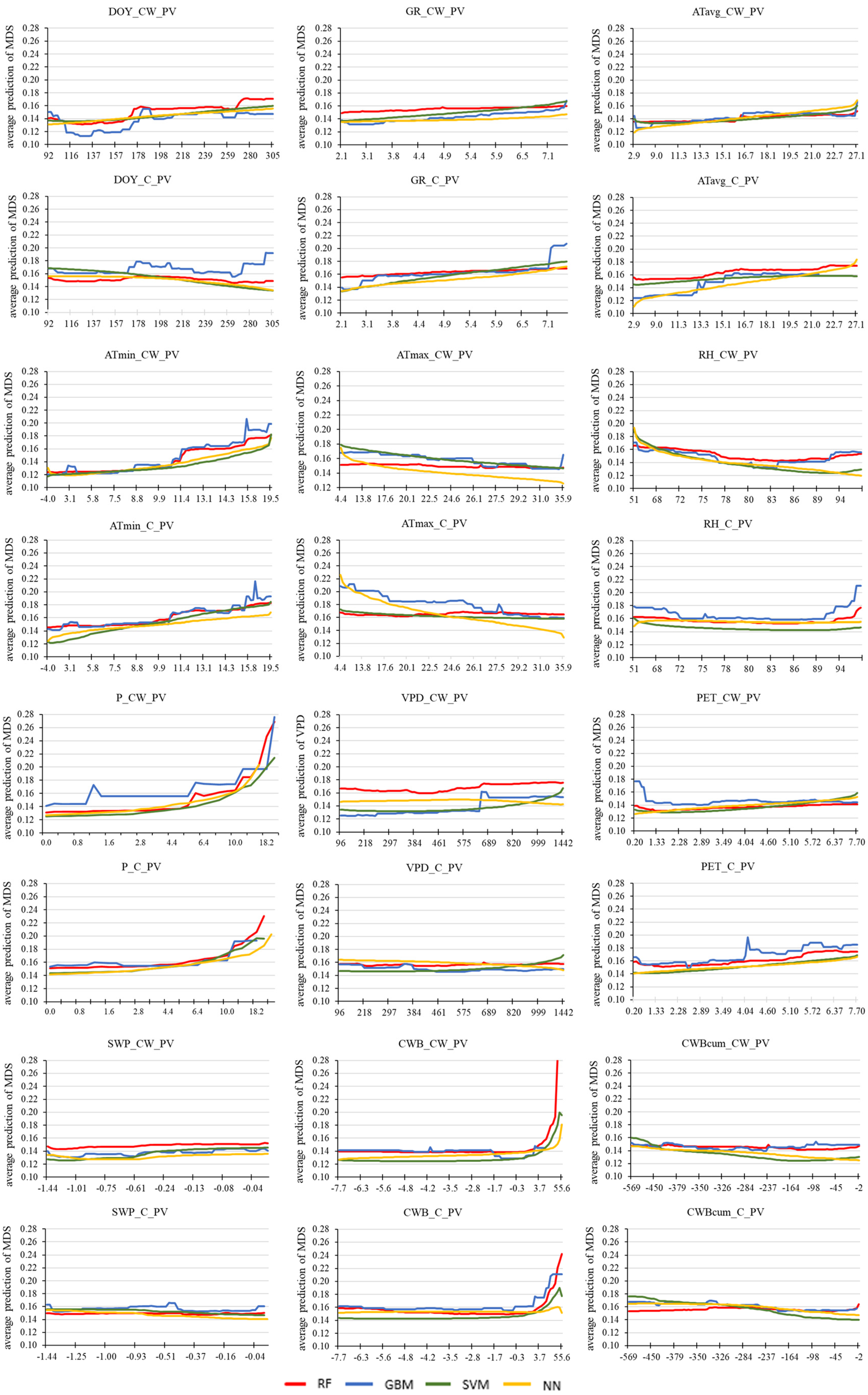
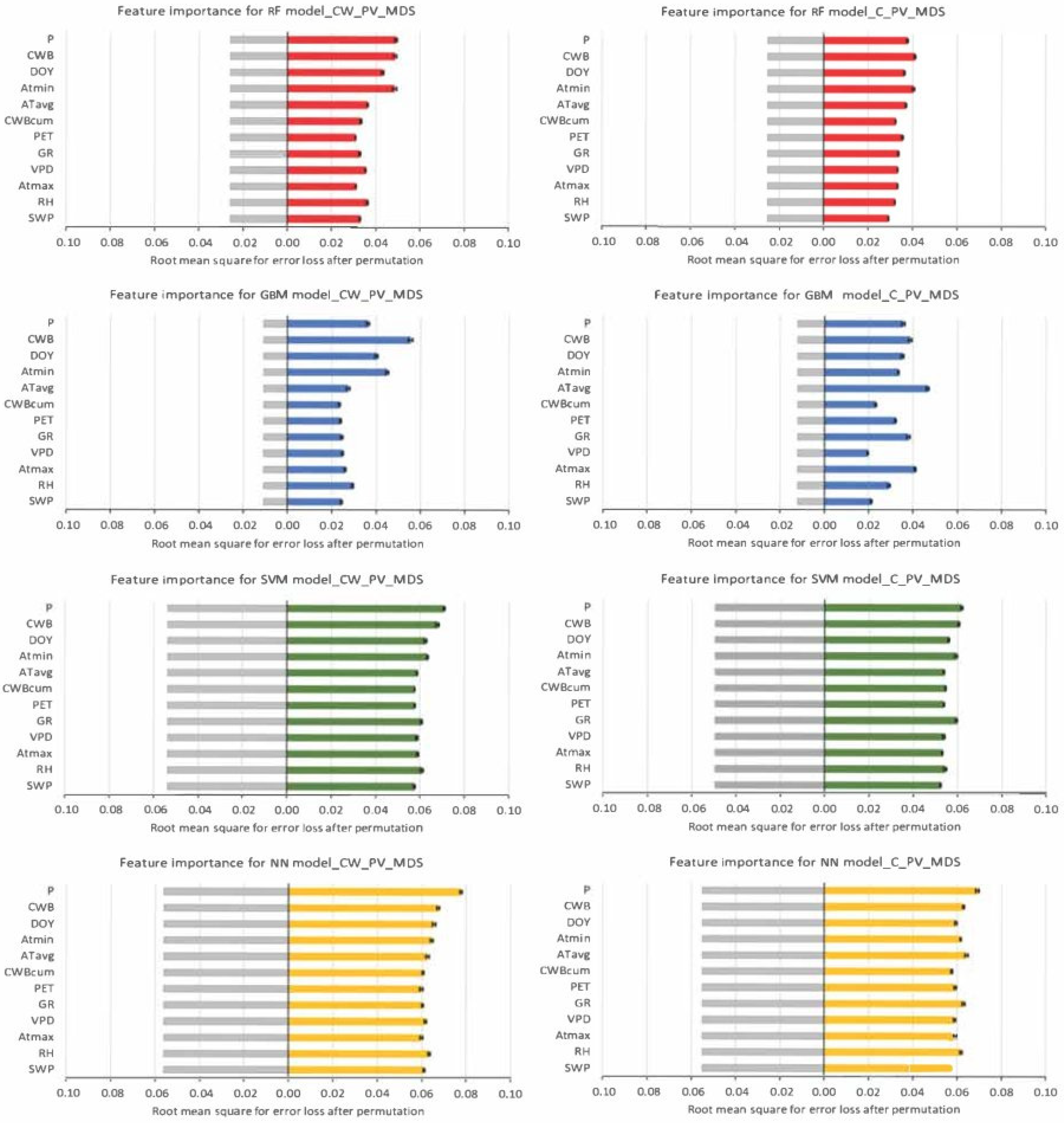
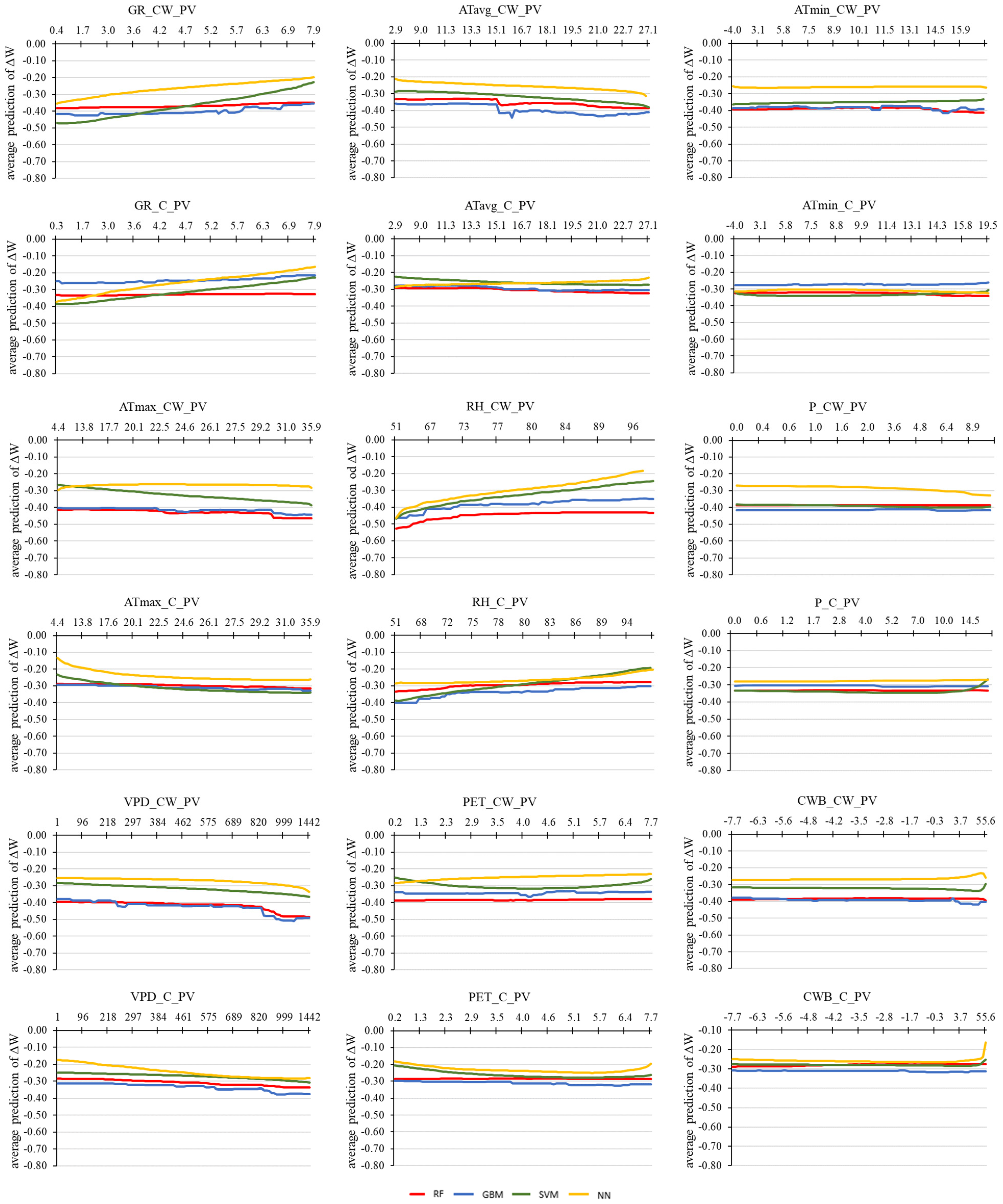
3.4. Machine Learning Techniques for Detecting the Influence of Environmental Factors on the Predicted Tree Water Status Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
- Random forest model development
- Gradient boosting machine model development
- Support Vector Machine model development
- Neural network model development
Appendix B


References
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Pšidová, E.; Živčák, M.; Stojnić, S.; Orlović, S.; Gömöry, D.; Kučerová, J.; Ditmarová, L’.; Střelcová, K.; Brestič, M.; Kalaji, H.M. Altitude of origin influences the responses of PSII photochemistry to heat waves in European beech (Fagus sylvatica L.). Environ. Exp. Bot. 2018, 152, 97–106. [Google Scholar] [CrossRef]
- Matyas, C. Climatic adaptation of trees: Rediscovering provenance tests. Euphytica 1996, 92, 45–54. [Google Scholar] [CrossRef]
- Czajkowski, T.; Bolte, A. Unteschiedliche Reaktionen deutscher und polnischer Herkunfte der Buche (Fagus sylvatica L.) auf Trockenheit. Allg. Forstu. J. Ztg. 2006, 177, 30–40. [Google Scholar]
- Rose, L.; Leuschner, C.; Köckemann, B.; Buschmann, H. Are marginal beech (Fagus sylvatica L.) provenances a source for drought tolerant ecotypes? Eur. J. For. Res. 2009, 128, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Castagneri, D.; Storaunet, K.O.; Rolstand, J. Age and growth patterns of old Norway spruce trees in Trillemarka forest, Norway. Scand. J. Forest Res. 2013, 28, 232–240. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea abies in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e012300+. [Google Scholar]
- Spiecker, H. Growth of Norway spruce (Picea abies [L.] Karst.) under changing environmental conditions in Europe. In Spruce Monocultures in Central Europe—Problems and Prospects, EFI Proceedings, Brno, Czech Republic, 22–25 June 1998; Klimo, E., Hager, H., Kulhavý, J., Eds.; European Forest Institute: Joensuu, Finland, 2000; Volume 33, pp. 11–27. [Google Scholar]
- Conedera, M.; Colombaroli, D.; Tinner, W.; Krebs, P.; Whitlock, C. Insights about past forest dynamics as a tool for present and future forest management in Switzerland. For. Ecol. Manag. 2017, 388, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Carrer, M.; Motta, R.; Nola, P. Significant mean and extreme climate sensitivity of Norway spruce and silver fir at mid-elevation mesic sites in the Alps. PLoS ONE 2012, 7, e50755. [Google Scholar] [CrossRef] [PubMed]
- Kahle, H.P.; Unseld, R.; Spiecker, H. Forest Ecosystems in a Changing Environment: Growth Patterns as Indicators for Stability of Norway Spruce within and Beyond the Limits of its Natural Range. In Application and Analysis of the Map of the Natural Vegetation of Europe; Bohn, U., Hettwer, C., Gollub, G., Eds.; Bundesamt für Naturschutz: Bonn, Germany, 2005; pp. 399–409. [Google Scholar]
- Skrøppa, T. EUFORGEN-Technical Guidelines for Genetic Conservation and Use for Norway Spruce (Picea abies); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Tužinský, L.; Bublinec, E.; Tužinský, M. Development of soil water regime under spruce stands. Folia Oecol. 2017, 44, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Siegwolf, R.T.W.; Cotrufo, M.F. Tree rings indicate different drought resistance of a native (Abies alba Mill.) and a non-native (Picea abies (L.) Karst.) species co-occurring at a dry site in Southern Italy. For. Ecol. Manag. 2009, 257, 820–828. [Google Scholar] [CrossRef]
- Usoltsev, V.A.; Merganičová, K.; Konôpka, B.; Tsepordey, I.S. The principle of space-for-time substitution in predicting Picea spp. biomass change under climate shifts. Centr. Eur. For. J. 2022, 68, 174–189. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Shrinking and swelling of plant tissues. In Water Deficits and Plant Growth; Kozlowski, T.T., Ed.; Academic Press: New York, NY, USA, 1976; pp. 1–64. [Google Scholar]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Zimmermann, L.; Newbery, D.M. Modelling tree water deficit from microclimate: An approach to quantifying drought stress. Tree Physiol. 2005, 25, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Mäkinen, H.; Nöjd, P.; Saranpää, P. Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiol. 2003, 23, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. For. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Swidrak, I.; Gruber, A.; Oberhuber, W. Xylem and phloem phenology in co-occurring conifers exposed to drought. Trees-Struct. Funct. 2014, 28, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Steppe, K.; De Pauw, D.J.W.; Lemeur, R.; Vanrolleghem, P.A. A mathematical model linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth. Tree Physiol. 2006, 26, 257–273. [Google Scholar] [CrossRef] [Green Version]
- Deslauriers, A.; Rossi, S.; Anfondillo, T. Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia 2007, 25, 113–124. [Google Scholar] [CrossRef]
- Oberhuber, W.; Hammerle, A.; Kofler, W. Tree water status and growth of saplings and mature Norway spruce (Picea abies) at a dry distribution limit. Front. Plant Sci. 2015, 6, 703. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, C.; Rötzer, T.; Thurm, E.A.; Biber, P.; Kallenbach, C.; Pretzsch, H. Growth and Tree Water Deficit of Mixed Norway Spruce and European Beech at Different Heights in a Tree and under Heavy Drought. Forests 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Herzog, K.M.; Thum, R.; Häsler, R. Diurnal changes in the radius of a subalpine Norway spruce stem: Their relation to the sap flow and their use to estimate transpiration. Trees 1995, 10, 94–101. [Google Scholar] [CrossRef]
- Offenthaler, I.; Hietz, P.; Richter, H. Wood diameter indicates diurnal and long-term patterns of xylem water potential in Norway spruce. Trees 2001, 15, 215–221. [Google Scholar] [CrossRef]
- Downes, G.; Beadle, C.; Worledge, D. Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 1999, 14, 102–111. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Quebec (Canada). Trees 2003, 17, 477–484. [Google Scholar] [CrossRef]
- Bouriaud, O.; Bréda, N.; Dupouey, J.L.; Granier, A. Is ring width a reliable proxy for stem-biomass increment? A case study in European beech. Can. J. For. Res. 2005, 35, 2920–2933. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Seasonal and daily cycles of stem radial variation of pinus pinaster in a drought-prone environment. Agric. For. Meteorol. 2013, 180, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Ježík, M.; Blaženec, M.; Letts, M.G.; Ditmarová, L.; Sitková, Z.; Strelcová, K. Assessing seasonal drought stress response in Norway spruce (Picea abies (L.) Karst.) by monitoring stem circumference and sap flow. Ecohydrology 2015, 8, 378–386. [Google Scholar] [CrossRef]
- Vospernik, S.; Nothdurft, A. 2018: Can trees at high elevations compensate for growth reductions at low elevations due to climate warning? Can. J. For. Res. 2018, 48, 650–662. [Google Scholar] [CrossRef]
- Van der Maaten, E. Thinning prolongs growth duration of European beech (Fagus sylvatica L.) across a valley in southwestern Germany. For. Ecol. Manag. 2013, 306, 135–141. [Google Scholar] [CrossRef]
- Kalliokoski, T.; Reza, M.; Jyske, T.; Mäkinen, H.; Nöjd, P. Intra-annual tracheid formation of Norway spruce provenances in southern Finland. Trees-Struct. Funct. 2012, 26, 543–555. [Google Scholar] [CrossRef]
- Lukáčik, I. Arborétum Borová hora—História, súčasnost’ a perspektívy. In Dendroflora of Central Europe—Utilization of Knowledge in Research, Education and Practice; Lukáčik, I., Sarvašová, I., Eds.; Vydavatel’stvo TU vo Zvolene: Zvolen, Slovakia, 2015; pp. 9–19. [Google Scholar]
- Jones, P.D.; Lister, D.H.; Osborn, T.J.; Harpham, C.; Salmon, M.; Morice, C.P. Hemispheric and large-scale land surface air temperature variations: An extensive revision and an update to 2010. J. Geophys. Res. 2012, 117, D05127. [Google Scholar] [CrossRef] [Green Version]
- Penman, H.L. Natural evaporation from open water, bare soil, and grass. Proc. R. Soc. 1948, A193, 120–146. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; p. D05109. [Google Scholar]
- Ehrenberger, W.; Rüger, S.; Fitzke, R.; Vollenweider, P.; Günthardt-Goerg, M.S.; Kuster, T.; Zimmermann, U.; Arend, M. Concomitant dendrometer and leaf patch pressure probe measurements reveal the effect of microclimate and soil moisture on diurnal stem water and leaf turgor variations in young oak trees. Funct. Plant. Biol. 2012, 39, 297–305. [Google Scholar] [CrossRef]
- Van der Maaten, E.; van der Maaten-Theunissen, M.; Smiljani´c, M.; Rossi, S.; Simard, S.; Wilmking, M.; Deslauriers, A.; Fonti, P.; von Arx, G.; Bouriaud, O. dendrometeR: Analyzing the pulse of tree in R. Dendrochronologia 2016, 40, 12–16. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? N. Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Alzubi, J.; Nayyar, A.; Kumar, A. Machine Learning from Theory to Algorithms: An Overview. J. Phys. Conf. Ser. 2018, 1142, 012012. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.; Stone, C.J.; Olshen, R.A. Classification and Regression Trees; CRC Press: Boca Raton, FL, USA, 1984. [Google Scholar]
- Sarica, A.; Cerasa, A.; Quattrone, A. Random Forest Algotithm fot the Classification of Neuroimaging Data in Alzheimer’s Disease: A Systematic Review. Front. Aging Neurosci. 2017, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Cha, G.W.; Moon, H.J.; Kim, Y.C. Comparison of Random Forest and Gradient Boosting Machine Models for Predicting Demolition Waste Based on Small Datasete and Categorical Variables. Int. J. Environ. Res. Public Health 2021, 18, 8530. [Google Scholar] [CrossRef]
- López, O.A.M.; López, A.M.; Crossa, J. Support Vector Machine and Support Vector Regression. In Multivariate Statistical Machine Learning Methods for Genomic Prediction; Springer: Cham, Switzerland, 2022. [Google Scholar]
- López, O.A.M.; López, A.M.; Crossa, J. Fundamentals of Artificial Neural Networks and Deep Learning. In Multivariate Statistical Machine Learning Methods for Genomic Prediction; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Novak, R.; Bahri, Y.; Abolafia, D.A.; Pennington, J.; Sohl-Dickstein, J. Sensitivity and generalization in neural networks: An empirical study. arXiv 2018, arXiv:1802.08760. [Google Scholar]
- Elgeldawi, E.; Sayed, A.; Galal, A.R.; Zaki, A.M. Hyperparameter Tuning for Machine Learning Algorithms Used for Arabic Sentiment Analysis. Informatics 2021, 8, 79. [Google Scholar] [CrossRef]
- Kuhn, M. Caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar]
- Biecek, P. DALEX: Explainers for Complex Predictive Models in R. J. Mach. Learn. Res. 2018, 19, 1–5. [Google Scholar]
- Biecek, P.; Burzykowski, T. Explanatory Model Analysis: Explore, Explain and Examine Predictive Models; Chapman and Hall/CRC: Boca Raton, FL, USA, 2021. [Google Scholar]
- Schmidt-Vogt, H. Die Fichte; Verlag Paul Parey: Hamburg-Berlin, Germany, 1977; p. 647. [Google Scholar]
- Spiecker, H. Silvicultural management in maintaining biodiversity and resistance of forests in Europe—Temperate zone. J. Environ. Manag. 2013, 67, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Boshier, D.; Broadhurst, L.; Cornelius, J.; Gallo, L.; Koskela, J.; Loo, J.; Petrokofsky, G.; Clair, B.S. Is local best? Examining the evidence for local adaptation in trees and its scale. Environ. Evid. 2015, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.; Pluess, A.R.; Howe, G.T.; Sperisen, C.; Heiri, C. Quantitative genetic differentiation and phenotypic plasticity of European beech in a heterogeneous landscape: Indications for past climate adaptation. Perspect. Plant Ecol. Evol. Syst. 2017, 26, 1–13. [Google Scholar] [CrossRef]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christine, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P.; et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Chang. Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. N. Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Suvanto, S.; Henttonen, H.M.; Nojd, P.; Makinen, H. Forest susceptibility to storm damage is affected by similar factors regardless of storm type: Comparison of thunder storms and autumn extra-tropical cyclones in Finland. For. Eco. Man. 2016, 381, 17–28. [Google Scholar] [CrossRef]
- Solberg, S. Summer drought: A driver for crown condition and mortality of Norway spruce in Norway. For. Pathol. 2004, 34, 93–104. [Google Scholar] [CrossRef]
- Cienciala, E.; Altman, J.; Doležal, J.; Kopáček, J.; Štěpánek, P.; Ståhl, G.; Tumajer, J. Increased Spruce Tree Growth in Central Europe Since 1960s. Sci. Total Environ. 2018, 619–620, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Schurman, J.S.; Babst, F.; Björklund, J.; Rydval, M.; Bače, R.; Čada, V.; Janda, P.; Mikolas, M.; Saulnier, M.; Trotsiuk, V.; et al. The climatic drivers of primary Picea forest growth along the Carpathian arc are changing under rising temperatures. Glob. Chang. Biol. 2019, 25, 3136–3150. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolář, T.; Čermák, P.; Oulehle, F.; Trnka, M.; Štěpánek, P.; Cudlín, P.; Hruška, J.; Büntgen, U.; Rybníček, M. Pollution control enhanced spruce growth in the “Black Triangle” near the Czech-Polish border. Sci. Total Environ. 2015, 538, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Reyer, C.P.O.; Rigaud, K.K.; Fernandes, E.; Hare, W.; Serdeczny, O.; Schellnhuber, H.J. Turn down the heat: Regional climate change impacts on development. Reg. Environ. Chang. 2017, 17, 1563–1568. [Google Scholar] [CrossRef] [Green Version]
- Bošeľa, M.; Kulla, L.; Roessiger, J.; Šebeň, V.; Dobor, L.; Büntgen, U.; Lukac, M. Long-term effects of environmental change and species diversity on tree radial growth in a mixed European forest. For. Ecol. Manag. 2019, 446, 293–303. [Google Scholar] [CrossRef]
- Pretzsch, H.; Ďurský, J. Growth reaction of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus silvatica L.) to possible climatic changes in Germany. A sensitivity study. Forstwiss. Centralbl. 2002, 121, 145–154. [Google Scholar]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Lévesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Bottero, A.; Forrester, D.I.; Cailleret, M.; Kohnle, U.; Gessler, A.; Michel, D.; Bose, A.K.; Bauhus, J.; Bugmann, H.; Cuntz, M.; et al. Growth resistance and resilience of mixed silver fir and Norway spruce forests in central Europe: Contrasting responses to mild and severe droughts. Glob. Chang. Biol. 2021, 27, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Børja, I.; De Wit, H.A.; Steffenrem, A.; Majdi, H. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiol. 2008, 28, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, S.; He, F. Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate. Proc. Natl. Acad. Sci. USA 2015, 112, 4009–4014. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Huang, J.G.; Cheng, J.; Dawson, A.; Stadt, K.J.; Comeau, P.G.; Chen, H.Y.H. Interspecific variation in growth responses to tree size, competition and climate of western Canadian boreal mixed forests. Sci. Total Environ. 2018, 631–632, 1070–1078. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, J.; Zhou, J.; Guan, C.; Lei, S.; Meng, P.; Yin, C. Long-Term Effects of Climate and Competition on Radial Growth, Recovery, and Resistance in Mongolian Pines. Front Plant Sci. 2021, 12, 729935. [Google Scholar] [CrossRef]
- Schuster, R.; Oberhuber, W. Drought sensitivity of three co-occurring conifers within a dry inner Alpine environment. Trees 2013, 27, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Lyr, H.; Fiedler, H.J.; Tranquillini, W. Physiologie und Ökologie der Gehölze; G. Fischer Verlag: Jena, Germany, 1992. [Google Scholar]
- Cochard, H. Vulnerability of several conifers to air embolism. Tree Physiol. 1992, 11, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Mayr, S.; Bardage, S.; Brändström, J. Hydraulic and anatomical properties of light bands in Norway spruce compression wood. Tree Physiol. 2006, 26, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodribb, T.J.; Bowman, D.J.; Nichols, S.; Delzon, S.; Burlett, R. Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit. N. Phytol. 2010, 188, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Drew, D.M.; Richards, A.E.; Downes, G.M.; Cook, G.D.; Baker, P. The development of seasonal tree water deficit in Callitris intratropica. Tree Physiol. 2011, 31, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Köcher, P.; Horna, V.; Leuschner, C. Stem water storage in five coexisting temperate broad-leaved tree species: Significance, temporal dynamics and dependence on tree functional traits. Tree Physiol. 2013, 33, 817–832. [Google Scholar] [CrossRef]
- Balducci, L.; Deslauriers, A.; Giovannelli, A.; Rossi, S.; Rathgeber, C.B.K. Effects of temperature and water deficit on cambial activity and woody ring features in Picea mariana saplings. Tree Physiol. 2013, 33, 1006–1017. [Google Scholar] [CrossRef]
- Daudet, F.A.; Améglio, T.; Cochard, H.; Archilla, O.; Lacointe, A. Experimental analysis of the role of water and carbon in tree stem diameter variations. J. Exp. Bot. 2005, 56, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, A.; Deslauriers, A.; Fragnelli, G.; Scaletti, L.; Castro, G.; Rossi, S.; Crivellaro, A. Evaluation of drought response of two poplar clones (Populus x canadensis Monch ’I-214’ and P. deltoides Marsh. ’Dvina’) through high resolution analysis of stem growth. J. Exp. Bot. 2007, 58, 2673–2683. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Item, H.; Häsler, R. Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees 2000, 15, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Item, H.; Häsler, R. Link between diurnal stem radius changes and tree water relations. Tree Physiol. 2001, 21, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Palacio, S.; Hoch, G.; Sala, A.; Körner, C.; Millard, P. Does carbon storage limit tree growth? N. Phytol. 2014, 4, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Wang, D.; Ma, X.; Li, H. Predicting Short-Term Subway Ridership and Prioritizing Its Influential Factors Using Gradient Boosting Decision Trees. Sustainability 2016, 8, 1100. [Google Scholar] [CrossRef] [Green Version]
- Matsuki, K.; Kuperman, V.; Van Dyke, J.A. The Random Forests statistical technique: An examination of its value for the study of reading. Sci. Stud. Read. 2016, 20, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Bornschein, J.; Visin, F.; Osindero, S. Small Data, Big Decisions: Model Selection in the Small-Data Regime. Proc. 37th Int. Conf. Mach. Learn. 2020, 119, 1035–1044. [Google Scholar]
- Nakkiran, P.; Kaplun, G.; Bansal, Y.; Yang, T.; Barak, B.; Sutskever, I. Deep double descent: Where bigger models and more data hurt. J. Stat. Mech. 2021, 2021, 124003. [Google Scholar] [CrossRef]
- Kraus, M.; Feuerriegel, S.; Oztekin, A. Deep learning in business analytics and operations research: Models, applications and managerial implications. Eur. J. Oper. Res. 2020, 281, 628–641. [Google Scholar] [CrossRef]
- Philipp, P.; Wright, M.N.; Boulesteix, A.-L. Hyperparameters and tuning strategies for random forest. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2019, 9, e1301. [Google Scholar]
- Abadi, M.; Agarwal, A.; Barham, P.; Brevdo, E.; Chen, Z.; Citro, C.; Corrado, G.S.; Davis, A.; Dean, J.; Devin, M.; et al. TensorFlow: Large-Scale Machine Learning on Heterogeneous Systems. 2015. Available online: https://tensorflow.org (accessed on 11 January 2023).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Study Area | Provenance | |
|---|---|---|---|
| CW_PV | C_PV | ||
| Orographic Unit | Zvolen valley | Podtatranská valley | Archangeľskaja and Volgogradskaja region |
| Elevation (m a. s. l.) | 350 | 800 | 33–117 |
| PA (mm) | 651 | 833 | 587 |
| PGS (mm) | 422 | 603 | 404 |
| TA (°C) | 7.9 | 5.3 | 2 |
| TGS (°C) | 13.6 | 10.7 | 9.5 |
| Year (Months) | P | ATavg | GR | RH | VPD | PET | CWBcum | SWP_CW_PV | SWP_C_PV |
|---|---|---|---|---|---|---|---|---|---|
| (mm) | (°C) | (kWh.m−2) | (%) | (kPa) | (mm) | (mm) | (MPa) | (MPa) | |
| 2017 (A-O) | 501 | 15.2 | 977 | 81 | 0.500 | 825 | −324 | −0.463 | −0.116 |
| 2018 (A-O) | 321 | 17.0 | 985 | 80 | 0.557 | 877 | −556 | −1.045 | −0.427 |
| 2019 (A-O) | 387 | 15.8 | 927 | 81 | 0.480 | 805 | −418 | −0.996 | −0.374 |
| CW_PV | C_PV | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Icum | SD± | ∆Wcum | SD± | MDScum | SD± | Icum | SD± | ∆Wcum | SD± | MDScum | SD± | |
| 2017 | 6.3 | 3.8 | −43.2 | 13.7 | 31.2 | 14.6 | 8.6 | 3.0 | −30.4 | 13.3 | 30.9 | 9.9 |
| 2018 | 3.7 | 2.5 | −95.7 | 35.1 | 29.2 | 13.6 | 4.2 | 2.8 | −107.8 | 63.2 | 35.5 | 11.4 |
| 2019 | 3.2 | 1.7 | −118.9 | 25.6 | 29.7 | 12.2 | 6.0 | 3.8 | −60.0 | 30.2 | 32.4 | 13.7 |
| 2017–2019 | 13.2 | −257.8 | 90.1 | 18.8 | −198.2 | 98.8 | ||||||
| RF | GBM | SVM | NN | ||
|---|---|---|---|---|---|
| ΔW_”CW_PV” | MSE | 0.005 | 0.0002 | 0.040 | 0.040 |
| RMSE | 0.070 | 0.0159 | 0.201 | 0.200 | |
| R2 | 0.956 | 0.963 | 0.638 | 0.642 | |
| MAD | 0.023 | 0.002 | 0.045 | 0.117 | |
| ΔW_”C_PV” | MSE | 0.002 | 0.0001 | 0.009 | 0.013 |
| RMSE | 0.039 | 0.011 | 0.097 | 0.113 | |
| R2 | 0.979 | 0.97 | 0.868 | 0.818 | |
| MAD | 0.017 | 0.002 | 0.027 | 0.051 | |
| MDS_”CW_PV” | MSE | 0.001 | 0.0002 | 0.002 | 0.003 |
| RMSE | 0.028 | 0.0164 | 0.054 | 0.055 | |
| R2 | 0.902 | 0.968 | 0.629 | 0.610 | |
| MAD | 0.013 | 0.002 | 0.018 | 0.028 | |
| MDS_”C_PV” | MSE | 0.001 | 0.0003 | 0.002 | 0.003 |
| RMSE | 0.025 | 0.019 | 0.050 | 0.051 | |
| R2 | 0.898 | 0.952 | 0.607 | 0.587 | |
| MAD | 0.011 | 0.004 | 0.017 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leštianska, A.; Fleischer, P., Jr.; Merganičová, K.; Fleischer, P., Sr.; Nalevanková, P.; Střelcová, K. Effect of Provenance and Environmental Factors on Tree Growth and Tree Water Status of Norway Spruce. Forests 2023, 14, 156. https://doi.org/10.3390/f14010156
Leštianska A, Fleischer P Jr., Merganičová K, Fleischer P Sr., Nalevanková P, Střelcová K. Effect of Provenance and Environmental Factors on Tree Growth and Tree Water Status of Norway Spruce. Forests. 2023; 14(1):156. https://doi.org/10.3390/f14010156
Chicago/Turabian StyleLeštianska, Adriana, Peter Fleischer, Jr., Katarína Merganičová, Peter Fleischer, Sr., Paulína Nalevanková, and Katarína Střelcová. 2023. "Effect of Provenance and Environmental Factors on Tree Growth and Tree Water Status of Norway Spruce" Forests 14, no. 1: 156. https://doi.org/10.3390/f14010156
APA StyleLeštianska, A., Fleischer, P., Jr., Merganičová, K., Fleischer, P., Sr., Nalevanková, P., & Střelcová, K. (2023). Effect of Provenance and Environmental Factors on Tree Growth and Tree Water Status of Norway Spruce. Forests, 14(1), 156. https://doi.org/10.3390/f14010156