
Urbanization Imprint on Soil Bacterial Communities in Forests and Grasslands


Abstract
:1. Introduction
2. Materials and Methods
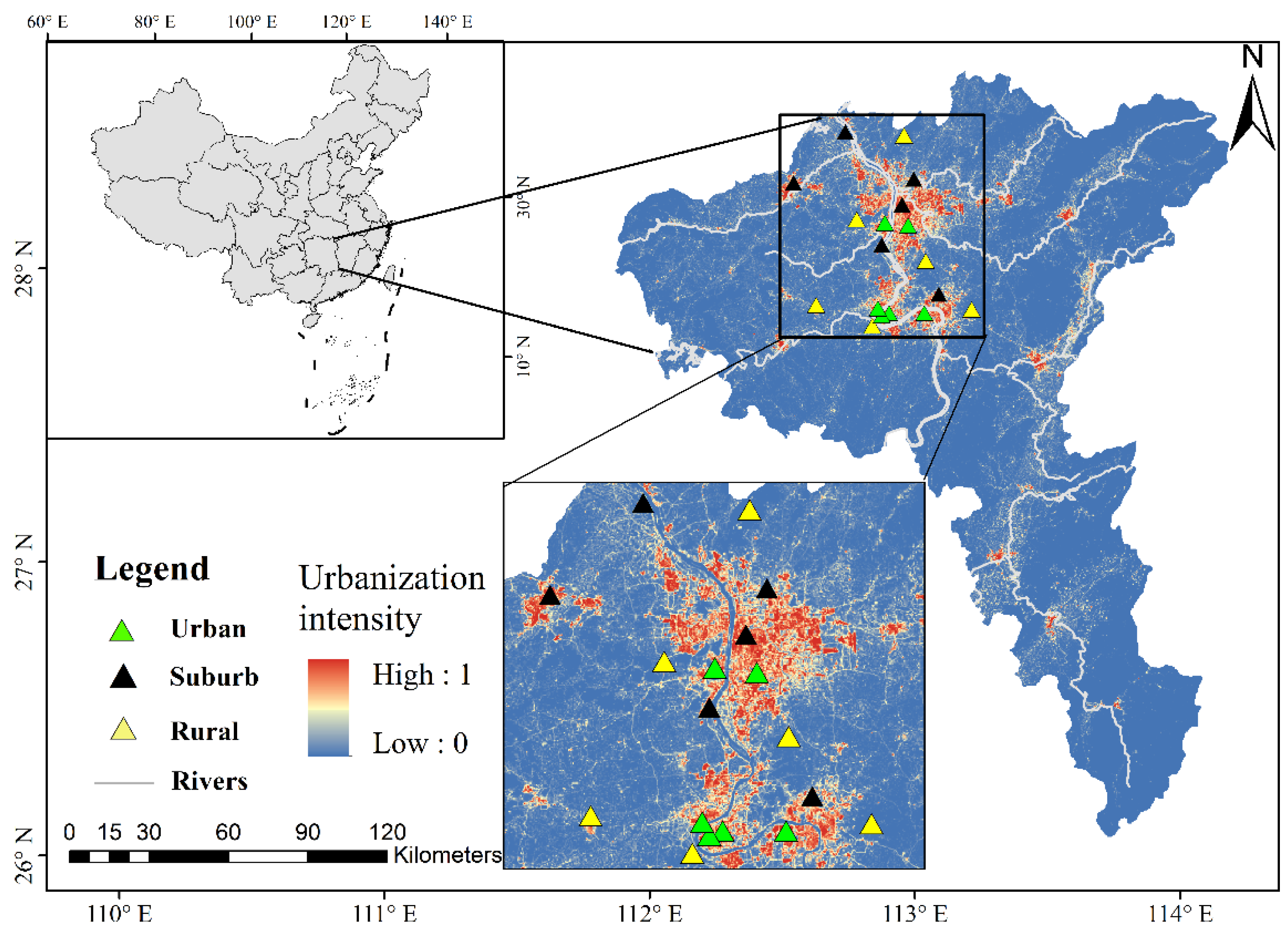
2.1. Soil Sampling and Soil Properties
2.2. DNA Extraction, Amplification and Sequencing
2.3. Statistical Analysis
3. Results
3.1. Soil Properties along the Rural-Suburban-Urban Gradient
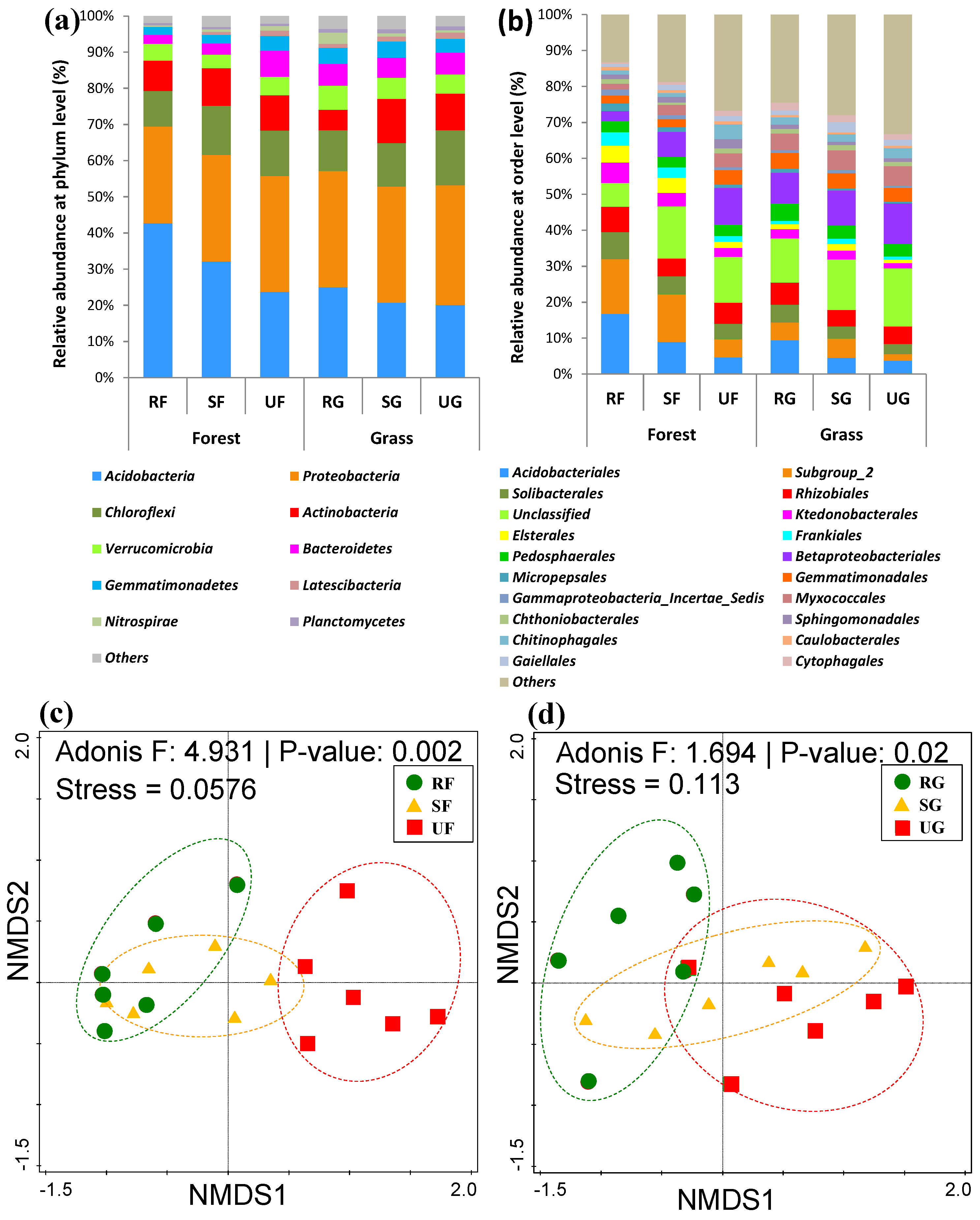
3.2. Impact of Urbanization on Soil Bacterial Diversity
3.3. The Responses of Soil Bacterial Community Compositions to Urbanization
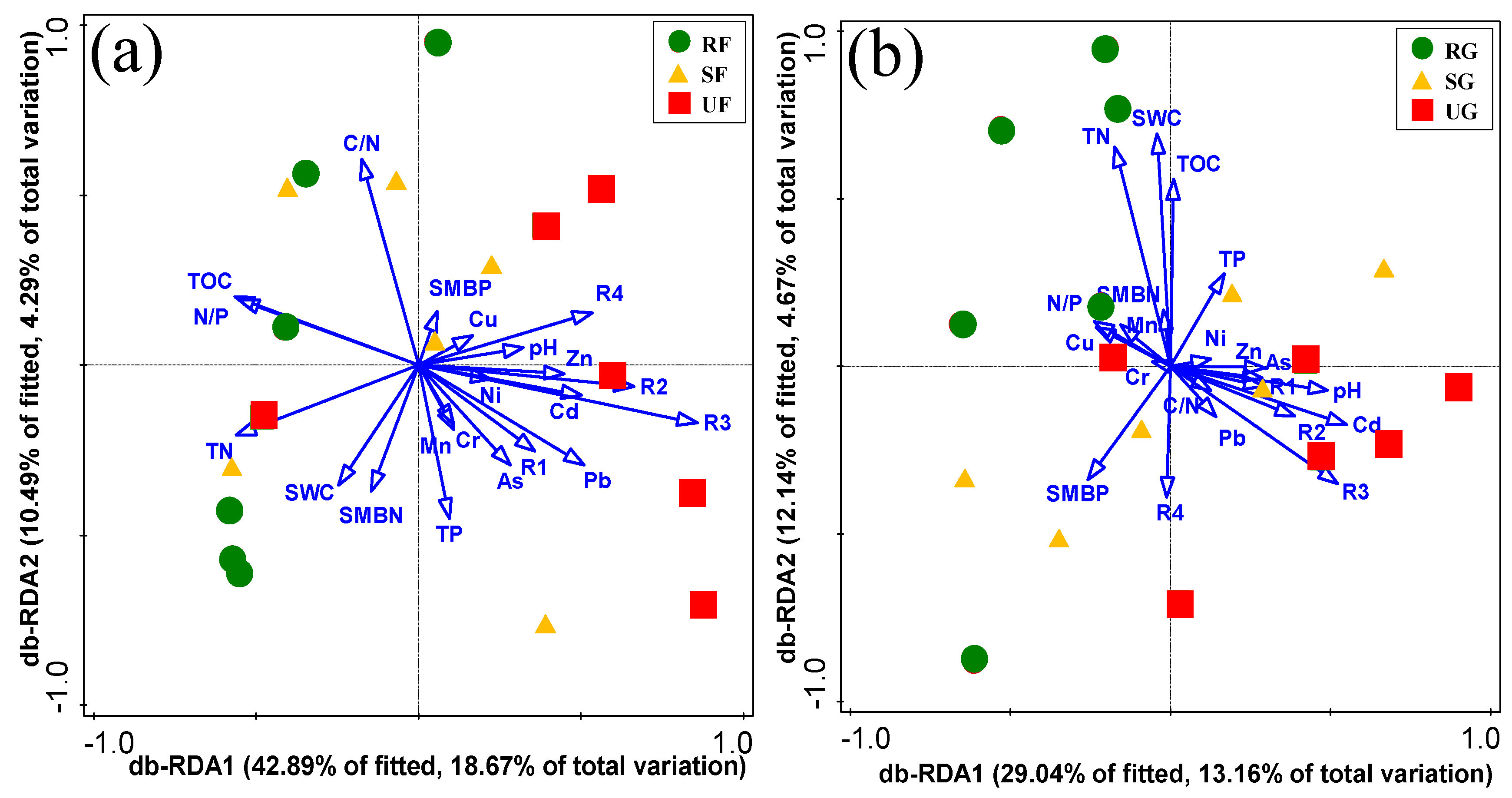
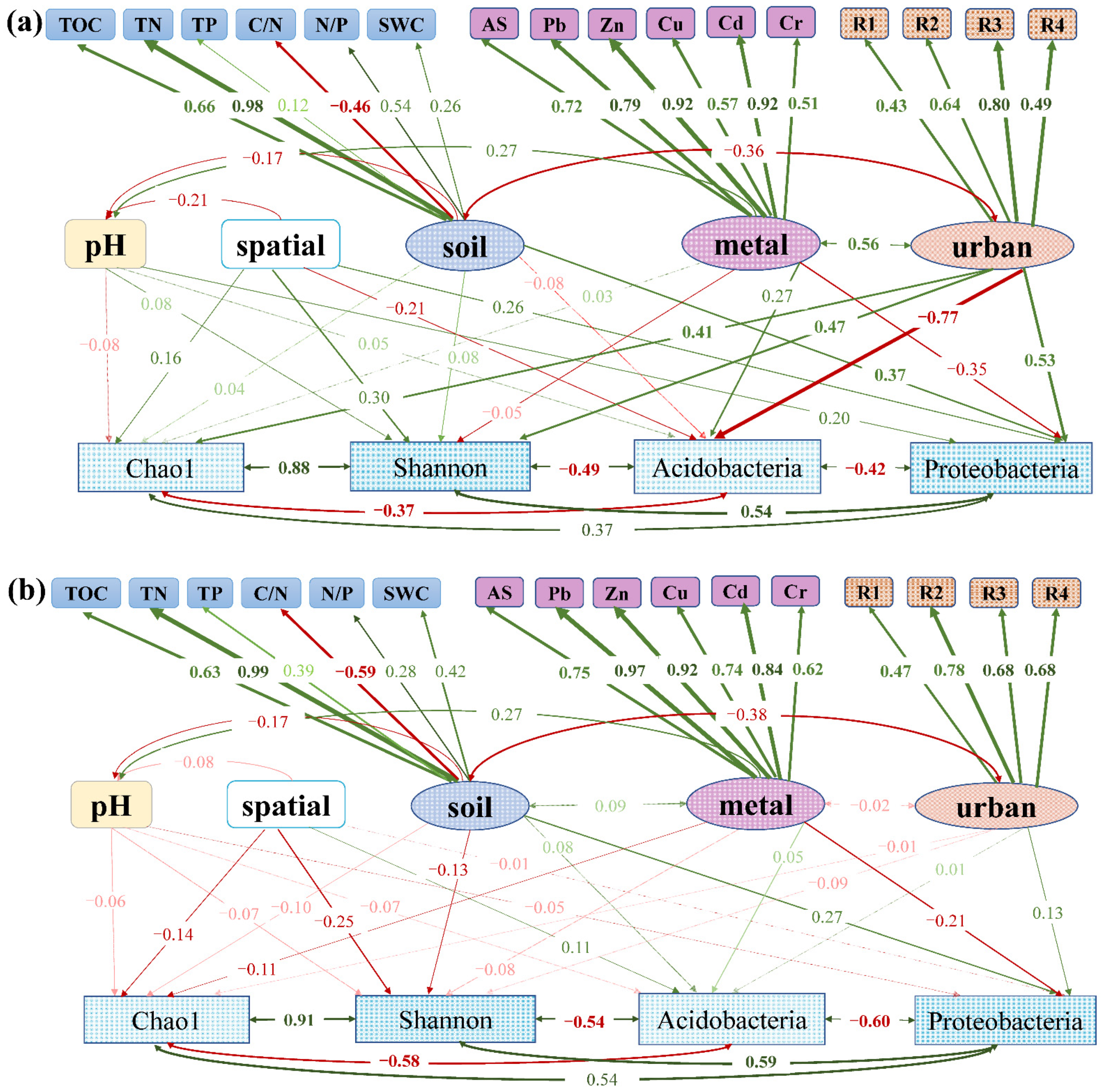
3.4. Relationship between Environmental Factors and Soil Bacterial Community
4. Discussion
4.1. The Influence of Urbanization on Diversity and Composition of Bacterial Communities
4.2. Differences in the Composition and Diversity of Bacterial Communities between Forests and Grasslands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Cheng, M.; Dsouza, M.; Weisenhorn, P.; Zheng, T. Soil Bacterial Diversity Is Associated with Human Population Density in Urban Greenspaces. Environ. Sci. Technol. 2018, 52, 5115–5124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Zhou, D.; Zhu, C.; Qu, W.; Zhao, J.; Sun, Y.; Huang, D.; Wu, W.; Liu, S. Rates and patterns of urban expansion in China’s 32 major cities over the past three decades. Landsc. Ecol. 2015, 30, 1541–1559. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Kuang, W.; Wu, J.; Liu, J.; Tian, H. Industrial land expansion in rural China threatens environmental securities. Front. Environ. Sci. Eng. 2021, 15, 29. [Google Scholar] [CrossRef]
- Trentanovi, G.; von der Lippe, M.; Sitzia, T.; Ziechmann, U.; Kowarik, I.; Cierjacks, A. Biotic homogenization at the community scale: Disentangling the roles of urbanization and plant invasion. Divers. Distrib. 2013, 19, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Aronson, M.F.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.; Cilliers, S.; Clarkson, B. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Sol, D.; Bartomeus, I.; González-Lagos, C.; Pavoine, S. Urbanisation and the loss of phylogenetic diversity in birds. Ecol. Lett. 2017, 20, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Merckx, T.; Van Dyck, H. Urbanization-driven homogenization is more pronounced and happens at wider spatial scales in nocturnal and mobile flying insects. Glob. Ecol. Biogeogr. 2019, 28, 1440–1455. [Google Scholar] [CrossRef]
- Epp Schmidt, D.J.; Pouyat, R.; Szlavecz, K.; Setälä, H.; Kotze, D.J.; Yesilonis, I.; Cilliers, S.; Hornung, E.; Dombos, M.; Yarwood, S.A. Urbanization erodes ectomycorrhizal fungal diversity and may cause microbial communities to converge. Nat. Ecol. Evol. 2017, 1, 0123. [Google Scholar] [CrossRef]
- Stephanou, C.; Omirou, M.; Philippot, L.; Zissimos, A.M.; Christoforou, I.C.; Trajanoski, S.; Oulas, A.; Ioannides, I.M. Land use in urban areas impacts the composition of soil bacterial communities involved in nitrogen cycling. A case study from Lefkosia (Nicosia) Cyprus. Sci. Rep. 2021, 11, 8198. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, X.; Ning, C.; Feng, S.; Gao, H.; Zhao, J.; Liao, J.; Peng, Y.; Zhao, S.; Liu, S. Urbanization-induced environmental changes strongly affect wetland soil bacterial community composition and diversity. Environ. Res. Lett. 2022, 17, 014027. [Google Scholar] [CrossRef]
- Yan, B.; Li, J.; Xiao, N.; Qi, Y.; Fu, G.; Liu, G.; Qiao, M. Urban-development-induced changes in the diversity and composition of the soil bacterial community in Beijing. Sci. Rep. 2016, 6, 38811. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Kan, L.; Su, Z.; Liu, X.; Zhang, L. The composition and diversity of soil bacterial and fungal communities along an urban-to-rural gradient in South China. Forests 2019, 10, 797. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.C.; O’Riordan, R.; Stevens, C. Urban soil microbial community and microbial-related carbon storage are severely limited by sealing. J. Soils Sediments 2021, 21, 1455–1465. [Google Scholar] [CrossRef]
- Boeraeve, M.; Honnay, O.; Jacquemyn, H. Local abiotic conditions are more important than landscape context for structuring arbuscular mycorrhizal fungal communities in the roots of a forest herb. Oecologia 2019, 190, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Docherty, K.M.; Pearce, D.S.; Lemmer, K.M.; Hale, R.L. Distributing regionally, distinguishing locally: Examining the underlying effects of local land use on airborne bacterial biodiversity. Environ. Microbiol. 2018, 20, 3529–3542. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-J.; Li, S.; Su, J.-Q.; Nie, S.A.; Gibson, V.; Li, H.; Zhu, Y.-G. Does urbanization shape bacterial community composition in urban park soils? A case study in 16 representative Chinese cities based on the pyrosequencing method. FEMS Microbiol. Ecol. 2014, 87, 182–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Marshall, C.; Cheng, M.; Xu, H.; Li, H.; Yang, X.; Zheng, T. Changes in land use driven by urbanization impact nitrogen cycling and the microbial community composition in soils. Sci. Rep. 2017, 7, 44049. [Google Scholar] [CrossRef]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Using soil bacterial communities to predict physico-chemical variables and soil quality. Microbiome 2020, 8, 79. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, J.; Lu, J.; Sun, Y. Study on the influence of soil microbial community on the long-term heavy metal pollution of different land use types and depth layers in mine. Ecotoxicol. Environ. Saf. 2019, 170, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Mi, J.; Chu, P.; Cheng, J.; Zhang, L.; Pan, Q.; Xie, Y.; Bai, Y. Patterns and drivers of soil microbial communities along a precipitation gradient on the Mongolian Plateau. Landsc. Ecol. 2015, 30, 1669–1682. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Steenwerth, K.L.; Jackson, L.E.; Scow, K.M. Land use and climatic factors structure regional patterns in soil microbial communities. Glob. Ecol. Biogeogr. 2010, 19, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Xiang, D.; Veresoglou, S.D.; Chen, F.; Chen, Y.; Hao, Z.; Zhang, X.; Chen, B. Soil organic carbon and soil structure are driving microbial abundance and community composition across the arid and semi-arid grasslands in northern China. Soil Biol. Biochem. 2014, 77, 51–57. [Google Scholar] [CrossRef]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Zhalnina, K.; Dias, R.; de Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH determines microbial diversity and composition in the park grass experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef]
- Zhang, K.; Delgado-Baquerizo, M.; Zhu, Y.-G.; Chu, H. Space is more important than season when shaping soil microbial communities at a large spatial scale. Msystems 2020, 5, e00783-19. [Google Scholar] [CrossRef]
- Qi, D.; Wieneke, X.; Tao, J.; Zhou, X.; Desilva, U. Soil pH is the primary factor correlating with soil microbiome in karst rocky desertification regions in the Wushan County, Chongqing, China. Front. Microbiol. 2018, 9, 1027. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Barberán, A.; Gao, C.; Zhang, Z.; Wang, M.; Wurzburger, N.; Wang, X.; Zhang, R.; Li, J.; Zhang, J. Impact of urbanization on soil microbial diversity and composition in the megacity of Shanghai. Land Degrad. Dev. 2022, 33, 282–293. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrie, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef]
- Lu, C.; Kotze, D.J.; Setälä, H.M. Soil sealing causes substantial losses in C and N storage in urban soils under cool climate. Sci. Total Environ. 2020, 725, 138369. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, L.; Zhang, W.; Zhu, X.; Lei, M.; Niu, L. How did the bacterial community respond to the level of urbanization along the Yangtze River? Environ. Sci. Process. Impacts 2020, 22, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Elvidge, C.D.; Tuttle, B.T.; Sutton, P.S.; Baugh, K.E.; Howard, A.T.; Milesi, C.; Bhaduri, B.L.; Nemani, R. Global distribution and density of constructed impervious surfaces. Sensors 2007, 7, 1962–1979. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Dou, X.; Li, J.; Li, F. Impervious surfaces alter soil bacterial communities in urban areas: A case study in Beijing, China. Front. Microbiol. 2018, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Devi, N.B.; Yadava, P. Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India. Appl. Soil Ecol. 2006, 31, 220–227. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S. Tropical soil biology and fertility: A handbook of methods. Soil Sci. 1994, 157, 265. [Google Scholar] [CrossRef]
- Bremner, J.; Mulvaney, C. Total nitrogen. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Methods; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Sparling, G.; Whale, K.; Ramsay, A. Quantifying the contribution from the soil microbail biomass to the extractable P levels of fresh and air-dried soils. Soil Res. 1985, 23, 613–621. [Google Scholar] [CrossRef]
- Kalembasa, S.J.; Jenkinson, D.S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Luo, X.-S.; Xue, Y.; Wang, Y.-L.; Cang, L.; Xu, B.; Ding, J. Source identification and apportionment of heavy metals in urban soil profiles. Chemosphere 2015, 127, 152–157. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Szoecs, E. Vegan: Community Ecology Package R Package Version 2 2-1. 2015. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 15 February 2021).
- Eisenhauer, N.; Bowker, M.A.; Grace, J.B.; Powell, J.R. From patterns to causal understanding: Structural equation modeling (SEM) in soil ecology. Pedobiologia 2015, 58, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.-J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; De Cáceres, M. Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Grace, J.B. Structural equation modeling for observational studies. J. Wildl. Manag. 2008, 72, 14–22. [Google Scholar] [CrossRef]
- Barrico, L.; Castro, H.; Coutinho, A.P.; Gonçalves, M.T.; Freitas, H.; Castro, P. Plant and microbial biodiversity in urban forests and public gardens: Insights for cities’ sustainable development. Urban For. Urban Green. 2018, 29, 19–27. [Google Scholar] [CrossRef]
- Comte, J.; Del Giorgio, P.A. Linking the patterns of change in composition and function in bacterioplankton successions along environmental gradients. Ecology 2010, 91, 1466–1476. [Google Scholar] [CrossRef]
- Zhang, Y.; Cong, J.; Lu, H.; Yang, C.; Yang, Y.; Zhou, J.; Li, D. An integrated study to analyze soil microbial community structure and metabolic potential in two forest types. PLoS ONE 2014, 9, e93773. [Google Scholar] [CrossRef]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Gill, A.S.; Lee, A.; McGuire, K.L. Phylogenetic and functional diversity of total (DNA) and expressed (RNA) bacterial communities in urban green infrastructure bioswale soils. Appl. Environ. Microbiol. 2017, 83, e00287-17. [Google Scholar] [CrossRef]
- Gao, X.; Gu, Y.; Xie, T.; Zhen, G.; Huang, S.; Zhao, Y. Characterization and environmental risk assessment of heavy metals in construction and demolition wastes from five sources (chemical, metallurgical and light industries, and residential and recycled aggregates). Environ. Sci. Pollut. Res. 2015, 22, 9332–9344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Si, G.; Wang, J.; Luo, T.; Zhang, G. Bacterial community in alpine grasslands along an altitudinal gradient on the Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 87, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poeplau, C. Grassland soil organic carbon stocks along management intensity and warming gradients. Grass Forage Sci. 2021, 76, 186–195. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Sheng, R.; Moreira-Grez, B.; Liu, S.; Xu, R.; Li, K.; Wei, W. Influences of phosphorus and potassium deficiencies on the methanotrophic communities in rice rhizosphere. Appl. Soil Ecol. 2022, 170, 104265. [Google Scholar] [CrossRef]
- Mosier, A.C.; Francis, C.A. Relative abundance and diversity of ammonia--oxidizing archaea and bacteria in the San Francisco Bay estuary. Environ. Microbiol. 2008, 10, 3002–3016. [Google Scholar] [CrossRef]
- Wu, Y.; Song, Q.; Wu, J.; Zhou, J.; Zhou, L.; Wu, W. Field study on the soil bacterial associations to combined contamination with heavy metals and organic contaminants. Sci. Total Environ. 2021, 778, 146282. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Gao, X.; Huang, R.; Li, J.; Wang, C.; Lan, T.; Li, Q.; Deng, O.; Tao, Q.; Zeng, M. Temperature induces soil organic carbon mineralization in urban park green spaces, Chengdu, southwestern China: Effects of planting years and vegetation types. Urban For. Urban Green. 2020, 54, 126761. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forestland | Grassland | ||||||
|---|---|---|---|---|---|---|---|
| U1 | U2 | U3 | U1 | U2 | U3 | ||
| pH | / | 6.15 ± 0.28 ab | 6.67 ± 0.21 a | 6.75 ± 0.18 a | 6.54 ± 0.17 a | 6.79 ± 0.17 a | 6.85 ± 0.17 a |
| TOC | g/kg | 19.46 ± 1.0 a | 15.49 ± 0.94 b | 13.1 ± 1.13 bc | 14.71 ± 1.14 b | 14.58 ± 1.06 bc | 11.80 ± 0.71 c |
| TN | g/kg | 2.00 ± 0.15 a | 1.46 ± 0.12 bc | 1.44 ± 0.11 bc | 1.71 ± 0.16 ab | 1.29 ± 0.1 c | 1.18 ± 0.11 c |
| TP | g/kg | 0.83 ± 0.11 | 0.71 ± 0.08 | 0.86 ± 0.12 | 0.85 ± 0.06 | 0.75 ± 0.09 | 0.73 ± 0.08 |
| C/N | / | 10.36 ±0.58 ab | 11.62 ± 0.72 ab | 9.17 ± 0.63 b | 9.39 ± 0.96 ab | 12.04 ± 1.03 a | 11.03 ± 1.1 ab |
| N/P | / | 2.74 ± 0.19 a | 2.55 ± 0.39 a | 1.96 ± 0.22 ab | 2.17 ± 0.22 a | 1.93 ± 0.17 ab | 2.53 ± 0.98 a |
| SWC | % | 17.4 ± 0.94 ab | 16.3 ± 0.8 b | 16.3 ± 0.91 b | 18.94 ± 1.76 a | 17.22 ± 0.65 ab | 15.6 ± 0.53 b |
| SMBN | mg/kg | 14.82 ± 2.78 | 12.69 ± 2.06 | 10.94 ± 2.47 | 14.52 ± 2.15 | 12.86 ± 2.74 | 13.08 ± 2.72 |
| SMBP | mg/kg | 0.35 ± 0.08 | 0.49 ± 0.204 | 0.32 ± 0.13 | 0.31 ± 0.07 | 0.76 ± 0.33 | 0.31 ± 0.07 |
| As | mg/kg | 53.86 ± 0.83 | 55.34 ± 0.738 | 55.71 ± 1.05 | 54.14 ± 0.74 | 55.98 ± 0.95 | 54.61 ± 0.55 |
| Cd | mg/kg | 0.71 ± 0.01 b | 0.76 ± 0.02 ab | 0.88 ± 0.06 a | 0.74 ± 0.02 b | 0.76 ± 0.02 ab | 0.81 ± 0.03 ab |
| Cr | mg/kg | 243.84 ± 2.22 | 252.74 ± 2.87 | 247.12 ± 3.16 | 252.1 ± 3.92 | 254.26 ± 3.28 | 246.4 ± 2.6 |
| Cu | mg/kg | 98.3 ± 2.3 | 101.01 ± 1.78 | 99.79 ± 1.9 | 103.47 ± 2.78 | 103.51 ± 2.25 | 97.18 ± 1.45 |
| Mn | mg/kg | 425.14 ± 47.9 | 460.79 ± 32.73 | 480.27 ± 32.01 | 498.61 ± 39.96 | 494.69 ± 45.21 | 450.89 ± 37.63 |
| Ni | mg/kg | 106.69 ± 1.43 b | 112.15 ± 1.46 ab | 111.37 ± 1.69 ab | 110.78 ± 1.77 a | 112.8 ± 1.38 ab | 111.1 ± 2.19 ab |
| Pb | mg/kg | 117.12 ± 2.36 b | 120.79 ± 2.11 b | 135.18 ± 7.5 a | 119.31 ± 2.9 b | 120.4 ± 3.01 b | 119.22 ± 3.24 b |
| Zn | mg/kg | 348.2 ± 6.78 b | 361.26 ± 7.16 b | 396.87 ± 18.54 a | 365.68 ± 9.51 ab | 369.01 ± 9.88 ab | 370.42 ± 10.67 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, D.; Zhang, N.; Liu, S.; Ning, C.; Wang, X.; Feng, S. Urbanization Imprint on Soil Bacterial Communities in Forests and Grasslands. Forests 2023, 14, 38. https://doi.org/10.3390/f14010038
Gao D, Zhang N, Liu S, Ning C, Wang X, Feng S. Urbanization Imprint on Soil Bacterial Communities in Forests and Grasslands. Forests. 2023; 14(1):38. https://doi.org/10.3390/f14010038
Chicago/Turabian StyleGao, Dandan, Ning Zhang, Shuguang Liu, Chen Ning, Xinyue Wang, and Shuailong Feng. 2023. "Urbanization Imprint on Soil Bacterial Communities in Forests and Grasslands" Forests 14, no. 1: 38. https://doi.org/10.3390/f14010038
APA StyleGao, D., Zhang, N., Liu, S., Ning, C., Wang, X., & Feng, S. (2023). Urbanization Imprint on Soil Bacterial Communities in Forests and Grasslands. Forests, 14(1), 38. https://doi.org/10.3390/f14010038