


Anthropogenic Dusts Influence Leaf Anatomical and Eco-Physiological Traits of Black Locust (Robinia pseudoacacia L.) Growing on Vesuvius Volcano

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Species Selection
2.2. Dust Deposition Quantification
2.3. Leaf functional Traits and Leaf Anatomical Traits
2.4. Chlorophyll a Fluorescence Analysis and Photosynthetic Pigment Content
2.5. Statistical Analyses
3. Results
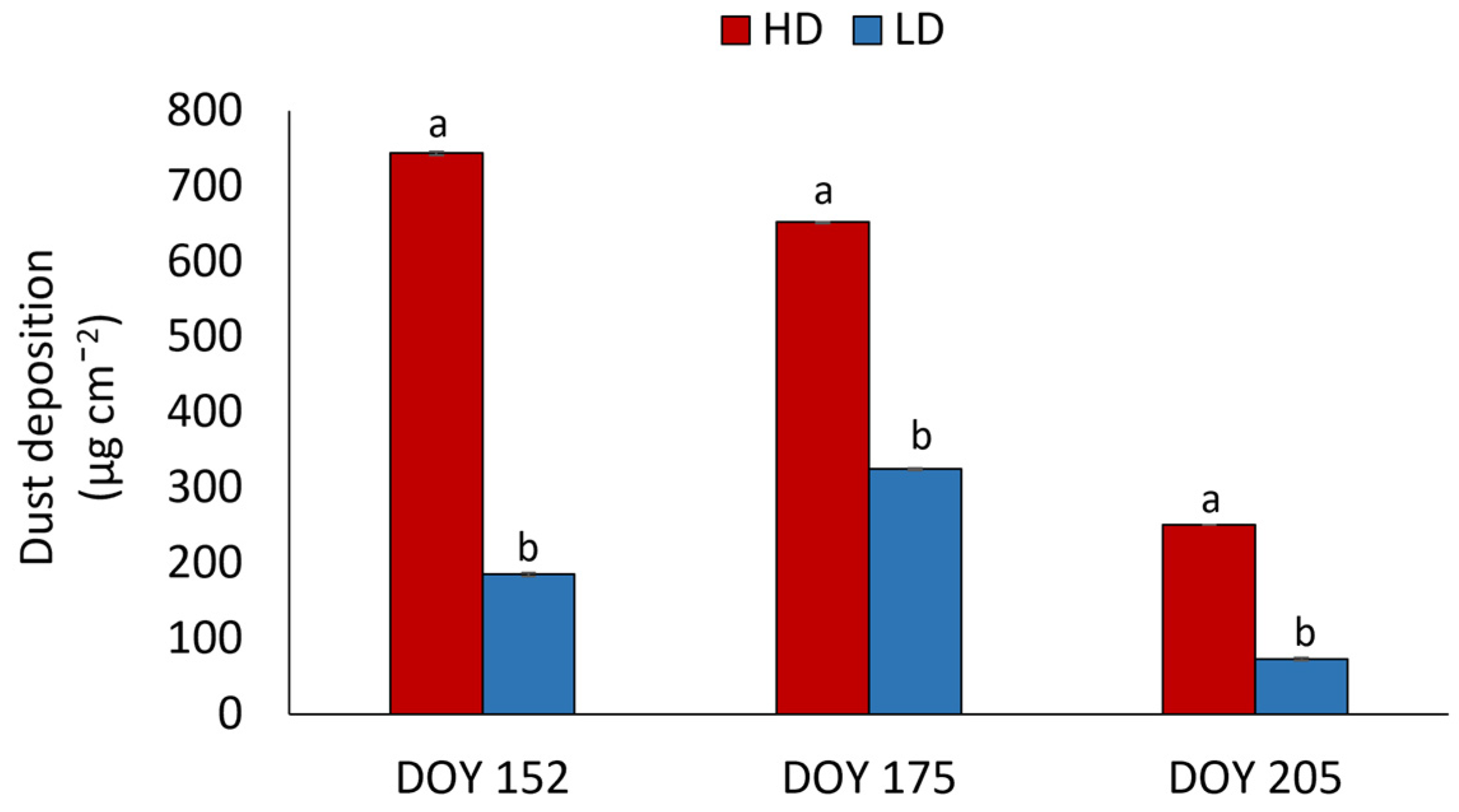
3.1. Dust Accumulation on Leaf Lamina
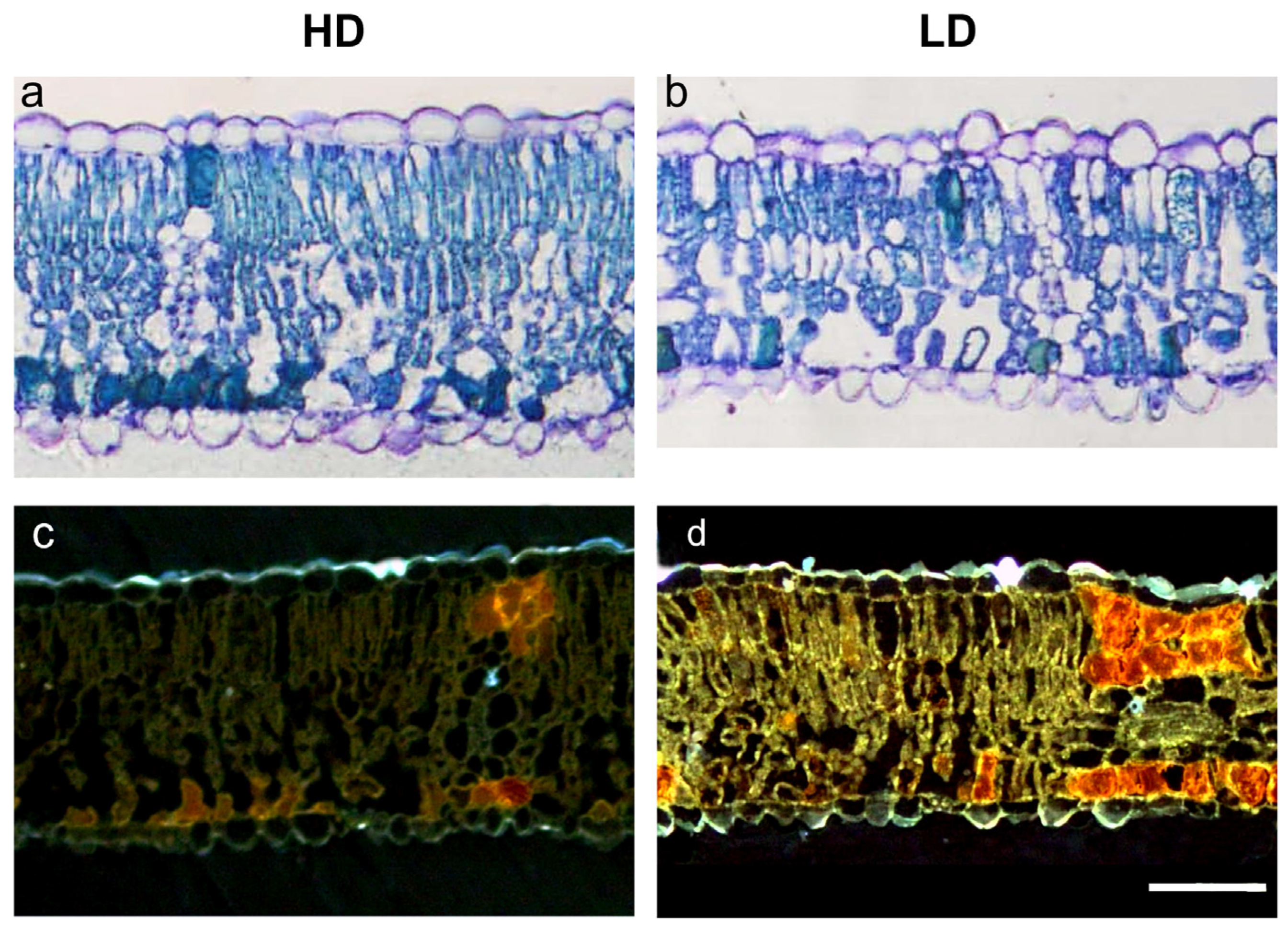
3.2. Leaf Functional Traits and Leaf Anatomical Traits
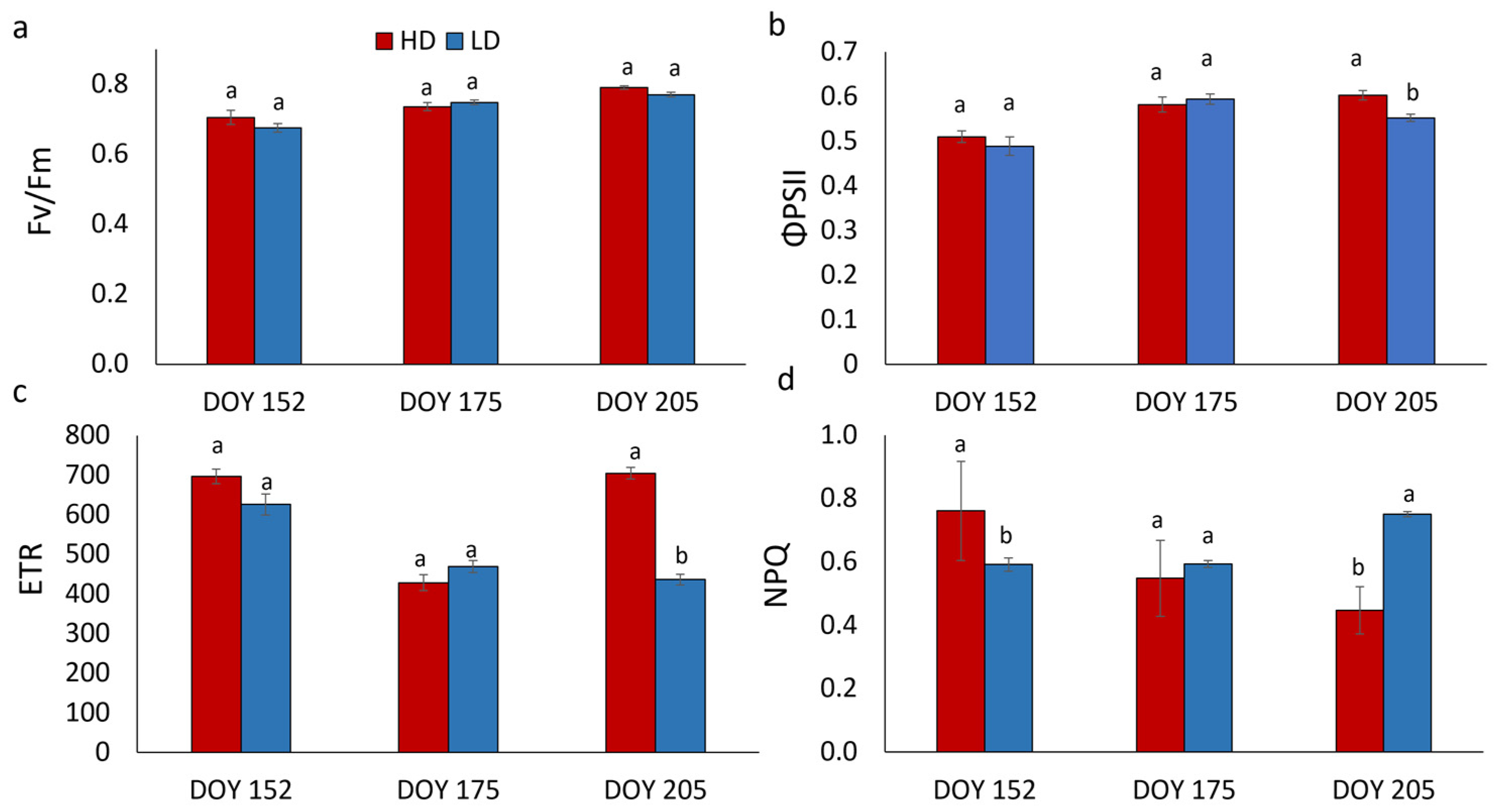
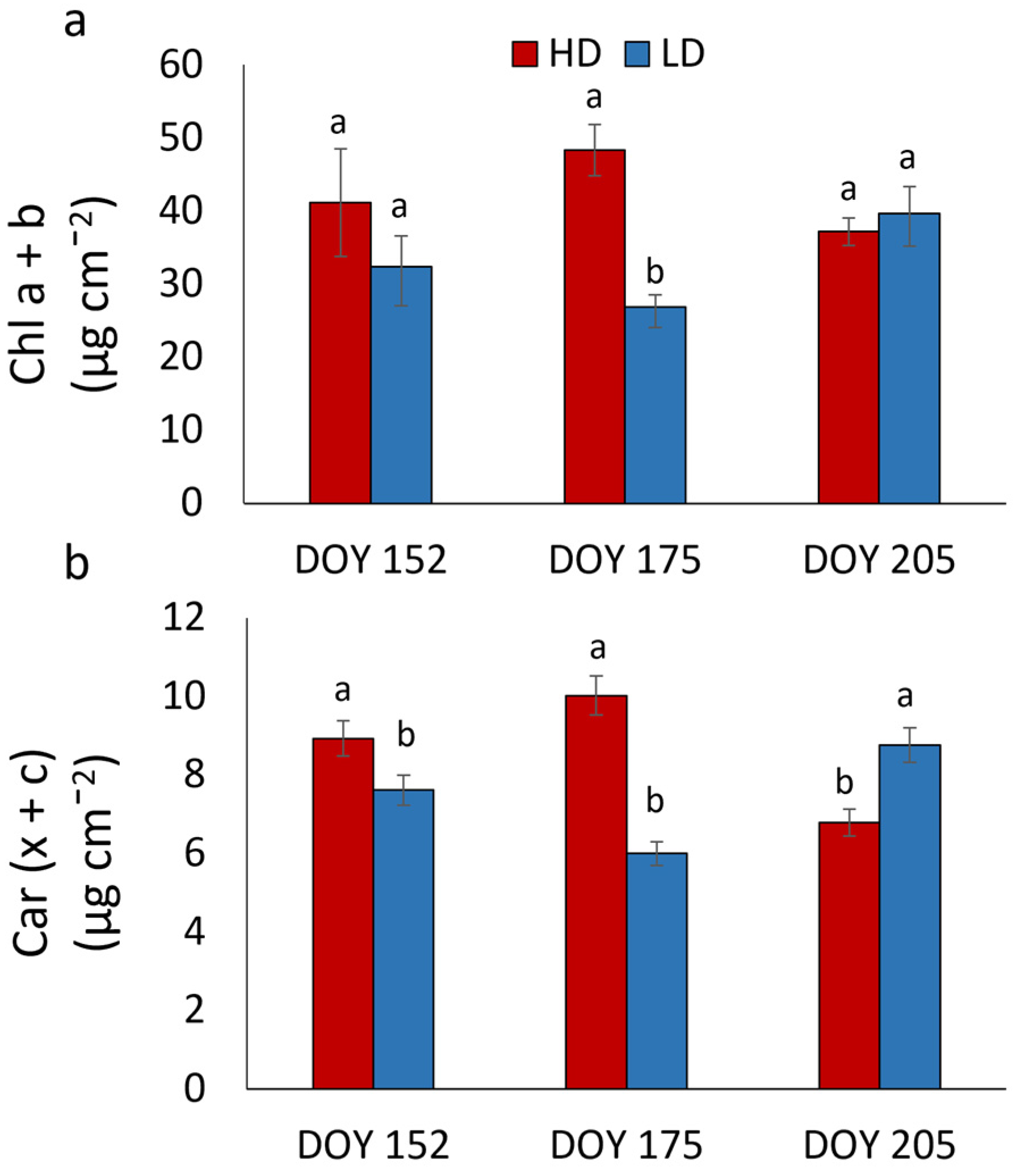
3.3. Chlorophyll a Fluorescence Analysis and Photosynthetic Pigment Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentili, R.; Ferrè, C.; Cardarelli, E.; Montagnani, C.; Bogliani, G.; Citterio, S.; Comolli, R. Comparing negative impacts of Prunus serotina, Quercus rubra and Robinia pseudoacacia on native forest ecosystems. Forests 2019, 10, 842. [Google Scholar] [CrossRef] [Green Version]
- Sukopp, H.; Wurzel, A. The effects of climate change on the vegetation of central European cities. Urban Habitats 2003, 1, 66–86. [Google Scholar]
- Chmielewski, F.M.; Müller, A.; Küchler, W. Possible impacts of climate change on natural vegetation in Saxony (Germany). Int. J. Biometeorol. 2005, 50, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Kleinbauer, I.; Dullinger, S.; Peterseil, J.; Essl, F. Climate change might drive the invasive tree Robinia pseudacacia into nature reserves and endangered habitats. Biol. Conserv. 2010, 143, 382–390. [Google Scholar] [CrossRef]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, M.; Capozzi, F.; Amitrano, C.; Giordano, S.; Arena, C.; Spagnuolo, V. Performance of three cardoon cultivars in an industrial heavy metal-contaminated soil: Effects on morphology, cytology and photosynthesis. J. Hazard. Mater. 2018, 351, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, A.; Conti, G.O.; Jho, E.H.; Zuccarello, P.; Grasso, A.; Copat, C.; Ferrante, M. Phytoremediation of contaminated soils by heavy metals and PAHs. A brief review. Environ. Technol. Innov. 2017, 8, 309–326. [Google Scholar] [CrossRef]
- Fatema, K.; Shoily, S.S.; Ahsan, T.; Haidar, Z.; Sumit, A.F.; Sajib, A.A. Effects of arsenic and heavy metals on metabolic pathways in cells of human origin: Similarities and differences. Toxicol. Rep. 2021, 8, 1109–1120. [Google Scholar] [CrossRef]
- Woś, B.; Pająk, M.; Krzaklewski, W.; Pietrzykowski, M. Verifying the utility of black locust (Robinia pseudoacacia L.) in the reclamation of a lignite combustion waste disposal site in Central European conditions. Forests 2020, 11, 877. [Google Scholar] [CrossRef]
- Środek, D.; Rahmonov, O. The properties of Black Locust Robinia pseudoacacia L. to selectively accumulate chemical elements from soils of ecologically transformed areas. Forests 2021, 13, 7. [Google Scholar] [CrossRef]
- Nicolescu, V.-N.; Rédei, K.; Mason, W.L.; Vor, T.; Pöetzelsberger, E.; Bastien, J.-C.; Brus, R.; Benčať, T.; Đodan, M.; Cvjetkovic, B. Ecology, growth and management of black locust (Robinia pseudoacacia L.), a non-native species integrated into European forests. J. For. Res. 2020, 31, 1081–1101. [Google Scholar] [CrossRef]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. Structure and ecosystem services of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.) in urban environments. Urban For. Urban Green. 2015, 14, 1110–1121. [Google Scholar] [CrossRef]
- Esfandiary, M.; Sodaiezadeh, H.; Hakimzadeh, M.A. Assessment some of heavy metals in black locust (Robinia pseudoascacia) in the Yazd highway green belt. J. Ornam. Plants 2019, 9, 193–203. [Google Scholar]
- Rai, P.K. Biodiversity of roadside plants and their response to air pollution in an Indo-Burma hotspot region: Implications for urban ecosystem restoration. J. Asia- Pac. Biodivers. 2016, 9, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, I.J.; Rathore, D. Dust pollution: Its removal and effect on foliage physiology of urban trees. Sustain. Cities Soc. 2019, 51, 101696. [Google Scholar] [CrossRef]
- Medlyn, B.E. Physiological basis of the light use efficiency model. Tree Physiol. 1998, 18, 167–176. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Arena, C.; Vitale, L.; Aronne, G.; Virzo De Santo, A. Anatomy and photochemical behaviour of Mediterranean Cistus incanus winter leaves under natural outdoor and warmer indoor conditions. Botany 2011, 89, 677–688. [Google Scholar] [CrossRef]
- Vitale, L.; Arena, C.; De Santo, A.V. Seasonal changes in photosynthetic activity and photochemical efficiency of the Mediterranean shrub Phillyrea angustifolia L. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2012, 146, 443–450. [Google Scholar]
- Arena, C.; Mistretta, C.; Di Natale, E.; Mennella, M.R.F.; De Santo, A.V.; De Maio, A. Characterization and role of poly (ADP-ribosyl) ation in the Mediterranean species Cistus incanus L. under different temperature conditions. Plant Physiol. Biochem. 2011, 49, 435–440. [Google Scholar] [CrossRef]
- Vitale, L.; Magliulo, V.; Arena, C. Morphological and physiological modifications of Cistus salvifolius L. winter leaves in response to the rise in winter temperatures. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 1093–1101. [Google Scholar]
- Gonsamo, A.; Chen, J.M. 3.11—Vegetation Primary Productivity. In Comprehensive Remote Sensing; Liang, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 3, pp. 163–189. [Google Scholar]
- Osmond, B.; Förster, B. Photoinhibition: Then and now. In Photoprotection, photoinhibition, gene regulation, and environment; Springer: Berlin/Heidelberg, Germany, 2008; pp. 11–22. [Google Scholar]
- Oguchi, R.; Onoda, Y.; Terashima, I.; Tholen, D. Leaf Anatomy and Function. In The Leaf: A Platform for Performing Photosynthesis; Springer: Berlin/Heidelberg, Germany, 2018; pp. 97–139. [Google Scholar]
- Calzavara, A.; Rocha, J.; Lourenço, G.; Sanada, K.; Medri, C.; Bianchini, E.; Pimenta, J.; Stolf-Moreira, R.; Oliveira, H. Acclimation responses to high light by Guazuma ulmifolia Lam.(Malvaceae) leaves at different stages of development. Plant Biol. 2017, 19, 720–727. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; De Santo, A.V. Paraheliotropism in Robinia pseudoacacia L.: An efficient strategy to optimise photosynthetic performance under natural environmental conditions. Plant Biol. 2008, 10, 194–201. [Google Scholar] [CrossRef] [PubMed]
- De Marco, A.; Arena, C.; Giordano, M.; Virzo De Santo, A. Impact of the invasive tree black locust on soil properties of Mediterranean stone pine-holm oak forests. Plant. Soil 2013, 372, 473–486. [Google Scholar] [CrossRef]
- De Micco, V.; Amitrano, C.; Stinca, A.; Izzo, L.; Zalloni, E.; Balzano, A.; Barile, R.; Conti, P.; Arena, C. Dust accumulation due to anthropogenic impact induces anatomical and photochemical changes in leaves of Centranthus ruber growing on the slope of the Vesuvius volcano. Plant Biol. 2020, 22, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Motti, R.; Stinca, A.; Ricciardi, M. Flora e Vegetazione. Laboratorio per il monitoraggio della biodiversità e cartografia del Parco Nazionale del Vesuvio. Napoli. Vesuvio; Carpino, F., Sammichelu, F., Eds.; Ente Parco Nazionale del Vesuvio: Napoli, Italy, 2009; pp. 17–64. [Google Scholar]
- Memoli, V.; Eymar, E.; García-Delgado, C.; Esposito, F.; Santorufo, L.; De Marco, A.; Barile, R.; Maisto, G. Total and fraction content of elements in volcanic soil: Natural or anthropogenic derivation. Sci. Total Environ. 2018, 625, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Memoli, V.; Eymar, E.; García-Delgado, C.; Esposito, F.; Panico, S.C.; De Marco, A.; Barile, R.; Maisto, G. Soil element fractions affect phytotoxicity, microbial biomass and activity in volcanic areas. Sci. Total Environ. 2018, 636, 1099–1108. [Google Scholar] [CrossRef]
- Balzano, A.; Čufar, K.; Battipaglia, G.; Merela, M.; Prislan, P.; Aronne, G.; De Micco, V. Xylogenesis reveals the genesis and ecological signal of IADFs in Pinus pinea L. and Arbutus unedo L. Ann. Bot. 2018, 121, 1231–1242. [Google Scholar] [CrossRef]
- Montecchiari, S. Syntaxonomical and Ecological Characterization of Two Main Alien Forest Communities: Robinia Pseudoacacia and Ailanthus Altissima at Their Southern Limit in Europe. Ph.D. Thesis, Polytechnic University of Marche, Ancona, Italy, 2021. [Google Scholar]
- Capozzi, F.; Di Palma, A.; Sorrentino, M.C.; Adamo, P.; Giordano, S.; Spagnuolo, V. Morphological Traits Influence the Uptake Ability of Priority Pollutant Elements by Hypnum cupressiforme and Robinia pseudoacacia Leaves. Atmosphere 2020, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Younis, U.; Bokhari, T.Z.; Malik, S.A.; Ahmad, S.; Raja, R. Variations in leaf dust accumulation, foliage and pigment attributes in fruiting plant species exposed to particulate pollution from Multan. Int. J. Agric. Sci. Res. 2013, 3, 1–12. [Google Scholar]
- Cornelissen, J.; Cerabolini, B.; Castro-Díez, P.; Villar-Salvador, P.; Montserrat-Martí, G.; Puyravaud, J.; Maestro, M.; Werger, M.; Aerts, R. Functional traits of woody plants: Correspondence of species rankings between field adults and laboratory-grown seedlings? J. Veg. Sci. 2003, 14, 311–322. [Google Scholar] [CrossRef]
- Feder, N.; O’brien, T. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Fujii, T. Peroxide dependent and independent lipid peroxidation: Site-specific mechanisms of initiation by chelated iron and inhibition by α-tocopherol. Lipids 1992, 27, 227. [Google Scholar] [CrossRef] [PubMed]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999; Volume 198. [Google Scholar]
- Kolb, C.A.; Kaser, M.A.; Kopecký, J.; Zotz, G.; Riederer, M.; Pfundel, E.E. Effects of natural intensities of visible and ultraviolet radiation on epidermal ultraviolet screening and photosynthesis in grape leaves. Plant Physiol. 2001, 127, 863–875. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Arena, C.; Aronne, G. Anatomical alterations of Phaseolus vulgaris L. mature leaves irradiated with X-rays. Plant Biol. 2014, 16, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Conti, S.; Francesca, S.; Melchionna, G.; Hájek, J.; Barták, M.; Barone, A.; Rigano, M.M. Eco-physiological screening of different tomato genotypes in response to high temperatures: A combined field-to-laboratory approach. Plants 2020, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Butler, W. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. Biochim. Biophys. Acta (BBA)-Bioenerg. 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Krall, J.P.; Edwards, G.E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods Enzymol; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- De Micco, V.; Aronne, G. Morpho-Anatomical Traits for Plant Adaptation to Drought. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.; Carins Murphy, M.R. Xylem and stomata, coordinated through time and space. Plant Cell Environ. 2017, 40, 872–880. [Google Scholar] [CrossRef]
- Amitrano, C.; Arena, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Vapour pressure deficit: The hidden driver behind plant morphofunctional traits in controlled environments. Ann. Appl. Biol. 2019, 175, 313–325. [Google Scholar] [CrossRef]
- Amitrano, C.; Junker, A.; D’Agostino, N.; De Pascale, S.; De Micco, V. Integration of high-throughput phenotyping with anatomical traits of leaves to help understanding lettuce acclimation to a changing environment. Planta 2022, 256, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Pepe, M.; Crescente, M.F.; Varone, L. Effect of Water Stress on Physiological and Morphological Leaf Traits: A Comparison among the Three Widely-Spread Invasive Alien Species Ailanthus altissima, Phytolacca americana, and Robinia pseudoacacia. Plants 2022, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Leishman, M.R.; Haslehurst, T.; Ares, A.; Baruch, Z. Leaf trait relationships of native and invasive plants: Community-and global-scale comparisons. New Phytol. 2007, 176, 635–643. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, A.; Sorrell, B.K.; Eller, F. Carbon assimilation through a vertical light gradient in the canopy of invasive herbs grown under different temperature regimes is determined by leaf and whole-plant architecture. AoB Plants 2020, 12, plaa031. [Google Scholar] [CrossRef]
- Gallé, A.; Haldimann, P.; Feller, U. Photosynthetic performance and water relations in young pubescent oak (Quercus pubescens) trees during drought stress and recovery. New Phytol. 2007, 174, 799–810. [Google Scholar] [CrossRef]
- Jiang, Y.; Feng, X.; Wang, H.; Chen, Y.; Sun, Y. Heat-induced down-regulation of photosystem II protects photosystem I in honeysuckle (Lonicera japonica). J. Plant Res. 2021, 134, 1311–1321. [Google Scholar] [CrossRef]
- Zhang, R.; Sharkey, T.D. Photosynthetic electron transport and proton flux under moderate heat stress. Photosynth. Res. 2009, 100, 29–43. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lehmeier, C.; Pajor, R.; Lundgren, M.R.; Mathers, A.; Sloan, J.; Bauch, M.; Mitchell, A.; Bellasio, C.; Green, A.; Bouyer, D. Cell density and airspace patterning in the leaf can be manipulated to increase leaf photosynthetic capacity. Plant J. 2017, 92, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Way, D.A.; Pearcy, R.W. Sunflecks in trees and forests: From photosynthetic physiology to global change biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.C. The physiological significance of variation in leaf structure. Sci. Prog. 1972, 60, 25–51. [Google Scholar]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic efficiency in sun and shade plants. Photosynthetica 2018, 56, 354–365. [Google Scholar] [CrossRef]
- Du, Q.; Liu, T.; Jiao, X.; Song, X.; Zhang, J.; Li, J. Leaf anatomical adaptations have central roles in photosynthetic acclimation to humidity. J. Exp. Bot. 2019, 70, 4949–4962. [Google Scholar] [CrossRef]
- Cruces, E.; Flores-Molina, M.R.; Díaz, M.J.; Huovinen, P.; Gómez, I. Phenolics as photoprotective mechanism against combined action of UV radiation and temperature in the red alga Gracilaria chilensis? J. Appl. Phycol. 2018, 30, 1247–1257. [Google Scholar] [CrossRef]
- Valladares, F.; Pearcy, R. Drought can be more critical in the shade than in the sun: A field study of carbon gain and photo-inhibition in a Californian shrub during a dry El Niño year. Plant Cell Environ. 2002, 25, 749–759. [Google Scholar] [CrossRef]
- Hall, F.G.; Hilker, T.; Coops, N.C.; Lyapustin, A.; Huemmrich, K.F.; Middleton, E.; Margolis, H.; Drolet, G.; Black, T.A. Multi-angle remote sensing of forest light use efficiency by observing PRI variation with canopy shadow fraction. Remote Sens. Environ. 2008, 112, 3201–3211. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Douthe, C.; Flexas, J. Differential coordination of stomatal conductance, mesophyll conductance, and leaf hydraulic conductance in response to changing light across species. Plant Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Amitrano, C.; Arena, C.; Cirillo, V.; De Pascale, S.; De Micco, V. Leaf morpho-anatomical traits in Vigna radiata L. affect plant photosynthetic acclimation to changing vapor pressure deficit. Environ. Exp. Bot. 2021, 186, 104453. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DOY 152 | DOY 175 | DOY 205 | ||
|---|---|---|---|---|
| LA (cm2) | HD | 7.38 ± 0.75a | 6.26 ± 0.46a | 6.32 ± 0.22a |
| LD | 5.04 ± 0.30b | 5.31 ± 0.41a | 6.62 ± 0.25a | |
| SLA (cm2 g−1) | HD | 215.45 ± 5.03a | 180.97 ± 4.03a | 162.95 ± 2.06b |
| LD | 226.59 ± 5.09a | 156.46 ± 6.32b | 178.39 ± 4.80a | |
| LDMC (g g−1) | HD | 0.28 ± 0.01a | 0.32 ± 0.01a | 0.34 ± 0.01a |
| LD | 0.27 ± 0.01b | 0.28 ± 0.02b | 0.35 ± 0.02a | |
| RWC (%) | HD | 86.52 ± 0.93a | 82.78 ± 0.81a | 84.85 ± 0.80a |
| LD | 86.04 ± 1.31a | 73.51 ± 1.94b | 78.64 ± 1.35b |
| HD | LD | |
|---|---|---|
| PPT (µm) | 57.52 ± 1.93a | 56.44 ± 1.65a |
| SPT (µm) | 62.54 ± 1.83a | 60.94 ± 1.72a |
| TPT (µm) | 157.57 ± 3.07a | 145.28 ± 2.19b |
| IS (%) | 24.44 ± 2.49b | 34.77 ± 2.94a |
| PC (%) | 29.66 ± 0.23b | 41.46 ± 0.22a |
| PCi | 5.93 ± 0.16b | 10.68 ± 0.07a |
| SF (n mm−1) | 3.04 ± 1.31a | 3.53 ± 1.55a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Micco, V.; Amitrano, C.; Balzano, A.; Cirillo, C.; Izzo, L.G.; Vitale, E.; Arena, C. Anthropogenic Dusts Influence Leaf Anatomical and Eco-Physiological Traits of Black Locust (Robinia pseudoacacia L.) Growing on Vesuvius Volcano. Forests 2023, 14, 212. https://doi.org/10.3390/f14020212
De Micco V, Amitrano C, Balzano A, Cirillo C, Izzo LG, Vitale E, Arena C. Anthropogenic Dusts Influence Leaf Anatomical and Eco-Physiological Traits of Black Locust (Robinia pseudoacacia L.) Growing on Vesuvius Volcano. Forests. 2023; 14(2):212. https://doi.org/10.3390/f14020212
Chicago/Turabian StyleDe Micco, Veronica, Chiara Amitrano, Angela Balzano, Chiara Cirillo, Luigi Gennaro Izzo, Ermenegilda Vitale, and Carmen Arena. 2023. "Anthropogenic Dusts Influence Leaf Anatomical and Eco-Physiological Traits of Black Locust (Robinia pseudoacacia L.) Growing on Vesuvius Volcano" Forests 14, no. 2: 212. https://doi.org/10.3390/f14020212
APA StyleDe Micco, V., Amitrano, C., Balzano, A., Cirillo, C., Izzo, L. G., Vitale, E., & Arena, C. (2023). Anthropogenic Dusts Influence Leaf Anatomical and Eco-Physiological Traits of Black Locust (Robinia pseudoacacia L.) Growing on Vesuvius Volcano. Forests, 14(2), 212. https://doi.org/10.3390/f14020212