
Habitat Ecology, Structure Influence Diversity, and Host-Species Associations of Wild Orchids in Undisturbed and Disturbed Forests in Peninsular Malaysia

Abstract
:1. Introduction
2. Materials and Methods
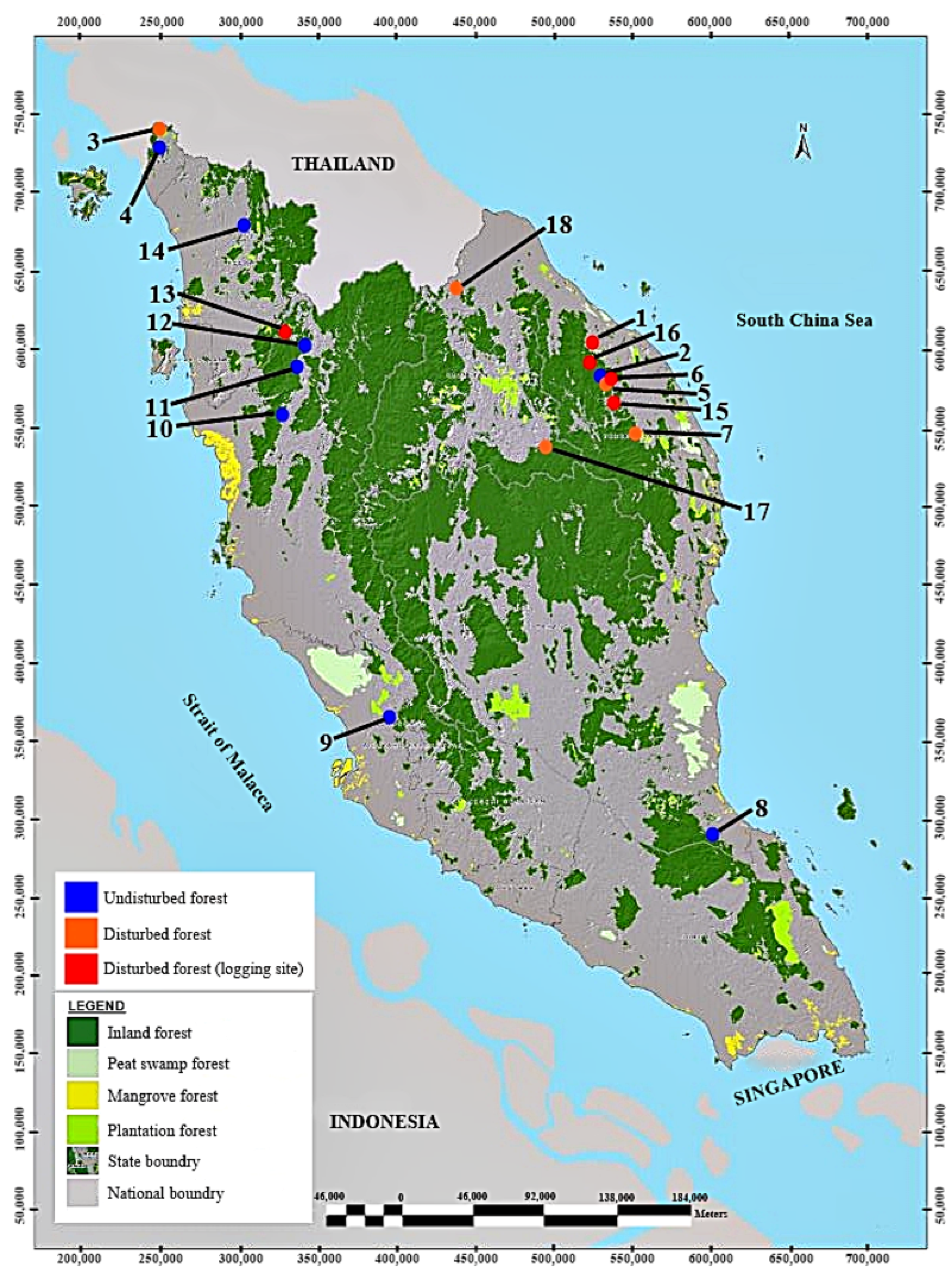
2.1. Study Area
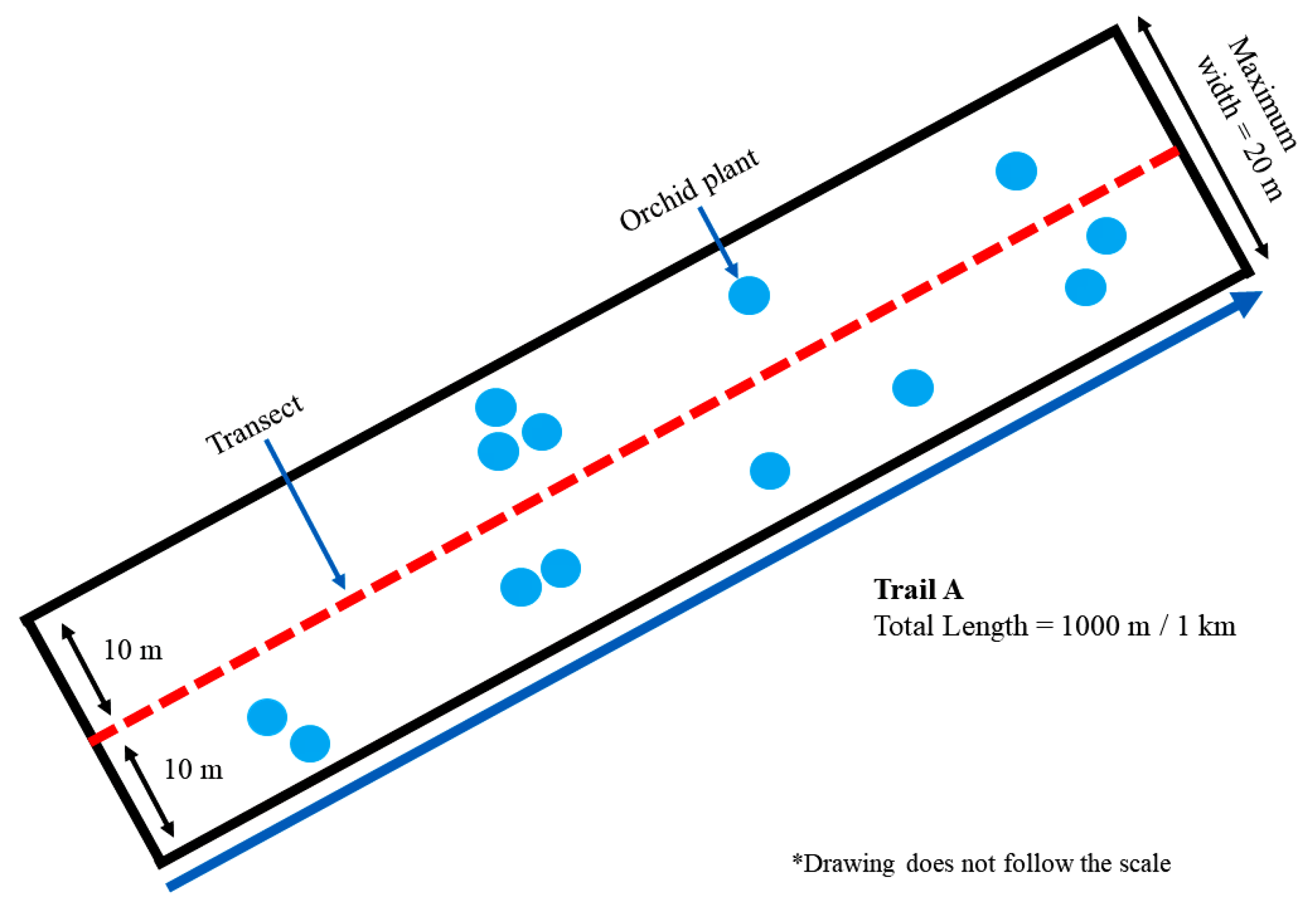
2.2. Sampling Design, Samples, and Data Collection
2.3. Ecological Study Design and In Situ Measurement of Microclimate Data
2.4. Diversity Analyses
2.5. Data Analyses
2.6. Statistical Analyses
2.7. Terminology
3. Results
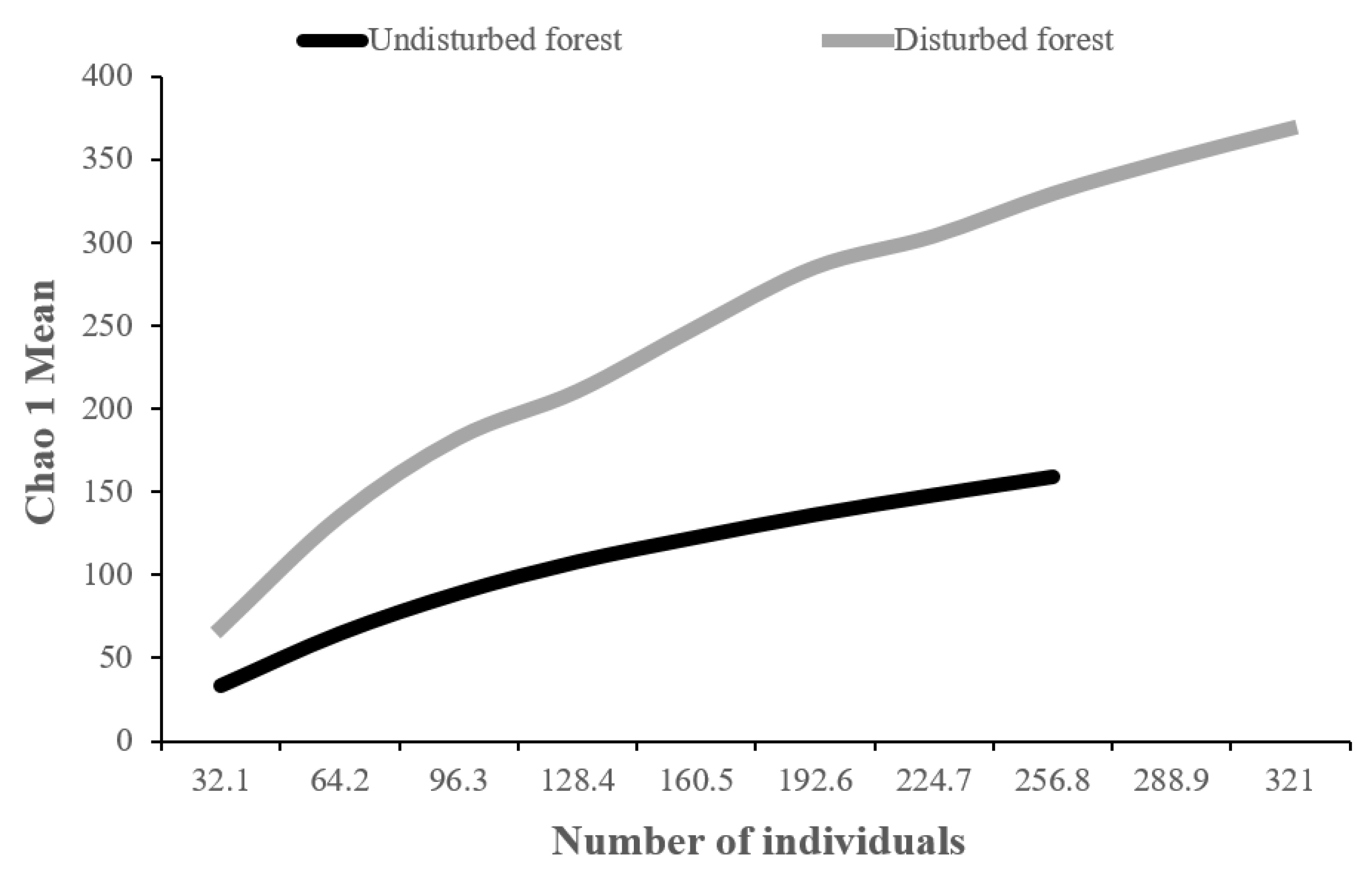
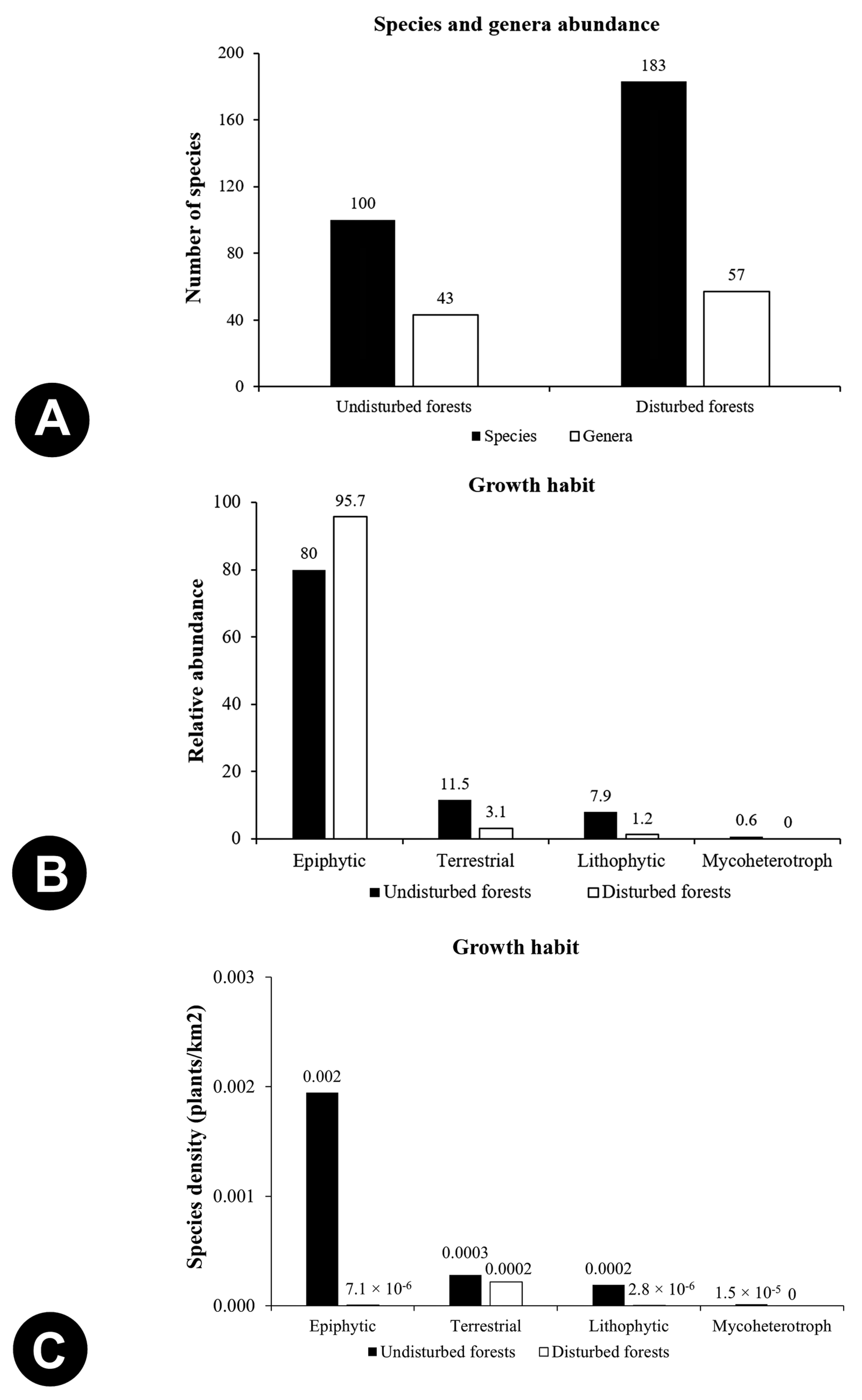
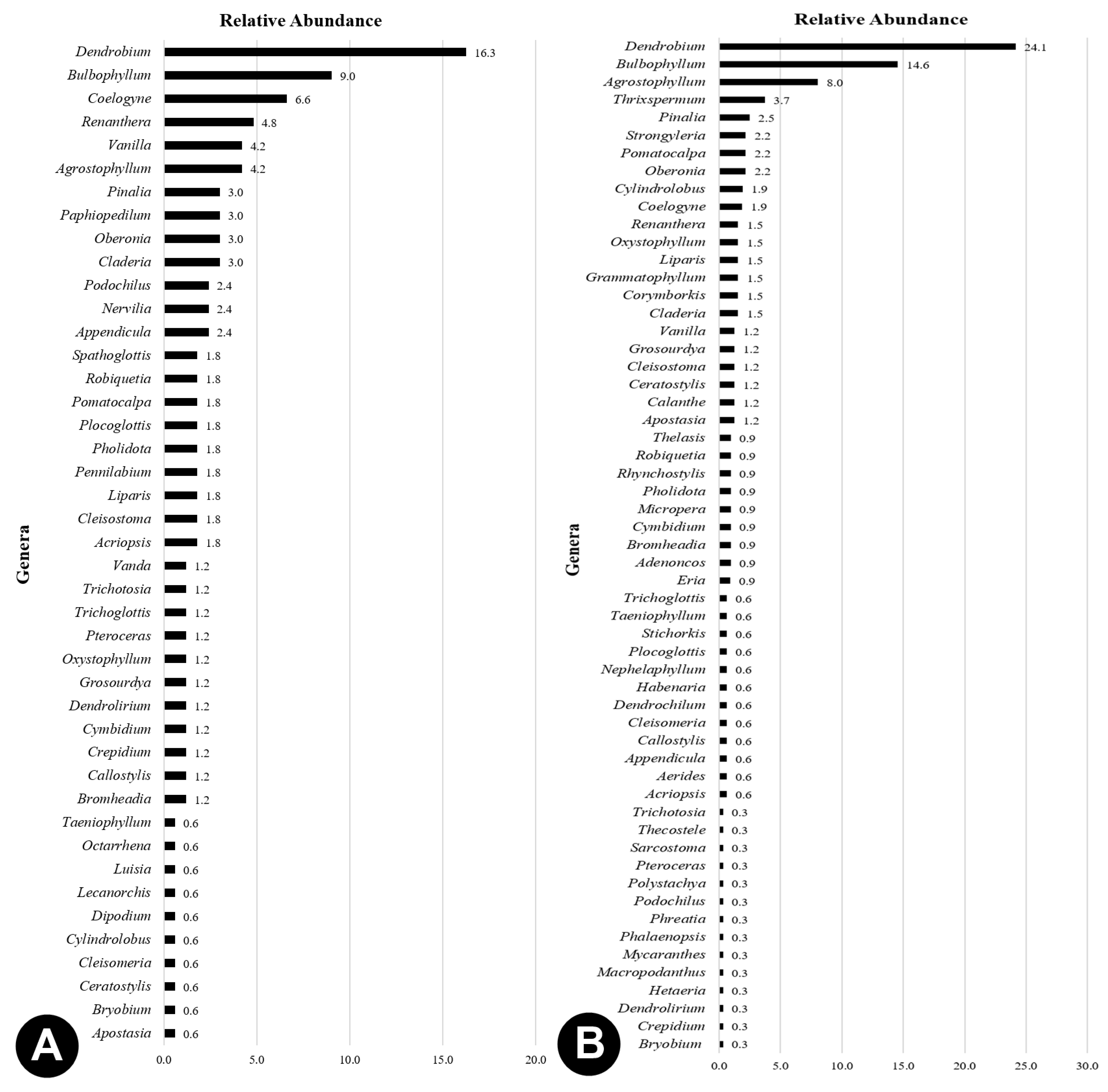
3.1. Species Richness and Abundance
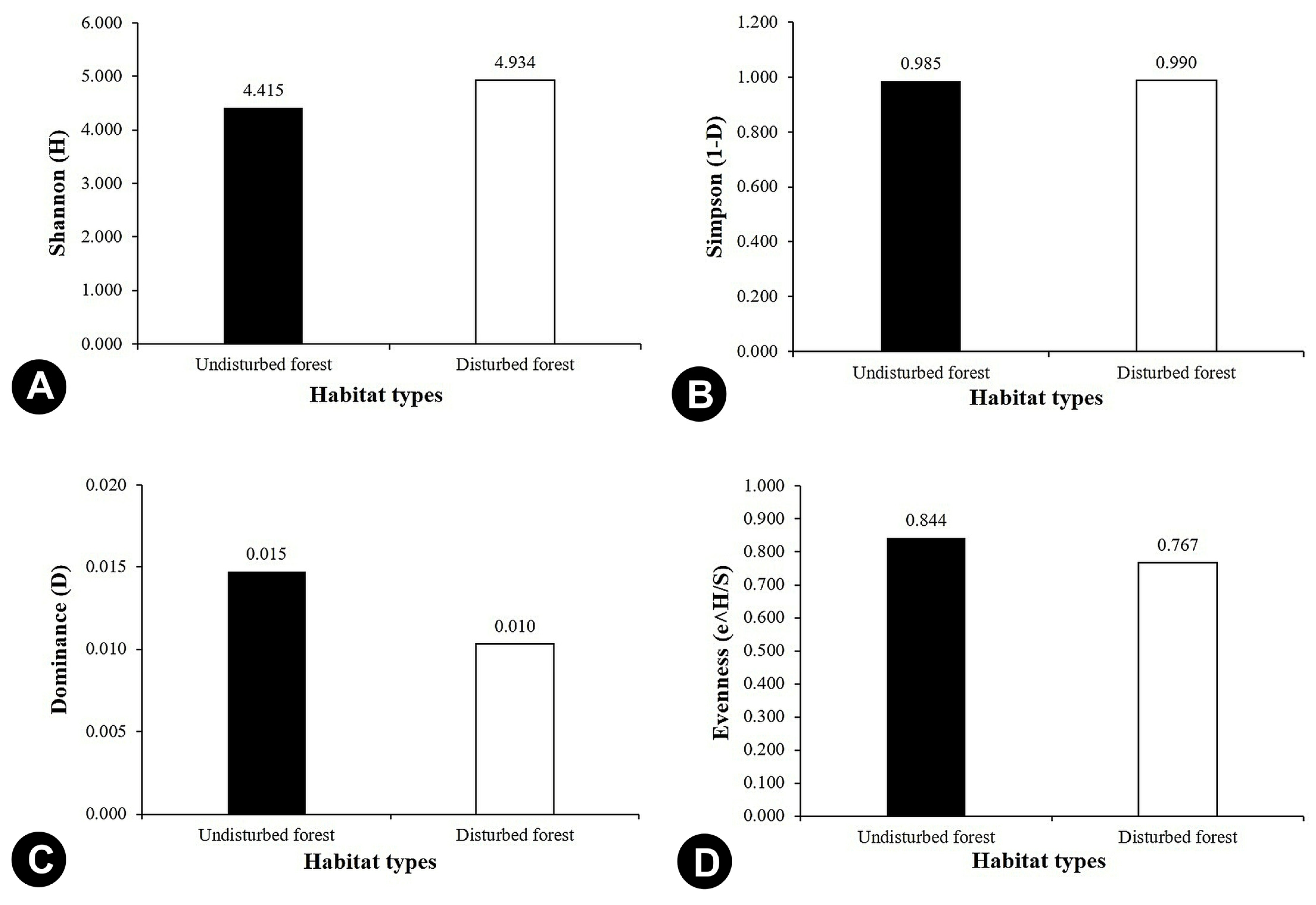
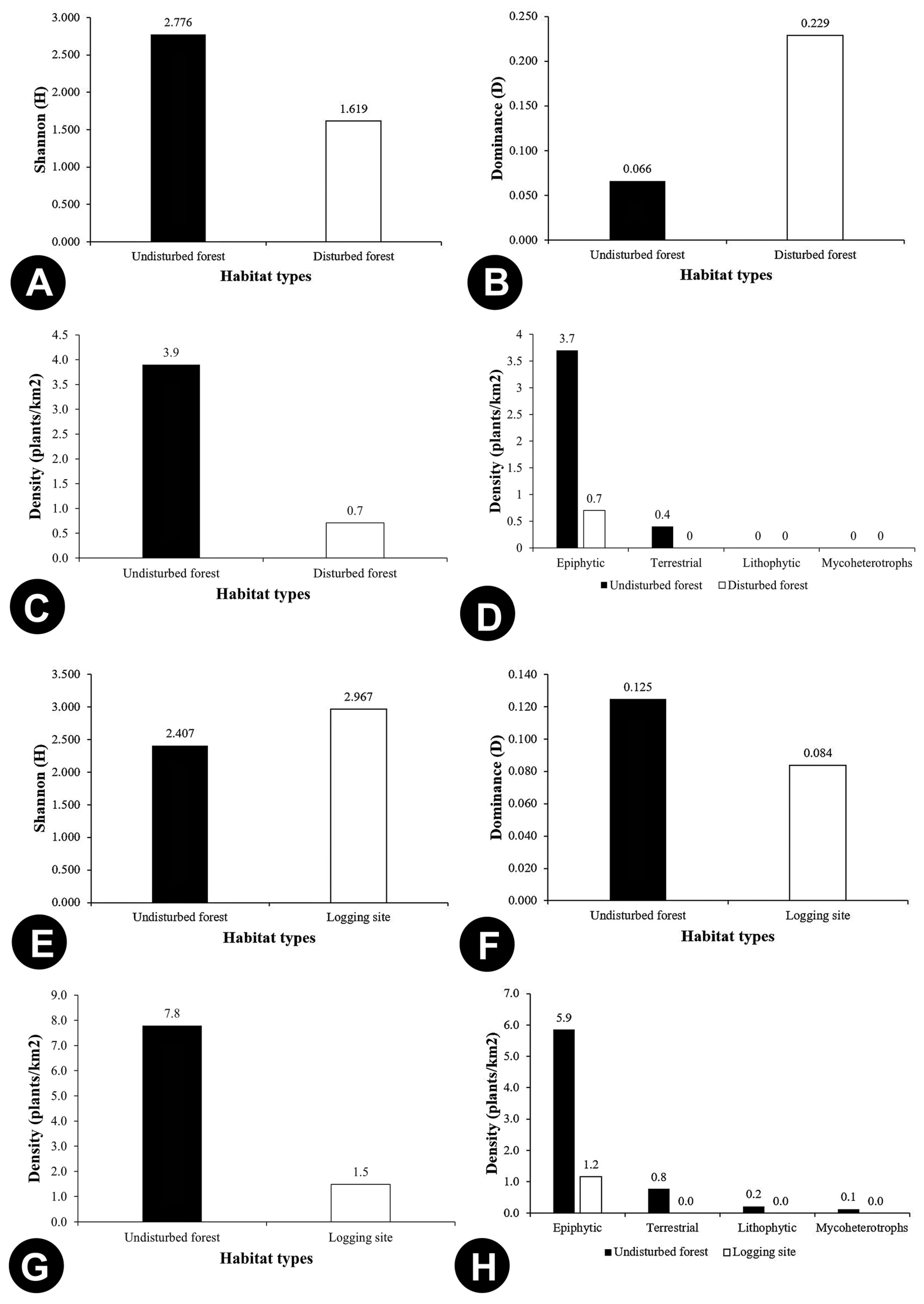
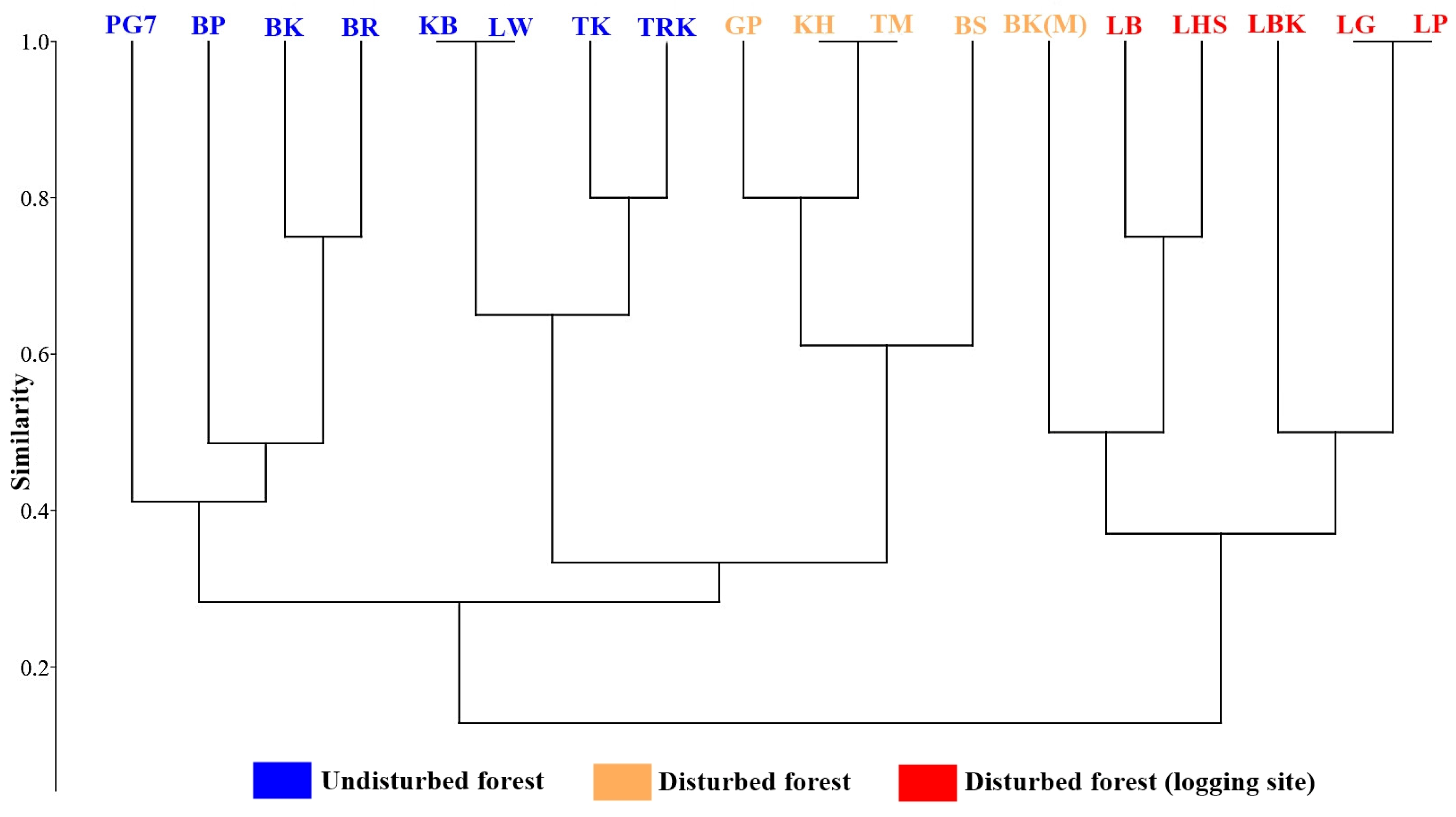
3.2. Species Diversity
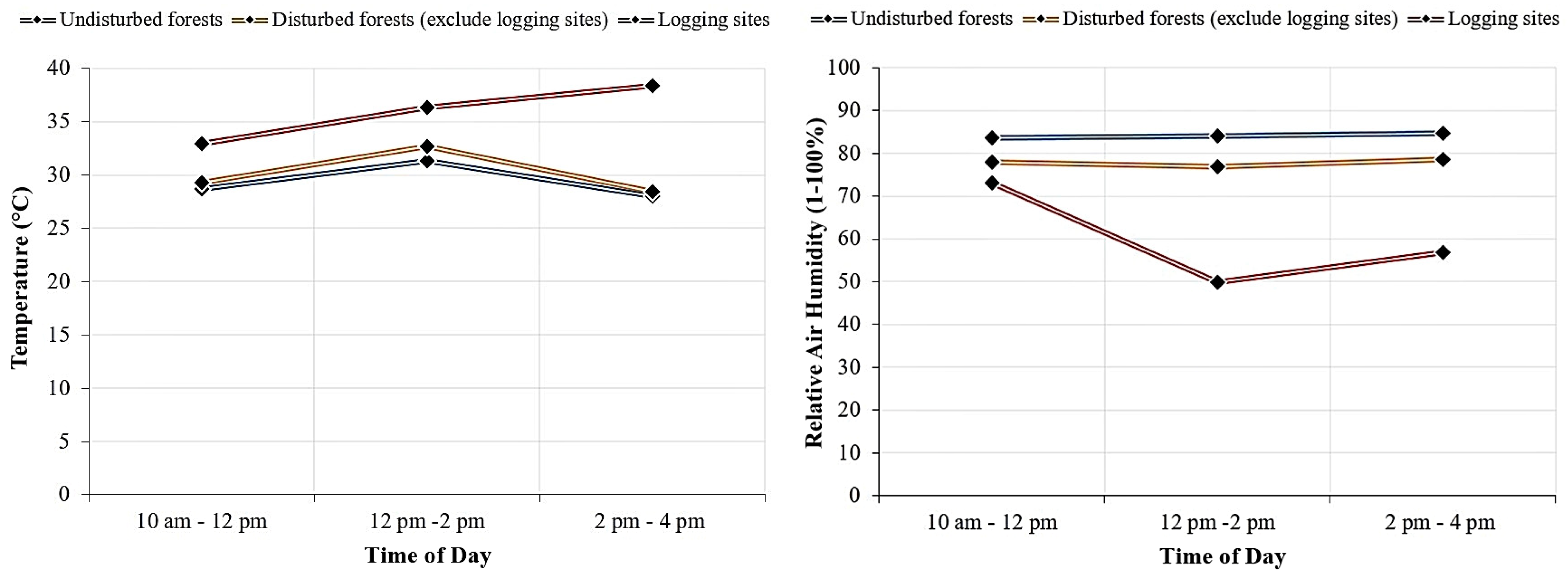
3.3. Microclimate Conditions
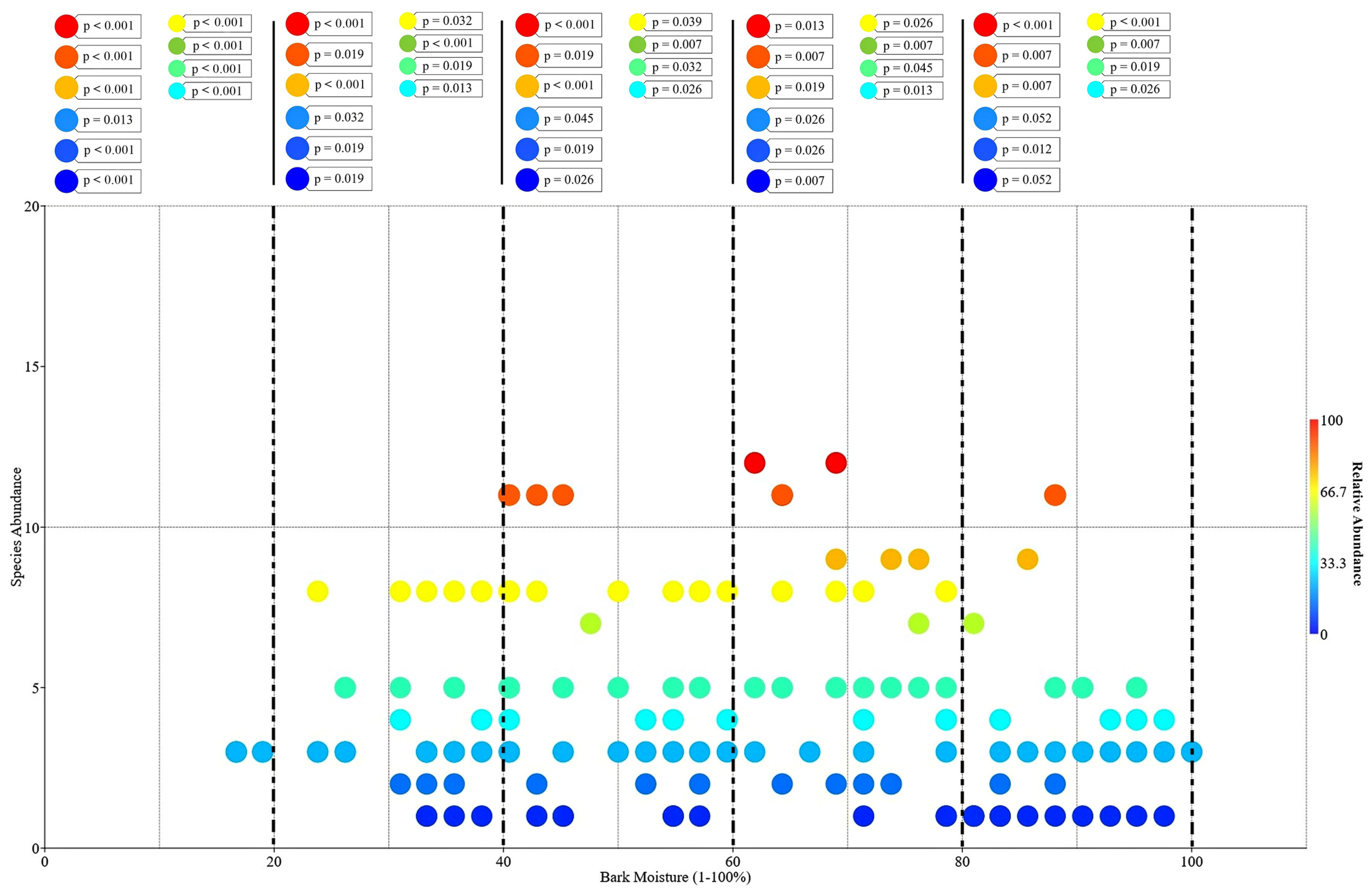
3.4. Factors Influencing Abundance of Epiphytic Orchids: Host Plants and Bark Moisture
4. Discussion
4.1. Ecological Implications of Current Findings on Orchid Diversity
4.2. Applications of the Current Findings for Informing Conservation and Restoration of Orchids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cribb, P.J.; Kell, S.P.; Dixon, K.W.; Barrett, R.L. Orchid Conservation: A Global Perspective. In Orchid Conservation; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2003; pp. 1–24. [Google Scholar]
- Nöske, N.; Hilt, N.; Werner, F.; Brehm, G.; Fiedler, K.; Sipman, H.; Gradstein, S. Disturbance effects on diversity in montane forest of Ecuador: Sessile epiphytes versus mobile moths. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Timsina, B.; Rokaya, M.B.; Münzbergová, Z.; Kindlmann, P.; Shrestha, B.; Bhattarai, B.; Raskoti, B.B. Diversity, distribution and host-species associations of epiphytic orchids in Nepal. Biodivers. Conserv. 2016, 25, 2803–2819. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S.; Rokaya, M.B.; Bhattarai, P.; Gruppe, A. Diversity, composition and host-species relationships of epiphytic orchids and ferns in two forests in Nepal. J. Mt. Sci. 2017, 14, 1065–1075. [Google Scholar] [CrossRef]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of tree age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; García-Franco, J.G. The relationship between tree size and epiphyte species richness: Testing four different hypotheses. J. Biogeogr. 2006, 33, 323–330. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S. Epiphytic orchids and their ecological niche under anthropogenic influence in central Himalayas, Nepal. J. Mt. Sci. 2016, 13, 774–784. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Zimmerman, J.K.; Lebrón, L.; Bayman, P.; Sastre, I.; Axelrod, F.; Alers-García, J. Host specificity and low reproductive success in the rare endemic Puerto Rican orchid Lepanthes caritensis. Biol. Conserv. 1998, 85, 297–304. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Dunn, R.R.; Harris, N.C. Coextinction and persistence of dependent species in a changing world. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 183–203. [Google Scholar] [CrossRef] [Green Version]
- Steege, H.T.; Cornelissen, J.H.C. Distribution and ecology of vascular epiphytes in lowland rain forest of Guyana. Biotropica 1989, 21, 331–339. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, J.; He, J.S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological diversity of orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef]
- Hietz, P. Diversity and Conservation of Epiphytes in a Changing Environment. In Proceedings of the Invited Lecture at the International Conference on Biodiversity and Bioresources: Conservation and Utilization, Phuket, Thailand, 23–27 November 1997; (Published in 1999). Available online: https://media.iupac.org/symposia/proceedings/phuket97/hietz.pdf (accessed on 10 July 2022).
- Callaway, R.M.; Reinhart, K.O.; Moore, G.W.; Moore, D.J.; Pennings, S.C. Epiphyte host preferences and host traits: Mechanisms for species-specific interactions. Oecologia 2002, 132, 221–230. [Google Scholar] [CrossRef]
- Partomihardjo, T.; Eizi, S.; Junichi, Y. Development and distribution of vascular epiphytes communities on the Krakatau Islands, Indonesia. South Pac. Stud. 2004, 25, 7–26. [Google Scholar]
- Hsu, R.C.C.; Wolf, J.H.; Tamis, W.L. Regional and elevational patterns in vascular epiphyte richness on an East Asian Island. Biotropica 2014, 46, 549–555. [Google Scholar] [CrossRef]
- Saw, L.G. Vegetation of Peninsular Malaysia. Seed Plants 2010, 1, 21–41. [Google Scholar]
- Whitmore, T.C. Tropical Rain Forests of the Far East, 2nd ed.; ELBS/Oxford University Press: Oxford, UK, 1984; p. 295. [Google Scholar]
- Kiew, R. A Flora Survey of Limestone Hills in Perlis; Project No. 272/93; World Wildlife Fund: Selangor, Malaysia, 1993; p. 58. [Google Scholar]
- Calderon-Aguilera, L.E.; Rivera-Monroy, V.H.; Porter-Bolland, L.; Martínez-Yrízar, A.; Ladah, L.B.; Martínez-Ramos, M.; Alcocer, J.; Santiago-Pérez, A.L.; Hernandez-Arana, H.A.; Reyes-Gómez, V.M.; et al. An assessment of natural and human disturbance effects on Mexican ecosystems: Current trends and research gaps. Biodivers. Conserv. 2012, 21, 589–617. [Google Scholar] [CrossRef]
- Bridson, D.; Forman, L. The Herbarium Handbook, 3rd ed.; Royal Botanic Gardens: Kew, UK, 2000; p. 348. [Google Scholar]
- Seidenfaden, G.; Wood, J.J. The Orchids of Peninsular Malaysia and Singapore; Royal Botanical Garden: Kew, UK, 1992; p. 779. [Google Scholar]
- Turner, I.M. A catalogue of the vascular plants of Malaya: Orchidaceae. Gard. Bull. Singap. 1995, 47, 559–620. [Google Scholar]
- Comber, J.B. Orchids of Sumatra; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2001; p. 1026. [Google Scholar]
- Ong, P.T.; O’Bryne, P.; Saw, L.G.; Chung, R.C.K. Checklist of Orchids of Peninsular Malaysia; Research Pamphlet (No. 136); Perpustakaan Negara Malaysia: Kuala Lumpur, Malaysia, 2017; p. 169. [Google Scholar]
- Govaerts, R.; Bernet, P.; Kratochvil, K.; Gerlach, G.; Carr, G.; Alrich, P.; Pridgeon, A.M.; Pfahl, J.; Campacci, M.A.; Baptista, D.H.; et al. World Checklist of Orchidaceae; The Board of Trustees of the Royal Botanic Gardens: Kew, UK; Available online: http://apps.kew.org/wcsp/ (accessed on 22 November 2021).
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Cooperation: Kuala Lumpur, Malaysia, 1966; Volume 1 (A–H), p. 1240.
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Cooperation: Kuala Lumpur, Malaysia, 1966; Volume 2 (I–Z), p. 2444.
- Ng, F.S.P. Tree Flora of Malaya; Volume 3; Forest Research Institute of Malaysia: Kepong, Malaysia, 1978; p. 339.
- Ng, F.S.P. Tree Flora of Malaya; Volume 4; Forest Research Institute of Malaysia: Kepong, Malaysia, 1989; p. 549.
- Ridley, H.N. Introduction. In Flora of Malay Peninsula; Ridley, H.N., Ed.; Volume, 1; Reeve & Co.: London, UK, 1922; pp. ix–xx. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 1, Polypetalae; L. Reeve & Company, Limited: London, UK, 1922; p. 470. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 2, Gamopetalae; L. Reeve & Company, Limited: London, UK, 1923; p. 672. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 3, Apetalae; L. Reeve & Company, Limited: London, UK, 1924; p. 406. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 4, Monocotyledones; L. Reeve & Company, Limited: London, UK, 1924; p. 383. [Google Scholar]
- Whitmore, T.C. Tree Flora of Malaya; Volume 1; Forest Research Institute of Malaysia: Kepong, Malaysia, 1972; p. 473.
- Whitmore, T.C. Tree Flora of Malaya; Volume 2; Forest Research Institute of Malaysia: Kepong, Malaysia, 1973; p. 444.
- Ingram, S.W.; Lowman, M.D. The collection and preservation of plant material from the tropical forest canopy. For. Canopies 1995, 22, 587–603. Available online: https://canopymeg.com/PDFs/papers/0047.pdf (accessed on 12 June 2021).
- Migenis, L.E.; Ackerman, J.D. Orchid—Phorophyte relationships in a forest watershed in Puerto Rico. J. Trop. Ecol. 1993, 9, 231–240. [Google Scholar] [CrossRef]
- Werner, F.A.; Gradstein, S.R. Diversity of dry forest epiphytes along a gradient of human disturbance in the tropical Andes. J. Veg. Sci. 2009, 20, 59–68. [Google Scholar] [CrossRef]
- McCree, K.J. Tests of Current Definitions of Photosynthetically Active Radiation Against Leaf Photosynthesis Data. Agric. For. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- Thimijan, R.W.; Heins, R.D. Photometric, Radiometric, and Quantum Light Units of Measure: A Review of Procedures for Interconversion. Hortic. Sci. 1983, 18, 818–822. [Google Scholar] [CrossRef]
- Harper, D.A.T. Numerical Palaeobiology–Computer-Based Modelling and Analysis of Fossils and Their Distributions; John Wiley & Sons: Oxford, UK, 1999; p. 468. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 12 June 2021).
- Gorelick, R. Combining richness and abundance into a single diversity index using matrix analogues of Shannon’s and Simpson’s indices. Ecography 2006, 29, 525–530. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Biodiversity; Blackwell Publishing: Malden, MA, USA, 2004; p. 256. [Google Scholar]
- Help, C. A new index measuring evenness. J. Mar. Biol. Assoc. United Kingd. 1974, 54, 555–557. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Besi, E.E.; Nikong, D.; Mustafa, M.; Go, R. Orchid diversity in anthropogenic-induced degraded tropical rainforest, an extrapolation towards conservation. Lankesteriana 2019, 19, 107–124. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9; User’s Guide and application; 2013. Available online: http://purl.oclc.org/estimates (accessed on 13 December 2021).
- Wagner, K.; Mendieta-Leiva, G.; Zotz, G. Host specificity in vascular epiphytes: A review of methodology, empirical evidence and potential mechanisms. AoB PLANTS 2015, 7, plu092. [Google Scholar] [CrossRef] [Green Version]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: New York, NY, USA, 1985; p. 472. [Google Scholar] [CrossRef]
- Turner, M.G.; Dale, V.H.; Everham, E.H. Fires, hurricanes, and volcanoes: Comparing large disturbances. BioScience 1997, 47, 758–768. [Google Scholar] [CrossRef]
- Turner, M.G.; Dale, V.H. Comparing large, infrequent disturbances: What have we learned? Ecosystems 1998, 1, 493–496. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Tecco, P.A.; Díaz, S. Plant Invasions in Undisturbed Ecosystems: The Triggering Attribute Approach. J. Veg. Sci. 2005, 16, 723–728. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Gandaseca, S.; Abd Wahab, N.L.; Pazi, A.M.; Rosli, N.; Zaki, P.H. Comparison of water quality status of undisturbed and disturbed mangrove forest at Awat-Awat Lawas Sarawak. Open J. For. 2016, 6, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Jamaluddin, J.; Azhar, B.; Chong, P. Estimating Wildlife Occurrence in Peninsular Malaysia Using GWR4. Malays. For. 2018, 81, 55–63. [Google Scholar]
- MacArthur, R.H. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, H.S.; Fischer, A. Host tree utilization by epiphytic orchids in different land-use intensities in Kathmandu valley, Nepal. Plant Ecol. 2012, 213, 1393–1412. [Google Scholar] [CrossRef]
- Preston, F.W. The canonical distribution of commonness and rarity: Part I. Ecology 1962, 43, 185–215. [Google Scholar] [CrossRef]
- Connor, E.F.; McCoy, E.D. Species–Area Relationships. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2002; p. 10. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P.; Larsen, T.H.; Docherty, T.D.; Ansell, F.A.; Hsu, W.W.; Derhé, M.A.; Hamer, K.C.; Wilcove, D.S. Degraded lands worth protecting: The biological importance of Southeast Asia’s repeatedly logged forests. Proc. R. Soc. B Biol. Sci. 2011, 278, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohnwald, S. Beiträge zum Mikroklima interandiner Trockentäler Boliviens. Geoökodynamik 1999, 20, 221–229. [Google Scholar]
- Gradstein, S.R. Epiphytes of Tropical Montane Forests—Impact of Deforestation and Climate Change. In The Tropical Mountain Forest: Patterns and Processes in a Biodiversity Hotspot; Gradstein, S.R., Homeier, J., Gansert, D., Eds.; Biodiversity and Ecology Series: Göttingen, Germany, 2008; pp. 51–65. [Google Scholar]
- Gentry, A.H.; Dodson, C.H. Contribution of non-trees to species richness of a tropical rain forest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Wolf, J.H.D. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the northern Andes. Ann. Mo. Bot. Gard. 1993, 80, 928–960. [Google Scholar] [CrossRef]
- Kreft, H.; Köster, N.; Küper, W.; Nieder, J.; Barthlott, W. Diversity and biogeography of vascular epiphytes in Western Amazonia, Yasuni, Ecuador. J. Biogeogr. 2004, 31, 1463–1476. [Google Scholar] [CrossRef]
- Sanford, W.W. The Ecology of Orchids. In The Orchids (Scientific Studies); Withner, C.L., Ed.; Wiley: New York, NY, USA, 1974; p. 100. [Google Scholar]
- Gradstein, S.R.; Churchill, S.P.; Salazar Allen, N. Guide to the bryophytes of tropical America. Mem. N. Y. Bot. Gard. 2001, 86, 577. [Google Scholar]
- Richardson, D.H.S. The Biology of Mosses; Halsted Press: New York, NY, USA, 1981; p. 220. [Google Scholar]
- Alpert, P.; Oechel, W.C. Carbon balance limits the microdistribution of Grimmia laevigata, a desiccation-tolerant plant. Ecology 1985, 66, 660–669. [Google Scholar] [CrossRef]
- Cribb, P.; Butterfield, I. The Genus Pleione; Royal Botanic Gardens: Victoria, Australia, 1999; p. 129. [Google Scholar]
- Zhang, W.; Hu, H.; Zhang, S.B. Divergent adaptive strategies by two co-occurring epiphytic orchids to water stress: Escape or avoidance? Front. Plant Sci. 2016, 7, 588. [Google Scholar] [CrossRef] [Green Version]
- Stratton, L.; Goldstein, G.; Meinzer, F.C. Stem water storage capacity and efficiency of water transport: Their functional significance in a Hawaiian dry forest. Plant Cell Environ. 2000, 23, 99–106. [Google Scholar] [CrossRef]
- Köster, N.; Nieder, J.; Barthlott, W. Effect of host tree traits on epiphyte diversity in natural and anthropogenic habitats in Ecuador. Biotropica 2011, 43, 685–694. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S. Micro-site conditions of epiphytic orchids in a human impact gradient in Kathmandu valley, Nepal. J. Mt. Sci. 2012, 9, 331–342. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Lobakova, E.S.; Kolomeitseva, G.L.; Netrusov, A.I. Microbiota of the orchid rhizoplane. Microbiology 2001, 70, 492–497. [Google Scholar] [CrossRef]
- Herrera, H.; Fuentes, A.; Soto, J.; Valadares, R.; Arriagada, C. Orchid-Associated Bacteria and Their Plant Growth Promotion Capabilities. In Orchids Phytochemistry, Biology and Horticulture; Merillon, J.M., Kodja, H., Eds.; Reference Series in Phytochemistry; Springer: New York, NY, USA, 2021; p. 26. [Google Scholar] [CrossRef]
- Bates, B.; Kundzewicz, Z.; Wu, S. Climate Change and Water; Intergovernmental Panel on Climate Change Secretariat: Geneva, Switzerland, 2008; p. 210. [Google Scholar]
- Barthlott, W.; Schmit-Neuerburg, V.; Nieder, J.; Engwald, S. Diversity and abundance of vascular epiphytes: A comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecol. 2001, 152, 145–156. [Google Scholar] [CrossRef]
- Krömer, T.; Gradstein, S.R. Species richness of vascular epiphytes in two primary forests and fallows in the Bolivian Andes. Selbyana 2003, 24, 190–195. [Google Scholar]
- Acebey, A.; Gradstein, S.R.; Krömer, T. Species richness and habitat diversification of corticolous bryophytes in submontane rain forest and fallows of Bolivia. J. Trop. Ecol. 2003, 19, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Drehwald, U. Biomonitoring of disturbance in neotropical rainforests using bryophytes as indicators. J. Hattori Bot. Lab. 2005, 97, 117–126. [Google Scholar] [CrossRef]
- Holz, I.; Gradstein, S.R. Phytogeography of the bryophyte floras of oak forests and pâramo of the Cordillera de Talamanca, Costa Rica. J. Biogeogr. 2005, 32, 1591–1609. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Masino, S.A.; Faison, E.K. Intact forests in the United States: Proforestation mitigates climate change and serves the greatest good. Front. For. Glob. Change 2019, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F. Forest-climate interactions in fragmented tropical landscapes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, R.D. The microevolutionary consequences of climate change. Trends Ecol. Evol. 1990, 5, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Feng, C.L.; Luo, Y.B.; Chen, B.S.; Wang, Z.S.; Gu, H.Y. Potential challenges of climate change to orchid conservation in a wild orchid hotspot in southwestern China. Bot. Rev. 2010, 76, 174–192. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. Parasitic plants: Parallels and contrasts with herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef]
- Crain, B.J.; Tremblay, R.L. Update on the distribution of Lepanthes caritensis, a rare Puerto Rican endemic orchid. Endanger. Species Res. 2012, 18, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Merwin, M.C.; Rentmeester, S.A.; Nadkarni, N.M. The influence of host tree species on the distribution of epiphytic bromeliads in experimental monospecific plantations, La Selva, Costa Rica. Biotropica 2003, 35, 37–47. [Google Scholar] [CrossRef]
- Johansson, D. Ecology of vascular epiphytes in West African rain forests. Acta Phytogeogr. Suec. 1974, 59, 1–129. Available online: https://www.diva-portal.org/smash/get/diva2:565496/FULLTEXT01.pdf (accessed on 12 May 2020).
- Frei, J.K.; Dodson, C.H. The chemical effect of certain bark substrates on the germination and early growth of epiphytic orchids. Bull. Torrey Bot. Club 1972, 99, 301–307. [Google Scholar] [CrossRef]
- Laube, S.; Zotz, G. Neither host-specific nor random: Vascular epiphytes on three tree species in a Panamanian lowland forest. Ann. Bot. 2006, 97, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Zotz, G. Johansson revisited: The spatial structure of epiphyte assemblages. J. Veg. Sci. 2007, 18, 123–130. [Google Scholar] [CrossRef]
- Annaselvam, J.; Parthasarathy, N. Diversity and distribution of herbaceous vascular epiphytes in a tropical evergreen forest at Varagalaiar, Western Ghats, India. Biodivers. Conserv. 2001, 10, 317–329. [Google Scholar] [CrossRef]
- Mucunguzi, P. Diversity and distribution of epiphytic orchids in Kibale National Park, Uganda. Selbyana 2008, 29, 217–225. [Google Scholar]
- Ling, C.Y.; Sang, J. Diversity of Orchidaceae from Murum Dam, Belaga, Sarawak, Borneo. Sibbaldia 2018, 16, 67–85. Available online: https://core.ac.uk/download/pdf/267015996.pdf (accessed on 3 January 2020).
- Besi, E.E.; Nikong, D.; Pungga, R.S.; Go, R. Wild orchid diversity of highland forest in the Heart of Borneo: Long Banga and Tama Abu, Sarawak. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 125–135. [Google Scholar] [CrossRef]
- Tupac Otero, J.; Aragón, S.; Ackerman, J.D. Site variation in spatial aggregation and phorophyte preference in Psychilis monensis (Orchidaceae). Biotropica 2007, 39, 227–231. [Google Scholar] [CrossRef]
- de la Rosa-Manzano, E.; Andrade, J.L.; Zotz, G.; Reyes-García, C. Epiphytic orchids in tropical dry forests of Yucatan, Mexico–Species occurrence, abundance and correlations with host tree characteristics and environmental conditions. Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 100–109. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Olmsted, I.C. Host tree utilization by vascular epiphytes in a seasonally inundated forest (Tintal) in Mexico. Biotropica 1992, 24, 402–407. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A. Distribution pattern of the epiphytic orchid Rhynchostylis retusa under strong human influence in Kathmandu valley, Nepal. Bot. Orient. J. Plant Sci. 2011, 8, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Reinhart, K.O.; Tucker, S.C.; Pennings, S.C. Effects of epiphytic lichens on host preference of the vascular epiphyte Tillandsia usneoides. Oikos 2001, 94, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Watthana, S.; Pedersen, H.Æ. Phorophyte diversity, substrate requirements and fruit set in Dendrobium scabrilingue Lindl. (Asparagales: Orchidaceae): Basic observations for re-introduction experiments. Trop. Nat. Hist. 2008, 8, 135–142. Available online: https://li01.tci-thaijo.org/index.php/tnh/article/view/102971 (accessed on 15 January 2020).
- Ibisch, P. Neotropische Epiphytendiversität—Das Beispiel Bolivien; Galunder Verlag: Wiehl, Germany, 1996; p. 357. [Google Scholar]
- Wolseley, P.A.; Aguirre-Hudson, B. The ecology and distribution of lichens in tropical deciduous and evergreen forests of northern Thailand. J. Biogeogr. 1997, 24, 327–343. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Pauleit, S. Sustainable conservation perspectives for epiphytic orchids in the central Himalayas, Nepal. Appl. Ecol. Environ. Res. 2015, 13, 753–767. [Google Scholar] [CrossRef]
- Ong, P.T. Limestone orchids of Peninsular Malaysia. Orchid Rev. 2015, 123, 28–36. [Google Scholar]
- Porembski, S. Epiphytic orchids on arborescent Velloziaceae and Cyperaceae: Extremes of phorophyte specialisation. Nord. J. Bot. 2003, 23, 505–512. [Google Scholar] [CrossRef]
- Morris, B. The epiphytic orchids of the Shire Highlands, Malawi. Proc. Linn. Soc. Lond. 1968, 179, 51–66. [Google Scholar] [CrossRef]
- La Croix, I.F.; La Croix, E.A.S.; La Croix, T.M. Orchids of Malawi: The Epiphytic and Terrestrial Orchids from South and East Central Africa; Taylor & Francis: Rotterdam, The Netherlands, 1991; p. 358. [Google Scholar]
- Matias, L.Q.; Braga, P.I.S.; Freire, A.G. Reproductive biology of Constantia cipoensis Porto & Brade (Orchidaceae), an endemic species from the Serra do Cipo, Minas Gerais. Rev. Bras. De Botânica 1996, 19, 119–125. [Google Scholar]
- da Mota, R.C.; de Barros, F.; Stehmann, J.R. Two new species of Orchidaceae from Brazil: Bulbophyllum carassense and Lepanthopsis vellozicola. Novon J. Bot. Nomencl. 2009, 19, 380–387. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | States | Vegetation Types | Elevation (m) | No. of Transect Plots | Totals Transect Size/Area (km2) |
|---|---|---|---|---|---|
| Padang 7 (Rompin) (PG7) | Pahang | Hill mixed dipterocarp forest (with coastal mountain outcrops or kerangas) | 268–600 | 4 | 0.9 |
| Bukit Rongkit (BR) | Perlis | Hill limestone forest | 300–400 | 2 | 14.4 |
| Taman Rimba Komanwel (TRK) | Selangor | Lowland dipterocarp forest | 100–200 | 6 | 16.8 |
| Bukit Batu Kapal (BK) | Terengganu | Lowland riparian forest | 100–200 | 5 | 5.6 |
| Lata Kekabu (KB) | Perak | Lowland riparian forest | 100–200 | 5 | 5.5 |
| Lata Lawin (LW) | Perak | Lowland riparian forest | 100–200 | 3 | 3.6 |
| Lata Tanjung Kala (TK) | Perak | Lowland riparian forest | 100–200 | 2 | 12.0 |
| Bukit Pedu (BP) | Kedah | Conglomerate hill forest | 200–400 | 1 | 9.0 |
| Total | 28 | 67.8 | |||
| Site | State | Vegetation Types | S | E | F | D | PT | DI | Disturbance Regime | Disturbance Origin | Elevation (m) | No. of Transect Plots | Totals Transect Size/Area (km2) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bukit Sekayu (BS) | Terengganu | Lowland riparian forest | 1 | 1 | 1 | 3 | 1 | 3 | Low | Fragmentation caused by palm oil estate | 90–100 | 1 | 11.8 |
| Gunung Perlis (GP) | Perlis | Hill dipterocarp forest (granite) | 1 | 1 | 1 | 3 | 1 | 3 | Low | Erosion of trail due to heavy ecotourism activities | 500–800 | 4 | 34.0 |
| Bukit Batu Kapal [BK(M)] | Terengganu | Mixed dipterocarp forest | 1 | 1 | 1 | 1 | 1 | 1 | Low | Timber extraction (a forest margin bordered by logged forests) | 100–400–600 | 1 | 12.6 |
| Tanah Merah (TM) | Kelantan | Lowland forest | 1 | 1 | 1 | 1 | 1 | 1 | Low | Timber extraction, plantation, mud-flood | 40–100 | 2 | 0.3 |
| Hulu Setiu (LHS) | Terengganu | Lowland dipterocarp forest | 2 | 2 | 1 | 2 | 1 | 8 | Mild | Timber extraction | 40–100 | 6 | 13.0 |
| Kuala Koh (KH) | Kelantan | Lowland dipterocarp forest | 2 | 2 | 1 | 2 | 1 | 8 | Mild | Timber extraction (an encroachment), mud-flood | 90–130 | 8 | 0.8 |
| Bukit Batu Kapal (LBK) | Terengganu | Hill dipterocarp forest | 3 | 2 | 2 | 2 | 1 | 24 | Severe | Timber extraction | 300–500 | 1 | 33.3 |
| Bintang Hijau (LB) | Perak | Lowland dipterocarp forest | 3 | 2 | 3 | 3 | 2 | 108 | Severe | Timber extraction | 100–200 | 3 | 1200.0 |
| Gawi (LG) | Terengganu | Lowland dipterocarp forest | 3 | 2 | 3 | 3 | 2 | 108 | Severe | Timber extraction | 90–250 | 1 | 19.5 |
| Petuang (LP) | Terengganu | Lowland dipterocarp forest | 3 | 2 | 3 | 3 | 2 | 108 | Severe | Timber extraction | 200–330 | 2 | 53.0 |
| Total | 29 | 1409.7 | |||||||||||
| Variables | Definition | Scoring Value |
|---|---|---|
| Severity (S) | Loss in ecosystem quality based on the different types of disturbance | (1) small (disturbed ground vegetation due to flood or heavy human trampling) (2) medium (disturbed ground vegetation and forest canopy due to fallen trees and mud flood) (3) large (massive removal of ground vegetation and forest canopy due to clear-felled timber extraction and logs’ dragging extraction by tractor |
| Extension (E) | Area affected by a given disturbance | (1) small (>20% of area) (2) medium (>20% ≤ 50% of area) (3) large (>50% of area) |
| Frequency (F) | Occurrence of disturbance event in a given time period | (1) low (occurring at intervals longer than 10 years) (2) medium (once every 2–10 years) (3) high (once every year or seasonal) |
| Duration (D) | Length in time of the disturbance event | (1) <1 year (2) <3 years (3) >3 years |
| Patch type (PT) | Type of habitat patch created by the disturbance | (1) embedded (disturbed patches within a continuous undisturbed ecosystem) (2) isolated (disturbed patches isolated from an undisturbed ecosystem) |
| Parameters | Undisturbed Forests | Disturbed Forests | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PG7 | BR | TRK | BK | KB | LW | TK | BP | BS | GP | BK(M) | TM | LHS | KH | LBK | LB | LG | LP | |
| Taxa_S | 27 | 26 | 5 | 17 | 16 | 4 | 9 | 14 | 25 | 19 | 7 | 4 | 12 | 6 | 5 | 11 | 83 | 61 |
| Occurrence (ni) | 34 | 39 | 8 | 22 | 17 | 5 | 17 | 21 | 58 | 21 | 16 | 7 | 12 | 19 | 5 | 15 | 98 | 70 |
| Occurrence (ni) (Epiphytic orchids) | 31 | 28 | 7 | 21 | 17 | 4 | 17 | 6 | 49 | 19 | 17 | 10 | 13 | 11 | 5 | 16 | 70 | 57 |
| Occurrence (ni) (Terrestrial orchids) | 4 | 8 | 1 | 2 | 2 | 1 | 0 | 1 | 6 | 2 | 0 | 0 | 0 | 8 | 0 | 0 | 0 | 0 |
| Occurrence (ni) (Lithophytic orchids) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 15 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Occurrence (ni) (Mycoheterotroph orchids) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Dominance_D | 0.047 | 0.049 | 0.250 | 0.066 | 0.066 | 0.280 | 0.142 | 0.098 | 0.092 | 0.057 | 0.258 | 0.306 | 0.083 | 0.191 | 0.200 | 0.102 | 0.015 | 0.018 |
| Simpson_1-D | 0.953 | 0.951 | 0.750 | 0.934 | 0.934 | 0.720 | 0.858 | 0.903 | 0.908 | 0.943 | 0.742 | 0.694 | 0.917 | 0.809 | 0.800 | 0.898 | 0.985 | 0.982 |
| Shannon_H | 3.200 | 3.139 | 1.494 | 2.776 | 2.752 | 1.332 | 2.069 | 2.491 | 2.801 | 2.912 | 1.629 | 1.277 | 2.485 | 1.709 | 1.609 | 2.338 | 4.342 | 4.063 |
| Evenness_e^H/S | 0.909 | 0.888 | 0.891 | 0.944 | 0.979 | 0.947 | 0.879 | 0.863 | 0.658 | 0.969 | 0.728 | 0.897 | 1.000 | 0.920 | 1.000 | 0.942 | 0.926 | 0.953 |
| Area (km2) | 0.900 | 14.400 | 16.800 | 5.618 | 5.500 | 3.600 | 12 | 9 | 11.760 | 34 | 12.600 | 0.250 | 13 | 0.800 | 33.300 | 1200 | 19.500 | 53 |
| Density (plants/km2) | 37.800 | 2.700 | 0.500 | 3.900 | 3.100 | 1.400 | 1.400 | 2.300 | 4.900 | 0.600 | 1.300 | 28 | 0.900 | 23.800 | 0.200 | 0 | 5 | 1.300 |
| Density of epiphytic orchids (plants/km2) | 34.400 | 1.900 | 0.400 | 3.700 | 3.100 | 1.100 | 1.400 | 0.700 | 4.200 | 0.600 | 1.300 | 40 | 1 | 13.800 | 0.200 | 0 | 3.6 | 1.1 |
| Density of terrestrial orchids (plants/km2) | 4.400 | 0.600 | 0.100 | 0.400 | 0.400 | 0.300 | 0 | 0.100 | 0.500 | 0.100 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 |
| Density of lithophytic orchids (plants/km2) | 0 | 0.100 | 0 | 0 | 0 | 0 | 0 | 1.700 | 0.300 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Density of mycoheterotroph orchids (plants/km2) | 1.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Site | Temperature (°C) | Relative Air Humidity (1%–100%) | Light Intensity, PPFD (µmol m−2 s−1) | Elevation (m) |
|---|---|---|---|---|
| Undisturbed vs. disturbed forests (p < 0.005, n = 30, df = 58) | ||||
| Undisturbed forests | Min: 27.8 ± 0.3 a | Min: 77.1 ± 1.2 a | Min: 23.8 ± 3.3 a | 100–600 |
| Max: 31.2 ± 0.2 a | Max: 89.6 ± 0.9 a | Max: 171.7 ± 18.8 a | ||
| Disturbed forests | Min: 27.2 ± 0.3 a | Min: 64.5 ± 2.3 b | Min: 107.1 ± 16 b | 40–800 |
| Max: 35.7 ± 0.8 b | Max: 88.7 ± 1 a | Max: 456.5 ± 29.6 b | ||
| Adjacent undisturbed vs. disturbed forests (p < 0.005, n = 20, df = 38) | ||||
| Undisturbed forests | Min: 30.6 ± 0.1 a | Min: 86.5 ± 0.6 a | Min: 21.5 ± 2.2 a | 100–200 |
| Max: 30.7 ± 0.1 a | Max: 88.2 ± 0.6 a | Max: 125.8 ± 17 a | ||
| Disturbed forests | Min: 32.2 ± 0.6 a | Min: 44.8 ± 1.2 b | Min: 51.9 ± 7.7 b | 200–500 |
| Max: 40.6 ± 0.3 b | Max: 75.8 ± 1.5 b | Max: 406.8 ± 44.3 b | ||
| Undisturbed and extremely disturbed (logging sites) forests (p < 0.005, n = 20, df = 38) | ||||
| Undisturbed forests | Min: 28.1 ± 0.3 a | Min: 80.7 ± 1 a | Min: 33.4 ± 5 a | 100–600 |
| Max: 31.3 ± 0.3 a | Max: 89 ± 0.5 a | Max: 190.9 ± 24.7 a | ||
| Logging sites | Min: 32.9 ± 0.7 b | Min: 51.8 ± 2.4 b | Min: 132.1 ± 16.6 b | 200–500 |
| Max: 40.2 ± 0.4 b | Max: 79.5 ± 1.7 b | Max: 410.5 ± 34.5 b | ||
| Type | Habit | Crown Form | Crown Class | Substrate Cover | Height (m) | Species |
|---|---|---|---|---|---|---|
| 1 | Tree | Round | Overtopped | Basal | ca. 3–6 | Greenea corymbosa (Jack) Voigt (Rubiaceae), Streblus ilicifolius (S.Vidal) Corner (Moraceae) |
| 2 | Tree | Broad | Codominant | Trunk | ca. 20–30 | Castanopsis sp. (Fagaceae), Artocarpus elasticus Reinw. ex Blume (Moraceae) |
| 3 | Tree | Spreading | Dominant | Trunk | ca. 40 | Campnosperma auriculatum (Blume) Hook.f. (Anacardiaceae), Castanopsis sp. (Fagaceae) |
| 4 | Tree | Spreading | Intermediate | Basal, trunk, branches | ca. 5 | Ficus fistulosa Reinw. ex Blume (Moraceae), Polyalthia cauliflora Hook.f. & Thomson (Annonaceae), Goniothalamus sp. (Annonaceae), Tristaniopsis whiteana (Griff.) Peter G.Wilson & J.T.Waterh. (Myrtaceae), Syzygium cf. salictoides (Ridl.) I.M.Turner (Myrtaceae) |
| 5 | Tree | Columnar | Intermediate | Basal, trunk | ca. 15–20 | Syzygium sp. (Myrtaceae), Castanopsis sp. (Fagaceae) |
| 6 | Tree | Spreading | Codominant | Basal | ca. 20 | Dipterocarpus elongatus Korth. (Dipterocarpaceae), Dipterocarpus oblongifolius Blume (Dipterocarpaceae) |
| 7 | Tree | Vase | Intermediate | Basal, trunk, branches | ca. 15 | Playmitra macrocarpa Boetl. (Annonaceae) |
| 8 | Tree | Weeping | Overtopped | Basal, trunk, branches | ca. 15 | Hydnocarpus illicifolius King (Achariaceae) |
| 9 | Tree | Globular | Dominant | Basal, trunk, branches | ca. 7–10 | Calophyllum wallichianum Planch. & Triana (Calophyllaceae), Leptospermum polygalifolium Salisb. (Myrtaceae) |
| 10 | Tree | Globular | Intermediate | Basal | ca. 10 | Macaranga gigantea (Rchb.f. & Zoll.) Müll.Arg. (Euphorbiaceae) |
| 11 | Tree | Round | Intermediate | Branches | ca. 20 | Goniothalamus sp. (Annonaceae) |
| 12 | Tree | Oval | Dominant | Basal | ca. 40 | Ochanostachys amentacea Mast.(Olacaceae) |
| 13 | Tree | Broad | Intermediate | Basal, branches | ca. 10 | Neonauclea pallida subsp. malaccensis (Gand.) Ridsdale (Rubiaceae), Saraca declinata Miq. (Fabaceae), Eurya cf. acuminata DC (Pentaphylacaceae) |
| 14 | Liana | Leaflet wholly arranged | Intermediate | Stem | ca. 20 | Spatholobus ferrugineus (Zoll. & Moritzi) Benth. (Fabaceae) |
| 15 | Liana | Spreading | Overtopped | Stem | ca. 6 | Apocynaceae |
| 16 | Fallen tree | Spreading | Emergent | Branches, trunk | ca. 50 | Dipterocarpaceae |
| 17 | Fallen tree | Round | Codominant | Branches | ca. 20–30 | Ficus sp. (Moraceae), Artocarpus elasticus (Moraceae) |
| 18 | Fallen tree | Spreading | Dominant | Branches, trunk | ca. 40 | Shorea sp. (Balau) (Dipterocarpaceae), Hopea sp. (Dipterocarpaceae), Pentace eximia (Malvaceae) |
| 19 | Fallen tree | Broad | Codominant | Trunk | ca. 30 | Cynometra malaccensis Meeuwen (Fabaceae), Parkia sp. (Fabaceae) |
| Types | Bark Textures | Sites | Forest Types |
|---|---|---|---|
| 1 | Fissured | 1, 2, 3, 5, 8, 9, 10, 11, 12, 13, 14, 17, 18 | Undisturbed, disturbed |
| 2 | Fissured, lenticeled | 1 | Disturbed |
| 3 | Fissured, peeled off | 5, 6 | Disturbed |
| 4 | Fissured, peeled off, soft | 8 | Undisturbed |
| 5 | Fissured, shedding | 3 | Disturbed |
| 6 | Peeled off, dry | 1, 5, 13, 15, 16 | Disturbed |
| 7 | Ridged, warty | 5 | Disturbed |
| 8 | Smooth | 2, 4, 14 | Undisturbed, disturbed |
| 9 | Smooth to fissured, moist | 2, 3, 7, 17, 18 | Disturbed |
| 10 | Smooth to fissured, warty | 9, 12 | Undisturbed |
| 11 | Smooth, lenticeled | 2, 4, 8 | Undisturbed |
| 12 | Soft, small fissured, warty | 3, 5 | Disturbed |
| 13 | Strongly fissured | 9, 10, 11 | Undisturbed |
| 14 | Strongly fissured, shedding | 6, 13, 15, 16 | Disturbed |
| 15 | Strongly fissured, soft, dry | 1, 13 | Disturbed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Besi, E.E.; Mustafa, M.; Yong, C.S.Y.; Go, R. Habitat Ecology, Structure Influence Diversity, and Host-Species Associations of Wild Orchids in Undisturbed and Disturbed Forests in Peninsular Malaysia. Forests 2023, 14, 544. https://doi.org/10.3390/f14030544
Besi EE, Mustafa M, Yong CSY, Go R. Habitat Ecology, Structure Influence Diversity, and Host-Species Associations of Wild Orchids in Undisturbed and Disturbed Forests in Peninsular Malaysia. Forests. 2023; 14(3):544. https://doi.org/10.3390/f14030544
Chicago/Turabian StyleBesi, Edward Entalai, Muskhazli Mustafa, Christina Seok Yien Yong, and Rusea Go. 2023. "Habitat Ecology, Structure Influence Diversity, and Host-Species Associations of Wild Orchids in Undisturbed and Disturbed Forests in Peninsular Malaysia" Forests 14, no. 3: 544. https://doi.org/10.3390/f14030544
APA StyleBesi, E. E., Mustafa, M., Yong, C. S. Y., & Go, R. (2023). Habitat Ecology, Structure Influence Diversity, and Host-Species Associations of Wild Orchids in Undisturbed and Disturbed Forests in Peninsular Malaysia. Forests, 14(3), 544. https://doi.org/10.3390/f14030544