


Paleo Distribution and Habitat Risks under Climate Change of Helleborus thibetanus
Abstract
:1. Introduction
2. Materials and Methods
2.1. Occurrence Data and Distribution Range
2.2. Environmental Data and Correlation Analysis
2.3. Species Distribution Model Tuning and Construction
2.4. Potential Distribution Prediction and Geographical Analysis
3. Results
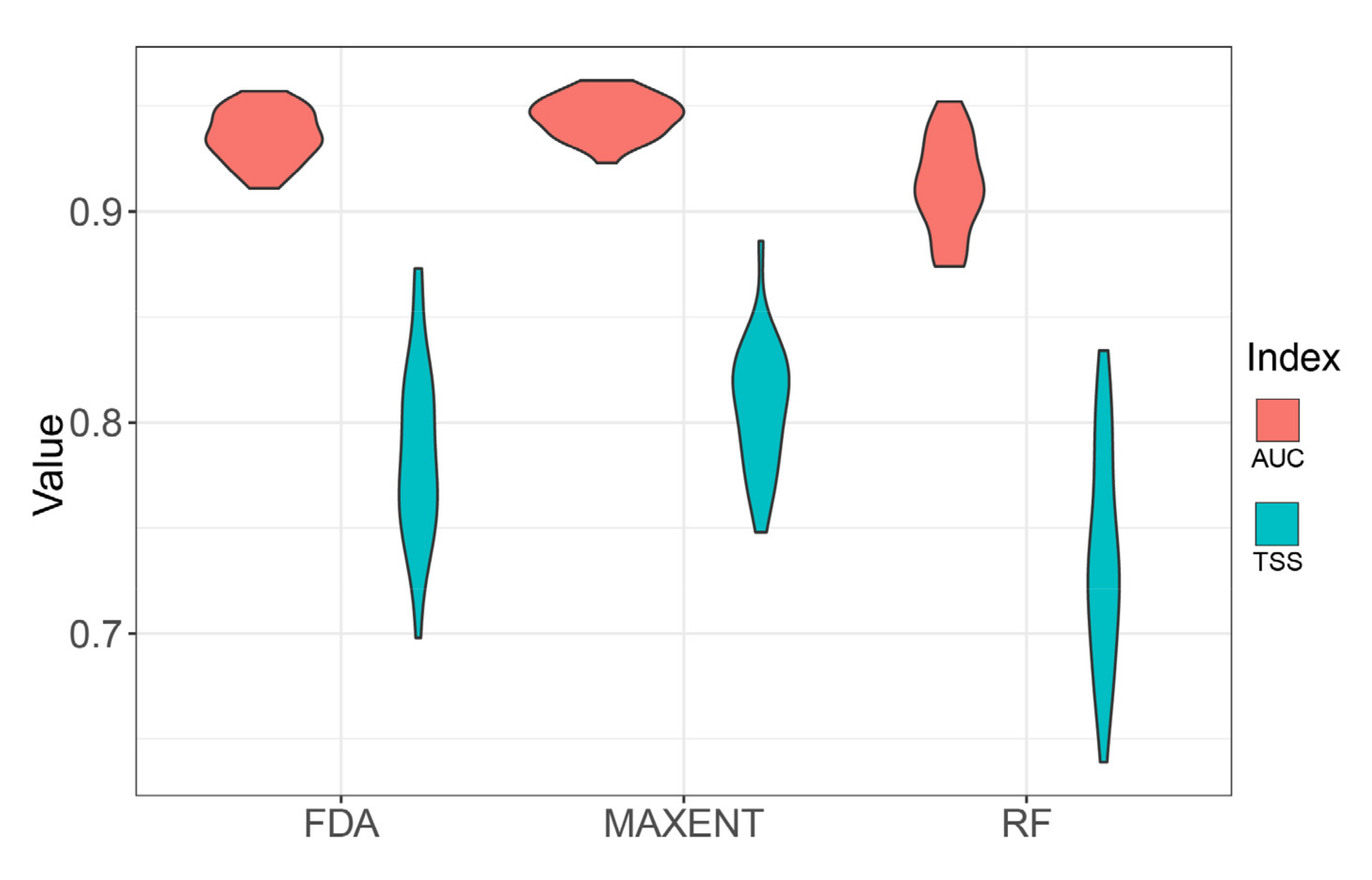
3.1. Single Model Accuracy and Ensembled Models
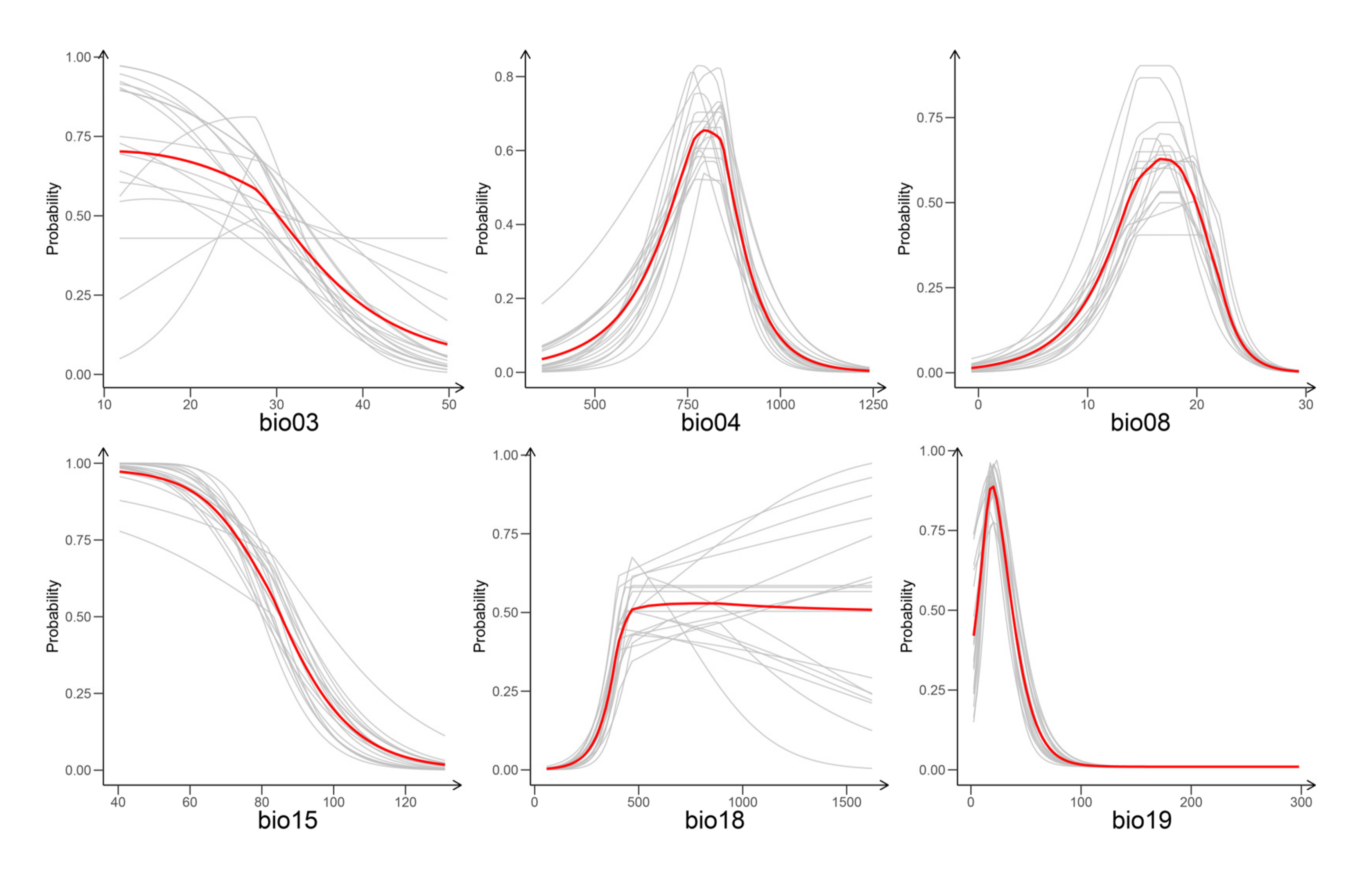
3.2. Climatic Niche and Proximal Variables
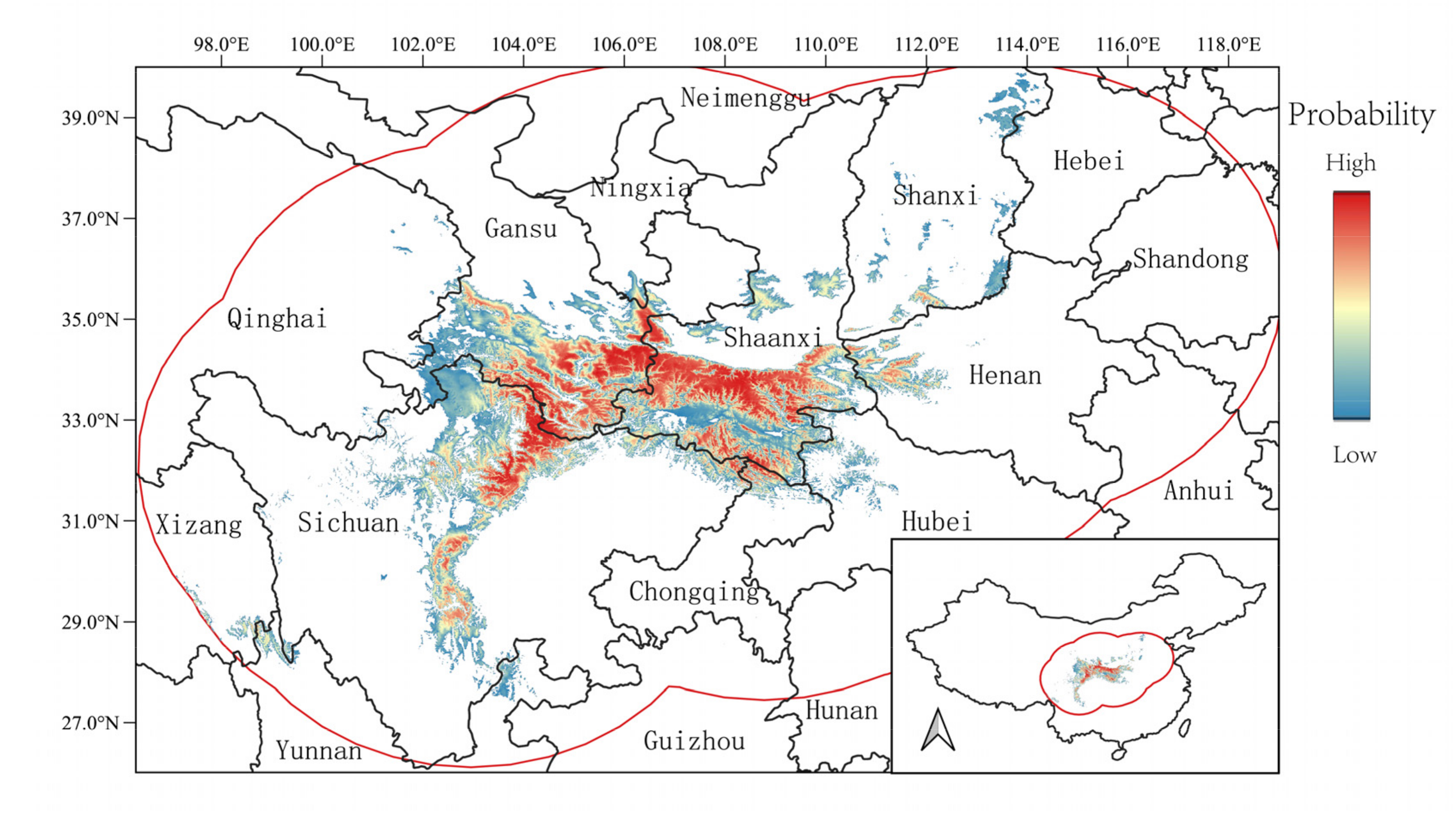
3.3. Current Potential Distribution of H. thibetanus
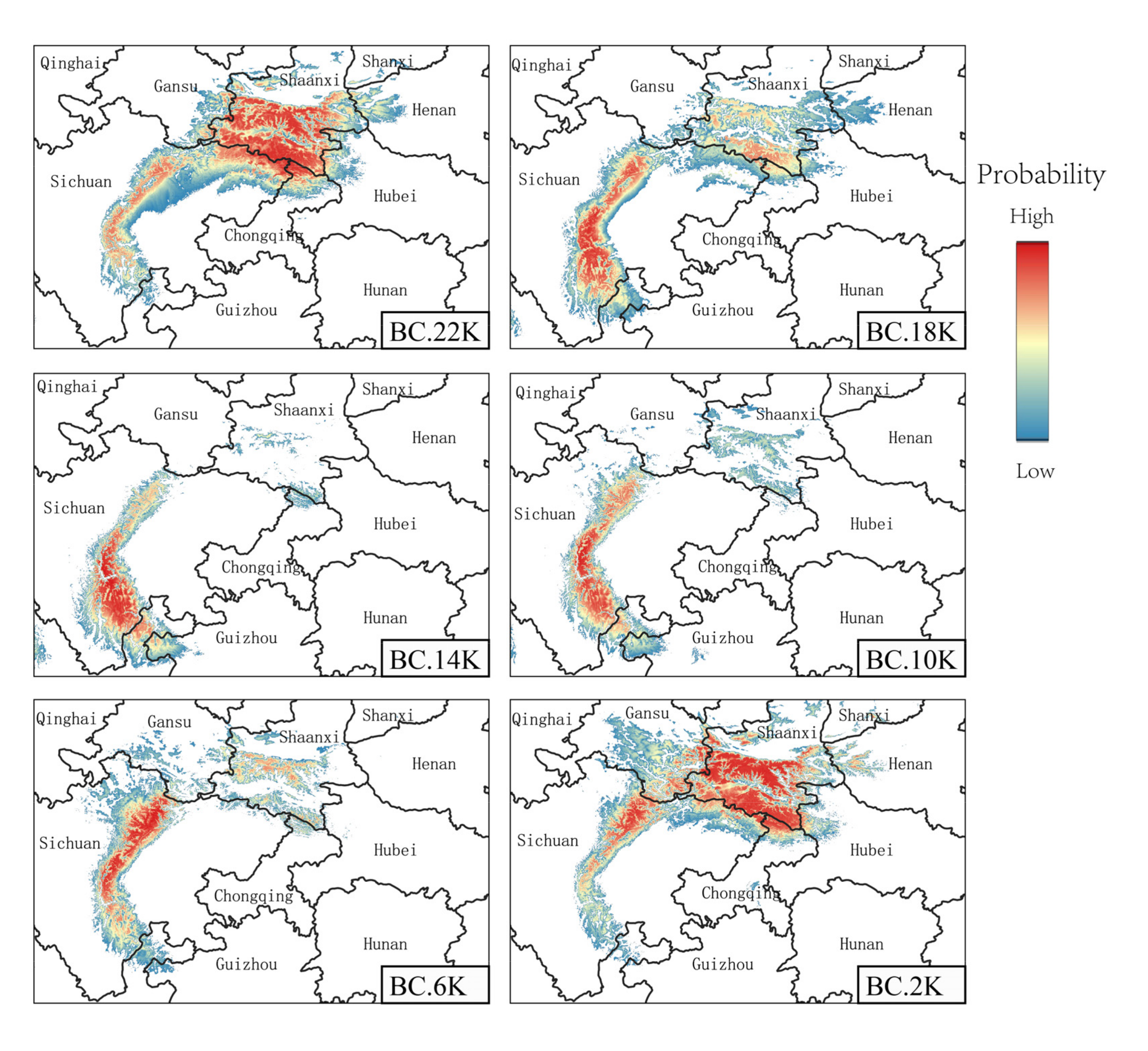
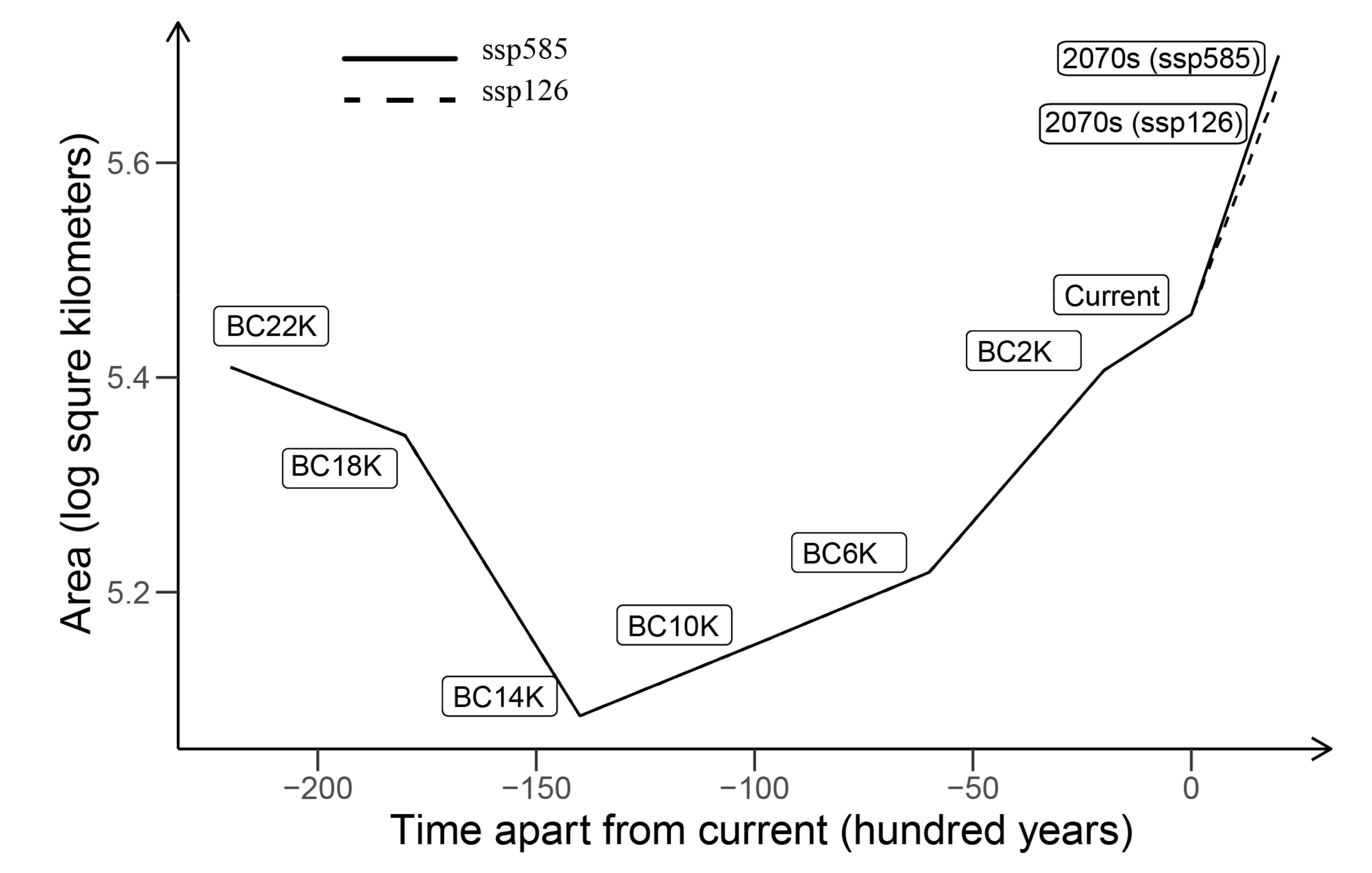
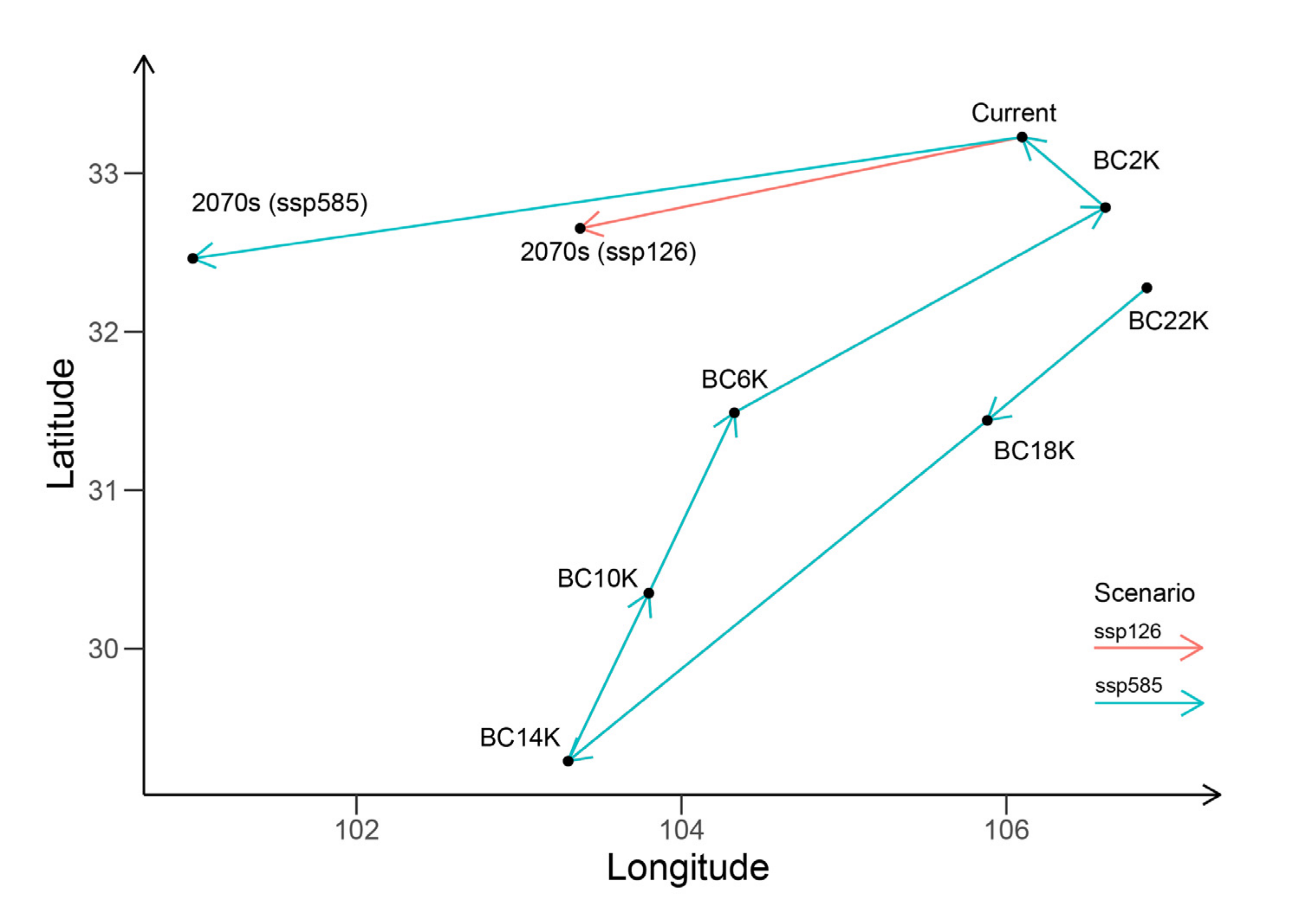
3.4. Paleo Potential Distribution and Historical Dynamics
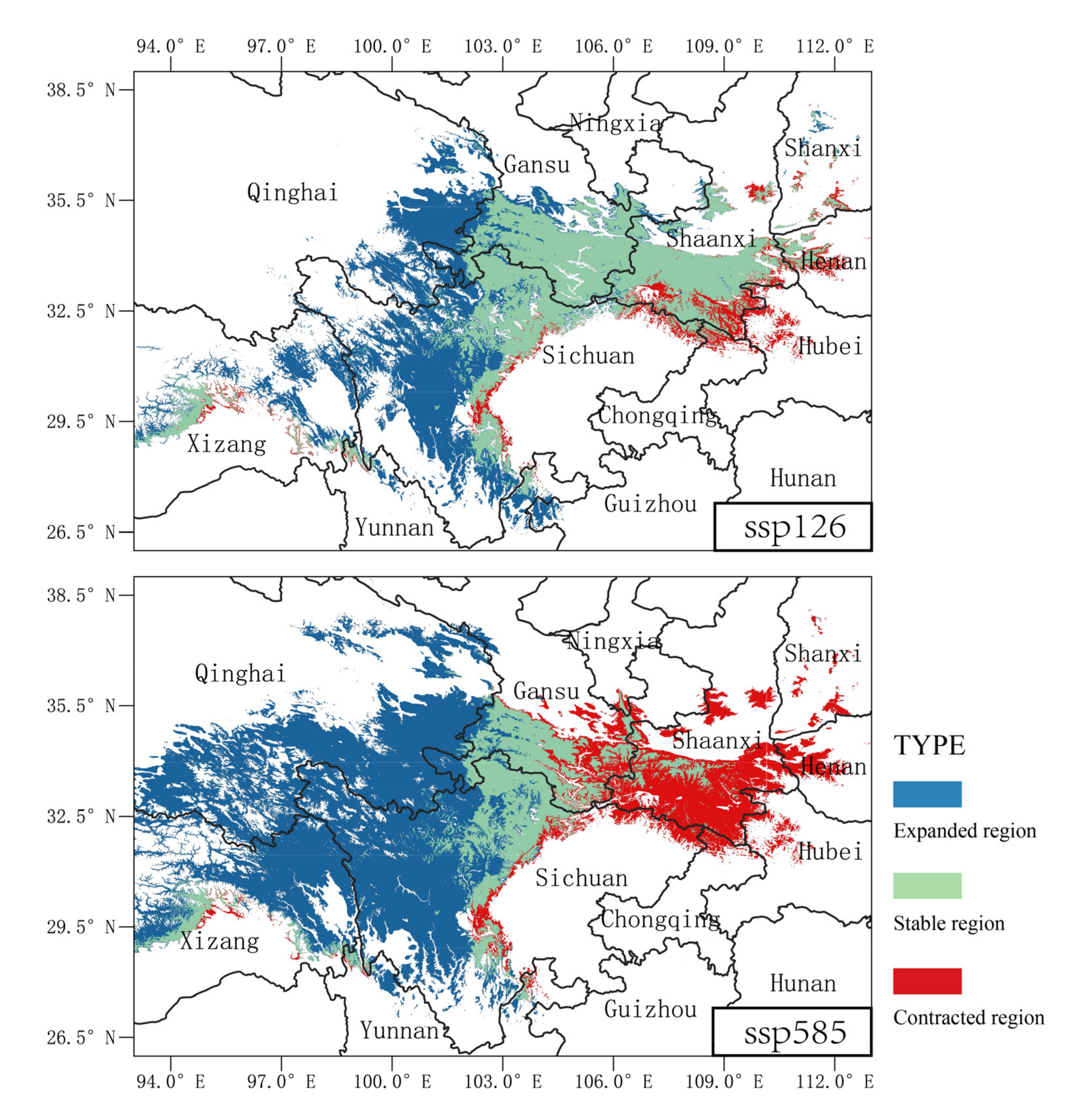
3.5. Future Potential Distribution and Ranges under Risks
4. Discussion
4.1. Climatic Niche and Proximal Variables
4.2. The Current Distribution Range and Climate Refugia
4.3. Habitat Risks under Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Michio, T. (Eds.) Flora of China; Missouri Botanical Garden Press: Saint Louis, MO, USA; Science Press: Beijing, China, 2013; Volume 6, p. 148. [Google Scholar]
- Song, H.; Ordonez, A.; Svenning, J.; Qian, H.; Yin, X.; Mao, L.; Deng, T.; Zhang, J. Regional disparity in extinction risk: Comparison of disjunct plant genera between eastern Asia and eastern North America. Glob. Change Biol. 2021, 27, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Balázs, V.L.; Filep, R.; Ambrus, T.; Kocsis, M.; Farkas, Á.; Stranczinger, S.; Papp, N. Ethnobotanical, historical and histological evaluation of Helleborus L. genetic resources used in veterinary and human ethnomedicine. Genet. Resour. Crop Evol. 2020, 67, 781–797. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Li, Y.; Zhang, Z.; Huang, W.; Zhang, H.; Jiang, Y.; Liu, J.; Zhang, D. Two New Spirostanol Glycosides from the Roots and Rhizomes of Helleborus thibetanus Franch. Rec. Nat. Prod. 2023, 17, 318–328. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Liang, X.; Song, B.; Zheng, X.; Wang, R.; Liu, L.; Song, X.; Liu, J. New cytotoxic bufadienolides from the roots and rhizomes of Helleborus thibetanus Franch. Nat. Prod. Res. 2020, 34, 950–957. [Google Scholar] [CrossRef]
- Deng, W.; Zhao, L.; Shi, X.; Du, L. Progress in the development of germplasme resource in genus Helleborus. Xiandai Hortic. 2022, 45, 1–3. (In Chinese) [Google Scholar]
- Fassou, G.; Kougioumoutzis, K.; Iatrou, G.; Trigas, P.; Papasotiropoulos, V. Genetic Diversity and Range Dynamics of Helleborus odorus subsp. cyclophyllus under Different Climate Change Scenarios. Forests 2020, 11, 620. [Google Scholar] [CrossRef]
- Záveská, E.; Kirschner, P.; Frajman, B.; Wessely, J.; Willner, W.; Gattringer, A.; Hülber, K.; Lazić, D.; Dobeš, C.; Schönswetter, P. Evidence for Glacial Refugia of the Forest Understorey Species Helleborus niger (Ranunculaceae) in the Southern as Well as in the Northern Limestone Alps. Front. Plant Sci. 2021, 12, 683043. [Google Scholar] [CrossRef]
- Sparey, M.; Cox, P.; Williamson, M.S. Bioclimatic change as a function of global warming from CMIP6 climate projections. Biogeosciences 2023, 20, 451–488. [Google Scholar] [CrossRef]
- Hoban, S.; Dawson, A.; Robinson, J.D.; Smith, A.B.; Strand, A.E. Inference of biogeographic history by formally integrating distinct lines of evidence: Genetic, environmental niche and fossil. Ecography 2019, 42, 1991–2011. [Google Scholar] [CrossRef] [Green Version]
- Mestre, F.; Barbosa, S.; Garrido-García, J.A.; Pita, R.; Mira, A.; Alves, P.C.; Paupério, J.; Searle, J.B.; Beja, P. Inferring past refugia and range dynamics through the integration of fossil, niche modelling and genomic data. J. Biogeogr. 2022, 49, 2064–2076. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Schapire, Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Smith, A.B.; Godsoe, W.; Rodriguez-Sanchez, F.; Wang, H.-H.; Warren, D. Niche Estimation Above and Below the Species Level. Trends Ecol. Evol. 2019, 34, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ci, X.; Hu, J.; Bai, Y.; Thornhill, A.H.; Conran, J.G.; Li, J. Riparian areas as a conservation priority under climate change. Sci. Total Environ. 2023, 858, 159879. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, M.; Rubolini, D.; Appukuttan, O.; Calvi, G.; Karger, D.N.; Kmecl, P.; Mihelič, T.; Sattler, T.; Seaman, B.; Teufelbauer, N.; et al. Identifying climate refugia for high-elevation Alpine birds under current climate warming predictions. Glob. Change Biol. 2022, 28, 4276–4291. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Chytrý, M.; Díaz-Varela, E.R.; Álvarez-Álvarez, P. Glacial refugia and mid-Holocene expansion delineate the current distribution of Castanea sativa in Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 491, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Li, J.; Wu, S. MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora. Forests 2022, 13, 402. [Google Scholar] [CrossRef]
- Li, Y.; Shao, W.; Huang, S.; Zhang, Y.; Fang, H.; Jiang, J. Prediction of Suitable Habitats for Sapindus delavayi Based on the MaxEnt Model. Forests 2022, 13, 1611. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Edzer, J.; Roger, S.B. Classes and methods for spatial data in R. R J. 2005, 5, 9–13. [Google Scholar]
- Robert, J.H. raster: Geographic Data Analysis and Modeling. R Package Version 2021, 2, 1–49. [Google Scholar]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.E.; Peña, L.S.; Delgado, S.G. Trayectorias Socioeconómicas Compartidas (SSP): Nuevas maneras de comprender el cambio climático y social. Estud. Demográficos Urbanos 2017, 32, 669–693. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Nobis, M.P.; Normand, S.; Graham, C.H.; Zimmermann, N.E. CHELSA-TraCE21k v1.0. Downscaled transient temperature and precipitation data since the last glacial maximum. Clim. Past Discuss. 2021, 1, 1–27. [Google Scholar]
- Naimi, B.; Hamm, N.A.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Trevor, H.; Robert, T.; Jerome, F. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A. Want to model a species niche? A step-by-step guideline on correlative ecological niche modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Elith, J.; Ferrier, S.; Huettmann, F.; Leathwick, J. The evaluation strip: A new and robust method for plotting predicted responses from species distribution models. Ecol. Model. 2005, 186, 280–289. [Google Scholar] [CrossRef]
- Van Der Wal, J.; Falconi, L.; Januchowski, S.; Shoo, L.; Storlie, C. SDMTools: Species Distribution Modelling Tools: Tools for processing data associated with species distribution modelling exercises. R Package Version 2014, 1, 1–221. [Google Scholar]
- Hadley, W. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; p. 260. [Google Scholar]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Sayre, R.; Trabucco, A.; Zomer, R. A high-resolution bioclimate map of the world: A unifying framework for global biodiversity research and monitoring. Glob. Ecol. Biogeogr. 2013, 22, 630–638. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shan, X.; Ovando, D.; Yang, T.; Dai, F.; Jin, X. Predicting current and future global distribution of black rockfish (Sebastes schlegelii) under changing climate. Ecol. Indic. 2021, 128, 107799. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, D.; Codella, S.G. Seed re-dispersal of four myrmecochorous plants by a keystone ant in central China. Ecol. Res. 2017, 32, 387–393. [Google Scholar] [CrossRef]
- Chen, Q.; Yin, Y.; Zhao, R.; Yang, Y.; Da Silva, J.A.T.; Yu, X. Incorporating Local Adaptation into Species Distribution Modeling of Paeonia mairei, an Endemic Plant to China. Front. Plant Sci. 2019, 10, 1717–1731. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Bioclimatic Variables | Unit | Mean Importance | Standard Deviation |
|---|---|---|---|---|
| bio01 | Mean temperature | °C | - | - |
| bio02 | Diurnal air temperature range | °C | - | - |
| bio03 | Isothermality (bio02/bio07 ×100) | \ | 2.20 | 0.08 |
| bio04 | Temperature seasonality (standard deviation ×100) | °C | 15.06 | 0.40 |
| bio05 | Max temperature of warm month | °C | - | - |
| bio06 | Min temperature of cold month | °C | - | - |
| bio07 | Annual temperature range | °C | - | - |
| bio08 | Mean temperature of wet quarter | °C | 23.41 | 0.60 |
| bio09 | Mean temperature of dry quarter | °C | - | - |
| bio10 | Mean temperature of warm quarter | °C | - | - |
| bio11 | Mean temperature of cold quarter | °C | - | - |
| bio12 | Annual precipitation | mm | - | - |
| bio13 | Precipitation of wet month | mm | - | - |
| bio14 | Precipitation of dry month | mm | - | - |
| bio15 | Precipitation seasonality (coefficient of variation) | \ | 11.56 | 0.37 |
| bio16 | Precipitation of wet quarter | mm | - | - |
| bio17 | Precipitation of dry quarter | mm | - | - |
| bio18 | Precipitation of warm quarter | mm | 14.68 | 0.46 |
| bio19 | Precipitation of cold quarter | mm | 33.09 | 0.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Mao, L.; Sun, M.; Ma, G.; Zhu, K. Paleo Distribution and Habitat Risks under Climate Change of Helleborus thibetanus. Forests 2023, 14, 630. https://doi.org/10.3390/f14030630
Shi X, Mao L, Sun M, Ma G, Zhu K. Paleo Distribution and Habitat Risks under Climate Change of Helleborus thibetanus. Forests. 2023; 14(3):630. https://doi.org/10.3390/f14030630
Chicago/Turabian StyleShi, Xiaohua, Lihui Mao, Miao Sun, Guangying Ma, and Kaiyuan Zhu. 2023. "Paleo Distribution and Habitat Risks under Climate Change of Helleborus thibetanus" Forests 14, no. 3: 630. https://doi.org/10.3390/f14030630
APA StyleShi, X., Mao, L., Sun, M., Ma, G., & Zhu, K. (2023). Paleo Distribution and Habitat Risks under Climate Change of Helleborus thibetanus. Forests, 14(3), 630. https://doi.org/10.3390/f14030630