



Rockrose Land Management: Contribution of Periodic Harvesting to Increase Value and to Control Cistus ladanifer L. Shrublands

 , , and
, , and 
Abstract
:1. Introduction
- Dense shrublands (248,382 and 460,088 ha in Portugal and Spain, respectively): most likely abandoned areas with no management and profit with a high probability of shrubland perpetuation due to cyclic fire events. (Cover index of 0.85 in Godinho-Ferreira et al. [15] and of 0.68 in Montero et al. [16] both considering the whole vegetation but with a dominance by C. ladanifer).
- Shrublands associated to oak, pine, or eucalyptus forests (1,783,666 and 2,450,857 ha in Portugal and Spain, respectively): most likely managed areas because of the expected profit (e.g., periodic total clearings of the shrub layer). Open oak woodlands would include the “montado” agro–forestry–pasture system.
- Shrublands associated to a diverse forest (1,111,669 ha in Portugal): most likely poorly managed and with no profit and some probability of dense C. ladanifer shrublands reclamation after a fire event.
2. Materials and Methods
2.1. Characterisation of the Study Area
2.2. Delimitation and Evaluation of Experimental Plots
2.3. Soil Sampling and Analysis
2.4. Experimental Design
2.5. Harvesting and Sampling
2.6. Labdanum Resin Extraction
2.7. Biomass Water Content
2.8. Statistical Analysis
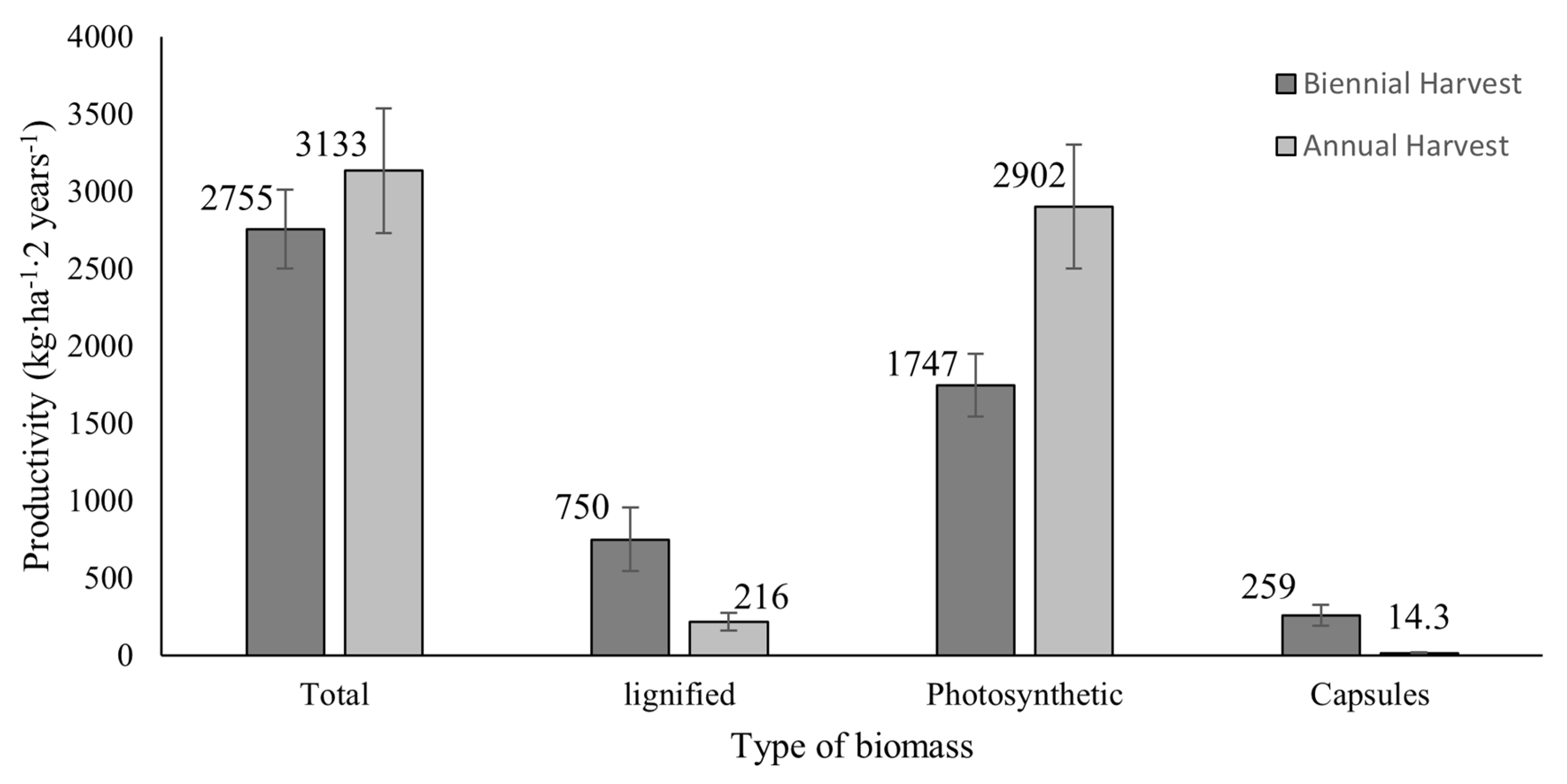
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Demoly, J.; Montserrat, P. Cistus. In Flora Ibérica; Castroviejo, S., Ed.; CSIC (Centro Superior de Investigaciones Científicas): Madrid, Spain, 1993; pp. 319–337. [Google Scholar]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands–Portugal case study. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2015, 149, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Oria-de-Rueda, J.A.; Martín-Pinto, P.; Olaizola, J. Bolete Productivity of Cistaceous Scrublands in Northwestern Spain. Econ. Bot. 2008, 62, 323–330. [Google Scholar] [CrossRef]
- Trabaud, L. Modalités de germination des cistes et des pins méditerranéens et colonisation des sites perturbés. Rev. D’écol. 1995, 50, 3–14. Available online: https://hal.science/hal-03529132 (accessed on 20 February 2023). [CrossRef]
- Alonso, I.; Luis, E.; Tarrega, R. First phases of regeneration of Cistus laurifolius and Cistus ladanifer after burning and cutting in experimental plots. Int. J. Wildland Fire 1992, 2, 7–14. [Google Scholar] [CrossRef]
- Tárrega, R.; Luis-Calabuig, E.; Alonso, I. Comparison of the regeneration after burning, cutting and ploughing in a Cistus ladanifer shrubland. Vegetatio 1995, 120, 59–67. [Google Scholar] [CrossRef]
- Luiscalabuig, E.; Tárrega, R.; Alonso, I. Seedling regeneration of two Cistus species after experimental disturbances. Int. J. Wildland Fire 1996, 6, 13–19. [Google Scholar] [CrossRef]
- Tárrega, R.; Luis-Calabuig, E.; Valbuena, L. Eleven years of recovery dynamic after experimental burning and cutting in two Cistus communities. Acta Oecol. 2001, 22, 277–283. [Google Scholar] [CrossRef]
- Frazão, D.F.; Raimundo, J.R.; Domingues, J.L.; Quintela-Sabarís, C.; Gonçalves, J.C.; Delgado, F. Cistus ladanifer (Cistaceae): A natural resource in Mediterranean-type ecosystems. Planta 2018, 247, 289–300. [Google Scholar] [CrossRef]
- Simões, M.P.; Madeira, M.; Gazarini, L. Ability of Cistus L. shrubs to promote soil rehabilitation in extensive oak woodlands of Mediterranean areas. Plant Soil 2009, 323, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Peinado, R.; Moreno, G.; Juarez, E.; Montero, G.; Roig, S. The contribution of two common shrub species to aboveground and belowground carbon stock in Iberian dehesas. J. Arid. Environ. 2013, 91, 22–30. [Google Scholar] [CrossRef]
- Alías, J.C.G.; García, M.; Sosa, T.; Valares, C.; Chaves, N. Carbon storage in the different compartments of two systems of shrubs of the southwestern Iberian Peninsula. Agrofor. Syst. 2015, 89, 575–585. [Google Scholar] [CrossRef]
- Carrión-Prieto, P.; Hernández-Navarro, S.; Martín-Ramos, P.; Sánchez-Sastre, L.; Garrido-Laurnaga, F.; Marcos-Robles, J.; Martín-Gil, J. Mediterranean shrublands as carbon sinks for climate change mitigation: New root-to-shoot ratios. Carbon Manag. 2017, 8, 67–77. [Google Scholar] [CrossRef]
- Raimundo, J.R.; Frazão, D.F.; Domingues, J.L.; Quintela-Sabarís, C.; Dentinho, T.P.; Anjos, O.; Alves, M.; Delgado, F. Neglected Mediterranean plant species are valuable resources: The example of Cistus ladanifer. Planta 2018, 248, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Godinho-Ferreira, P.; Azevedo, A.; Rego, F. Carta da tipologia florestal de Portugal Continental. Silva Lusit. 2005, 13, 1–34. [Google Scholar]
- Montero, G.; López-Leiva, C.; Ruiz-Peinado, R.; López-Senespleda, E.; Onrubia, R.; Pasalodos, M. Producción De Biomasa Y Fijación De Carbono Por Los Matorrales Españoles Y Por El Horizonte Orgánico Superficial De Los Suelos Forestales; Ministerio De Agricultura, Pesca Y Alimentación, Secretaría General Técnica: Madrid, Spain, 2020.
- Hernández-Rodríguez, M.; Martín-Pinto, P.; Oria-de-Rueda, J.A.; Diaz-Balteiro, L. Optimal management of Cistus ladanifer shrublands for biomass and Boletus edulis mushroom production. Agrofor. Syst. 2017, 91, 663–676. [Google Scholar] [CrossRef]
- Carrión-Prieto, P.; Martín-Ramos, P.; Hernández-Navarro, S.; Sánchez-Sastre, L.F.; Marcos-Robles, J.L.; Martín-Gil, J. Valorization of Cistus ladanifer and Erica arborea shrubs for fuel: Wood and bark thermal characterization. Maderas Cienc. Y Tecnol. 2017, 19, 443–454. [Google Scholar] [CrossRef]
- Encinar, J.M.; González, J.F.; Nogales-Delgado, S. Catalyzed steam gasification of Cistus ladanifer biochar. Catalysts 2020, 10, 1430. [Google Scholar] [CrossRef]
- Ferro, M.D.; Fernandes, M.C.; Paulino, A.F.; Prozil, S.O.; Gravitis, J.; Evtuguin, D.V.; Xavier, A.M. Bioethanol production from steam explosion pretreated and alkali extracted Cistus ladanifer (rockrose). Biochem. Eng. J. 2015, 104, 98–105. [Google Scholar] [CrossRef]
- Alves-Ferreira, J.; Lourenço, A.; Morgado, F.; Duarte, L.C.; Roseiro, L.B.; Fernandes, M.C.; Pereira, H.; Carvalheiro, F. Delignification of Cistus ladanifer biomass by organosolv and alkali processes. Energies 2021, 14, 1127. [Google Scholar] [CrossRef]
- Alves-Ferreira, J.; Carvalheiro, F.; Duarte, L.C.; Ferreira, A.R.; Martinez, A.; Pereira, H.; Fernandes, M.C. D-Lactic acid production from Cistus ladanifer residues: Co-fermentation of pentoses and hexoses by Escherichia coli JU15. Ind. Crops Prod. 2022, 177, 114519. [Google Scholar] [CrossRef]
- Carrión-Prieto, P.; Martín-Ramos, P.; Hernández-Navarro, S.; Sánchez-Sastre, L.F.; Marcos-Robles, J.L.; Martín-Gil, J. Crystallinity of cellulose microfibers derived from Cistus ladanifer and Erica arborea shrubs. Maderas Cienc. Tecnol. 2019, 21, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Jerónimo, E.; Alfaia, C.M.; Alves, S.P.; Dentinho, M.T.; Prates, J.A.; Vasta, V.; Santos-Silva, J.; Bessa, R.J. Effect of dietary grape seed extract and Cistus ladanifer L. in combination with vegetable oil supplementation on lamb meat quality. Meat Sci. 2012, 92, 841–847. [Google Scholar] [CrossRef]
- Dentinho, M.; Belo, A.; Bessa, R. Digestion, ruminal fermentation and microbial nitrogen supply in sheep fed soybean meal treated with Cistus ladanifer L. tannins. Small Rumin. Res. 2014, 119, 57–64. [Google Scholar] [CrossRef]
- Francisco, A.; Dentinho, M.; Alves, S.; Portugal, P.; Fernandes, F.; Sengo, S.; Jerónimo, E.; Oliveira, M.; Costa, P.; Sequeira, A. Growth performance, carcass and meat quality of lambs supplemented with increasing levels of a tanniferous bush (Cistus ladanifer L.) and vegetable oils. Meat Sci. 2015, 100, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Frazão, D.F.; Paulo, L.; Peres, F.; Resende, M.; Santo, C.E.; Barroca, C.; Moitinho, A.; Delgado, F. Cistus ladanifer seeds: From ancient snack to novel and sustainable food ingredient. J. Food Compos. Anal. 2022, 109, 104503. [Google Scholar] [CrossRef]
- El Farissi, H.; Lakhmiri, R.; El Fargani, H.; Albourine, A.; Safi, M. Production of bio-oil from Cistus ladanifer shell by fixed-bed pyrolysis. Int. J. Eng. Technol. 2017, 17, 25–30. [Google Scholar]
- Rodrigues, A.M.; Paulo, L.; Franco Frazão, D.; Beato, H.; Resende, M.; Delgado, F. Chemical Composition and Fatty Acid Profile of Cistus ladanifer L. Capsules for Animal Feed. In International Conference on Multidisciplinary Research; Millenium–Journal of Education, Technologies and Health: Viseu, Portugal, 2021. [Google Scholar] [CrossRef]
- Sosa, T.; Alías, J.C.; Escudero, J.C.; Chaves, N. Interpopulational variation in the flavonoid composition of Cistus ladanifer L. exudate. Biochem. Syst. Ecol. 2005, 33, 353–364. [Google Scholar] [CrossRef]
- Frazão, D.F.; Martins-Gomes, C.; Steck, J.L.; Keller, J.; Delgado, F.; Gonçalves, J.C.; Bunzel, M.; Pintado, C.M.; Díaz, T.S.; Silva, A.M. Labdanum Resin from Cistus ladanifer L.: A Natural and Sustainable Ingredient for Skin Care Cosmetics with Relevant Cosmeceutical Bioactivities. Plants 2022, 11, 1477. [Google Scholar] [CrossRef] [PubMed]
- Agroconsultores, G. Elaboraçao da Carta de Solos e de Aptidao das Terras da Zona Interior Centro (Making of the Soil Map and Land Suitability of Zona Interior Centro); Instituto de Desenvolvimento Rural e Hidráulica: Lisbon, Portugal, 2004.
- Monteiro, M. Normal Climatológica 1986–2015. Posto Meteorológico da Escola Superior Agrária de Castelo Branco; IPCB: Castelo Branco, Portugal, 2016; Available online: http://hdl.handle.net/10400.11/5406 (accessed on 20 February 2023).
- Costa, J.C.; Aguiar, C.; Capelo, J.; Lousã, M.; Neto, C. Biogeografia de Portugal Continental; Quercetea: Lisbon, Portugal, 1998; Available online: http://hdl.handle.net/10198/714 (accessed on 20 February 2023).
- Castro, H.; Freitas, H. Above-ground biomass and productivity in the Montado: From herbaceous to shrub dominated communities. J. Arid. Environ. 2009, 73, 506–511. [Google Scholar] [CrossRef] [Green Version]
- Morgado, J.M.; Tapias, R.; Alesso, P. Producción de Goma Bruta de Jara (Cistus ladanifer L.) en el Suroeste de la Península Ibérica. In IV Congreso Forestal Español; Escuela Politécnica Superior: Zaragoza, Spain, 2005. [Google Scholar]
- Burguer, L. Investigação e Comparação de Metodologias de Extração de Lábdano Obtido a Partir de Cistus ladanifer L. Master’s Thesis, Instituto Politécnico de Bragança, Universidadad de Salamanca, Bragança, Portugal, 2016. Available online: http://hdl.handle.net/10198/13662 (accessed on 20 February 2023).
- Chaves, N.; Escudero, J.C.; Gutierrez-Merino, C. Role of ecological variables in the seasonal variation of flavonoid content of Cistus ladanifer exudate. J. Chem. Ecol. 1997, 23, 579–603. [Google Scholar] [CrossRef]
- Alías, J.C.G.; Sosa, T.; Valares, C.; Escudero, J.C.; Chaves, N. Seasonal variation of Cistus ladanifer L. diterpenes. Plants 2012, 1, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Talavera, S.; Gibbs, P.; Herrera, J. Reproductive biology of Cistus ladanifer (Cistaceae). Plant Syst. Evol. 1993, 186, 123–134. [Google Scholar] [CrossRef]
- Simões, M.P.; Madeira, M.; Gazarini, L. The role of phenology, growth and nutrient retention during leaf fall in the competitive potential of two species of Mediterranean shrubs in the context of global climate changes. Flora Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 578–589. [Google Scholar] [CrossRef]
- Nuñez, E.; Cabeza, J.; Escudero, J. Relación entre la biomasa de jarales y su rendimiento energetico por pirolisis. Opt. Méditerr. 1989, 3, 345–350. Available online: http://om.ciheam.org/article.php?IDPDF=CI000565 (accessed on 20 February 2023).
- Alves-Ferreira, J.; Duarte, L.C.; Fernandes, M.C.; Pereira, H.; Carvalheiro, F. Hydrothermal treatments of Cistus ladanifer industrial residues obtained from essential oil distilleries. Waste Biomass Valoriz. 2019, 10, 1303–1310. [Google Scholar] [CrossRef]
- Francisco, M.; David, G.; Francisco, M. Notes of Changes in Biodiversity in the Exploited Populations of Cistus Ladanifer L.;(Cistaceae) from SW Iberian Peninsula. Biodivers. Online J. 2022, 2, BOJ.000540.2022. [Google Scholar]
- Vallejo, V.R.; Alloza, J.A. Shrubland and woodland restoration in the mediterranean basin. In Proceedings of the Chaparral Restoration Workshop, General Technical Report PSW-GTR-265, Albany, CA, USA, 2019; pp. 51–65. Available online: https://www.fs.usda.gov/psw/publications/documents/psw_gtr265/psw_gtr265_051.pdf (accessed on 20 February 2023).


{kind=link}
| Parameters | Values | |
|---|---|---|
| Texture class (%) | Sand | 56.4 |
| Silt | 24.7 | |
| Clay | 18.9 | |
| OM | 2.36 | |
| Available phosphorus (P2O5 mg∙kg−1) | 7 | |
| Available potassium (K2O mg∙kg−1) | 99 | |
| Exchangeable bases (mg∙kg−1) | Calcium | 280 |
| Magnesium | 51.4 | |
| Potassium | 57.4 | |
| Sodium | 8.86 | |
| pH (H2O) | 5.4 | |
| Mean Plant Height (m) | Mean Plant Cover Area (m2) | Plant Density (Plants∙m−2) | Cover (%) |
|---|---|---|---|
| 1.10 ± 0.29 | 0.872 ± 0.730 | 1.62 ± 1.17 | 76.9 ± 28.9 |
| Season | ||||
|---|---|---|---|---|
| ES | MS | LS | ||
| Resin yield (% dw/fw) | ||||
| Year | 2020 | 6.02 ± 0.82 | 7.49 ± 0.71 | 7.35 ± 0.65 |
| 2021 | 6.48 ± 0.41 | 6.11 ± 0.91 | 7.10 ± 0.39 | |
| Resin productivity (kg∙ha−1∙2 years−1) | ||||
| Periodicity | Biennial | 89.9 ± 11.8 | 119 ± 25 | 134 ± 20 |
| Annual (*) | 169 ± 33 | 181 ± 36 | 230 ± 50 | |
| Resin productivity (kg∙ha−1∙year−1) | ||||
| (*)Year | 2020 | 81.4 ± 19.5 | 89.0 ± 14.2 | 97.0 ± 40.7 |
| 2021 | 87.8 ± 14.0 | 92.3 ± 22.5 | 133 ± 23 | |
| Season | |||
|---|---|---|---|
| ES | MS | LS | |
| Biennial Periodicity | Seeds yield (% fw∙fw−1) | ||
| 33.1 ± 3.3 | 25.2 ± 5.1 | 28.2 ± 1.8 | |
| Seeds productivity (kg∙ha−1∙2 years−1) | |||
| 103 ± 52 | 81.1 ± 36.6 | 49.3 ± 9.9 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frazão, D.F.; Gonçalves, J.C.; Silva, A.M.; Delgado, F. Rockrose Land Management: Contribution of Periodic Harvesting to Increase Value and to Control Cistus ladanifer L. Shrublands. Forests 2023, 14, 638. https://doi.org/10.3390/f14030638
Frazão DF, Gonçalves JC, Silva AM, Delgado F. Rockrose Land Management: Contribution of Periodic Harvesting to Increase Value and to Control Cistus ladanifer L. Shrublands. Forests. 2023; 14(3):638. https://doi.org/10.3390/f14030638
Chicago/Turabian StyleFrazão, David Franco, José Carlos Gonçalves, Amélia M. Silva, and Fernanda Delgado. 2023. "Rockrose Land Management: Contribution of Periodic Harvesting to Increase Value and to Control Cistus ladanifer L. Shrublands" Forests 14, no. 3: 638. https://doi.org/10.3390/f14030638
APA StyleFrazão, D. F., Gonçalves, J. C., Silva, A. M., & Delgado, F. (2023). Rockrose Land Management: Contribution of Periodic Harvesting to Increase Value and to Control Cistus ladanifer L. Shrublands. Forests, 14(3), 638. https://doi.org/10.3390/f14030638