

Estimation of Biomass and Carbon Sequestration Potential of Dalbergia latifolia Roxb. and Melia composita Willd. Plantations in the Tarai Region (India)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
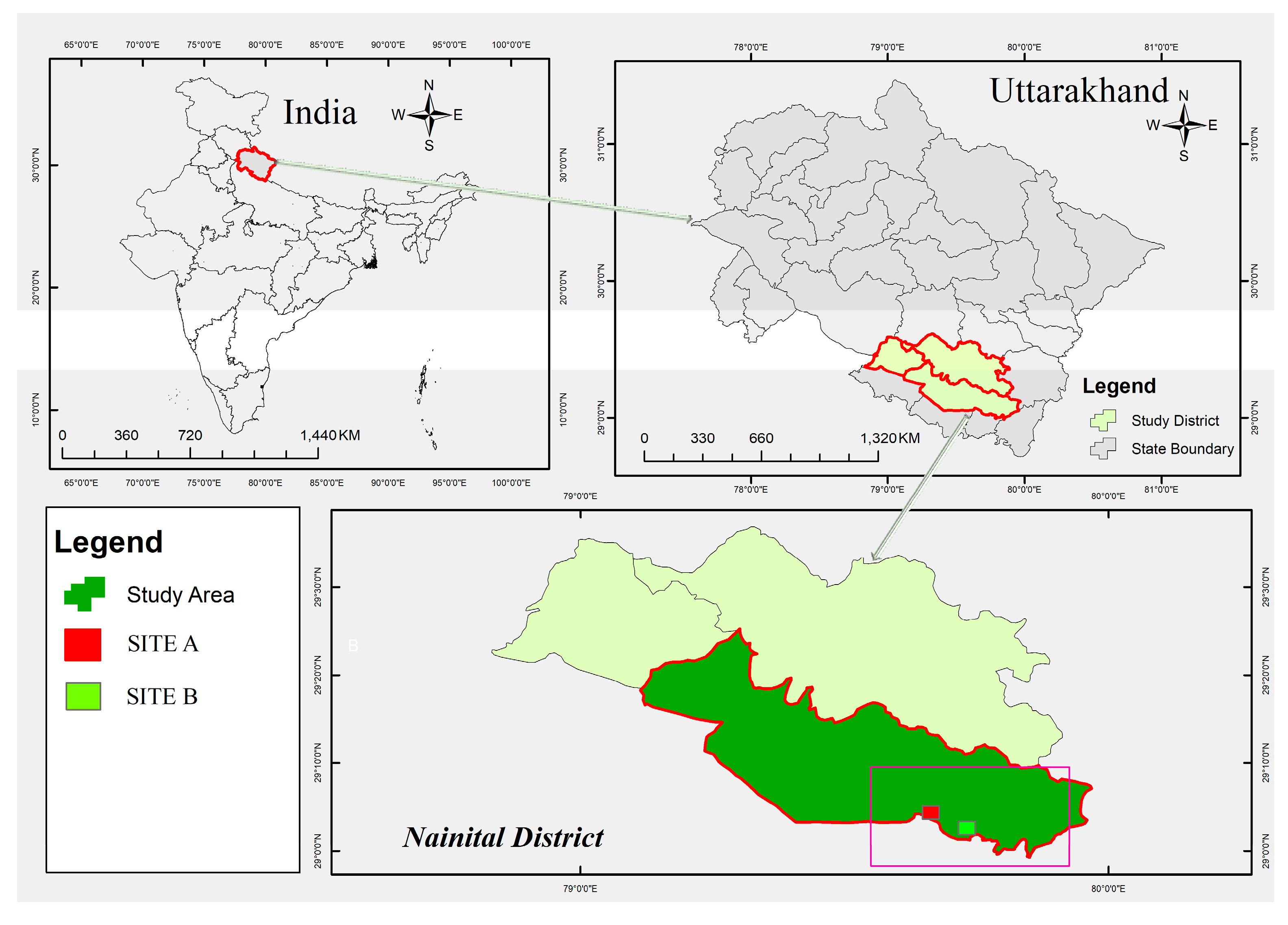
2.1. Study Area
2.2. Methods
2.2.1. Physicochemical Analysis of Soil
2.2.2. Phytosociological Analysis
2.2.3. Development of Regression Equations
2.2.4. Tree Biomass Accumulation
2.2.5. Understory Vegetation Biomass
2.2.6. Forest Floor Biomass and Litter Inputs
2.2.7. Carbon Accumulation and Sequestration
2.2.8. Statistical Analysis
3. Results
3.1. Stand Structure and Physicochemical Characteristics of Soil
3.2. Regression Equation Relationship
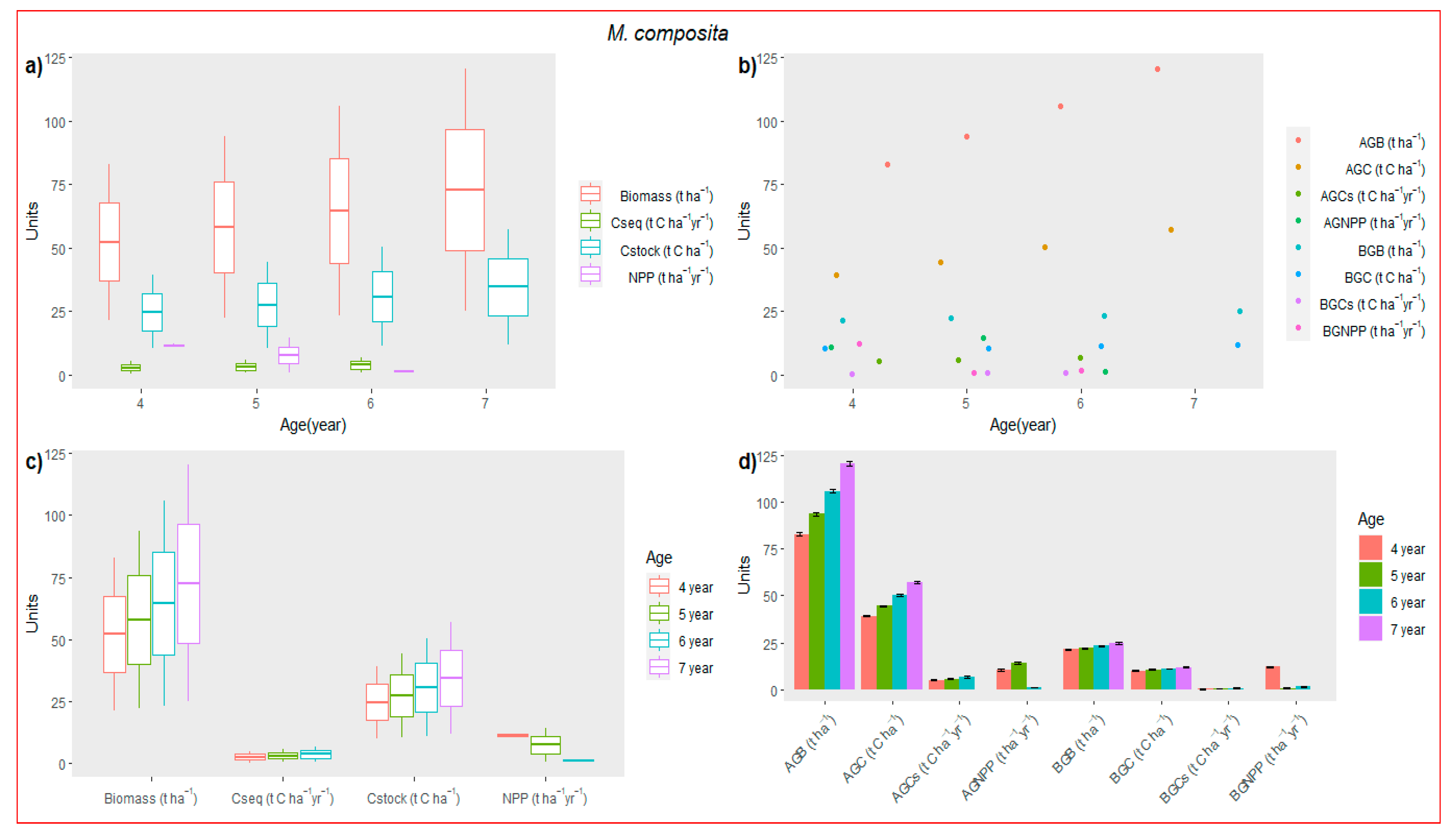
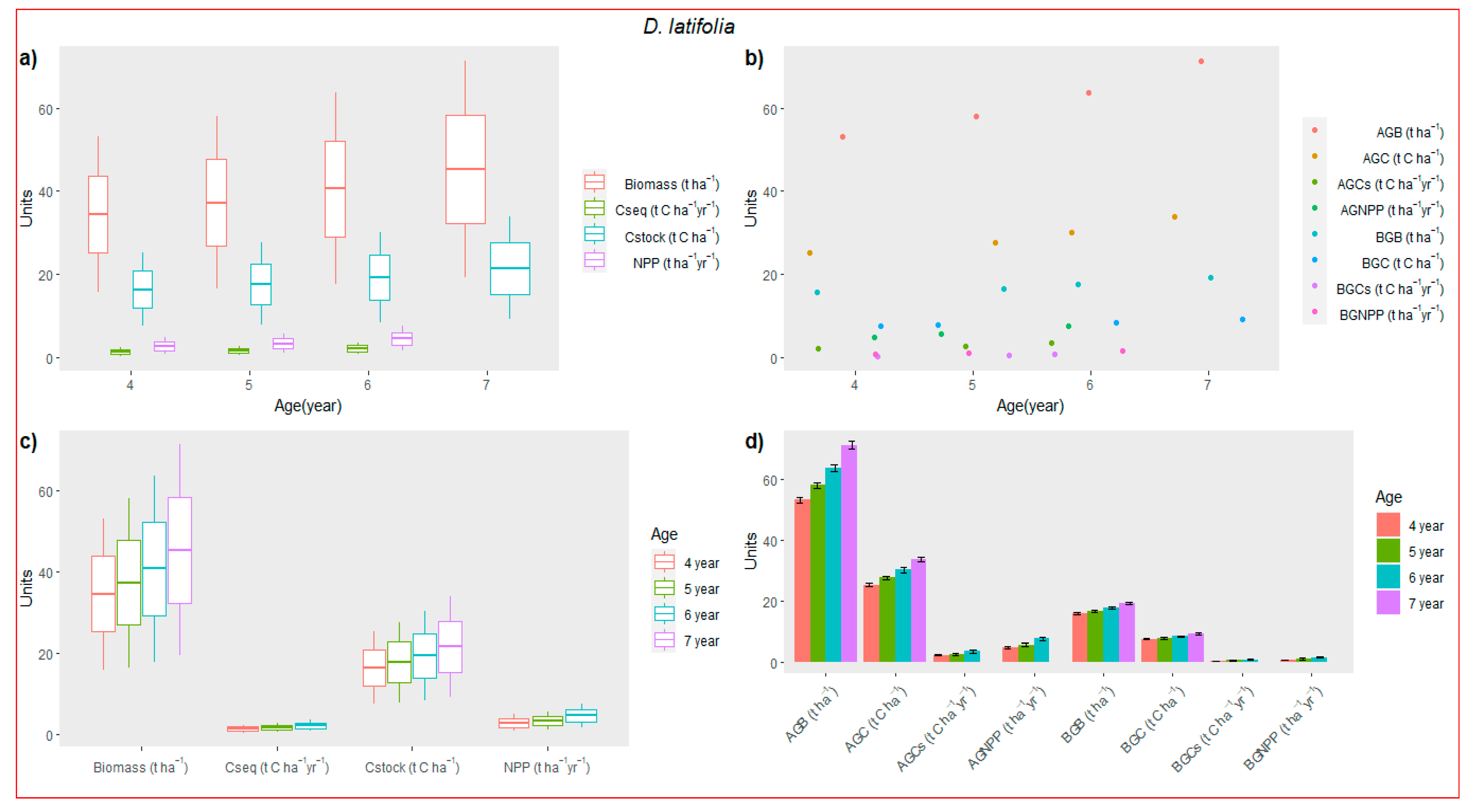
3.3. Biomass Accumulation
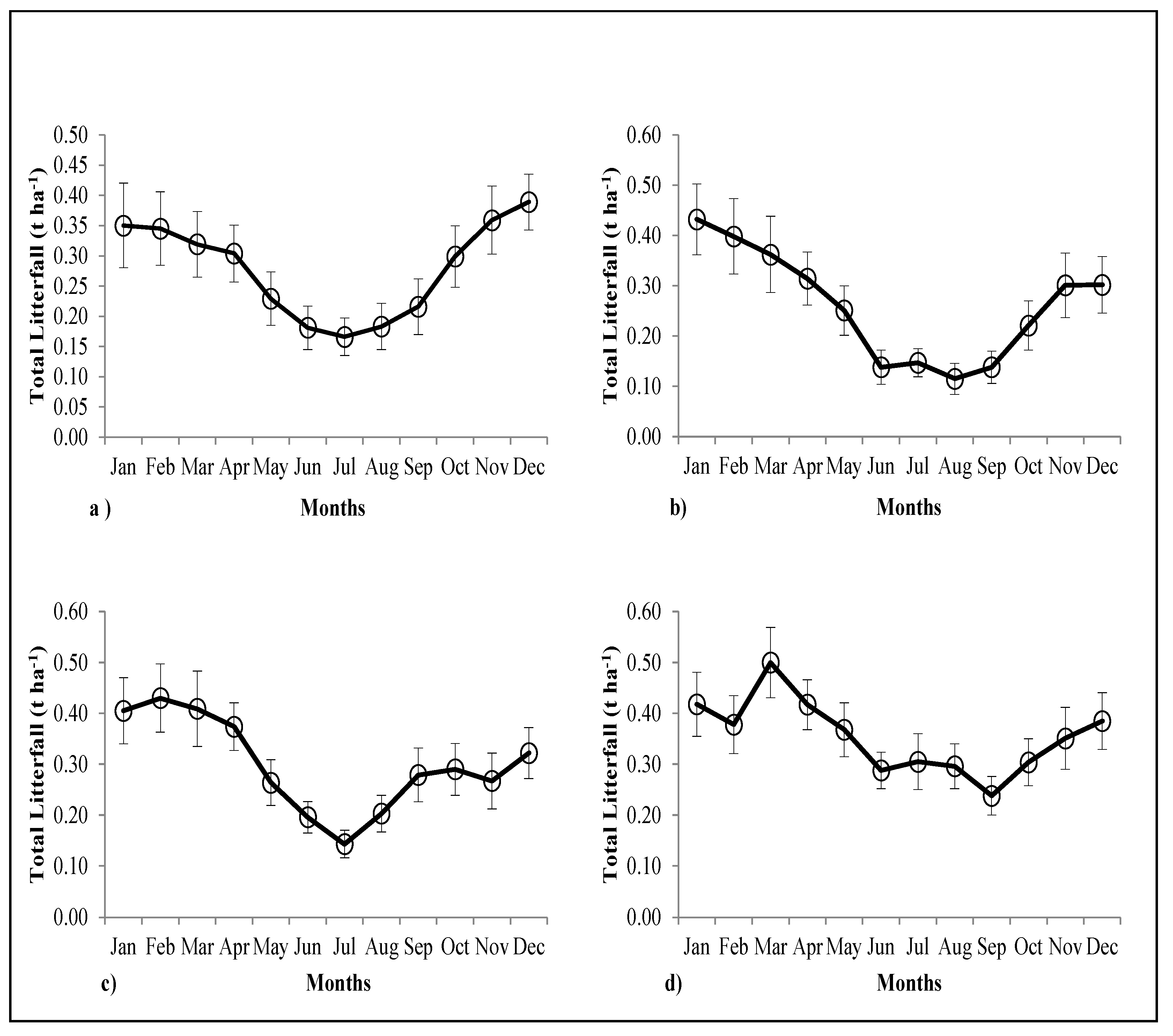
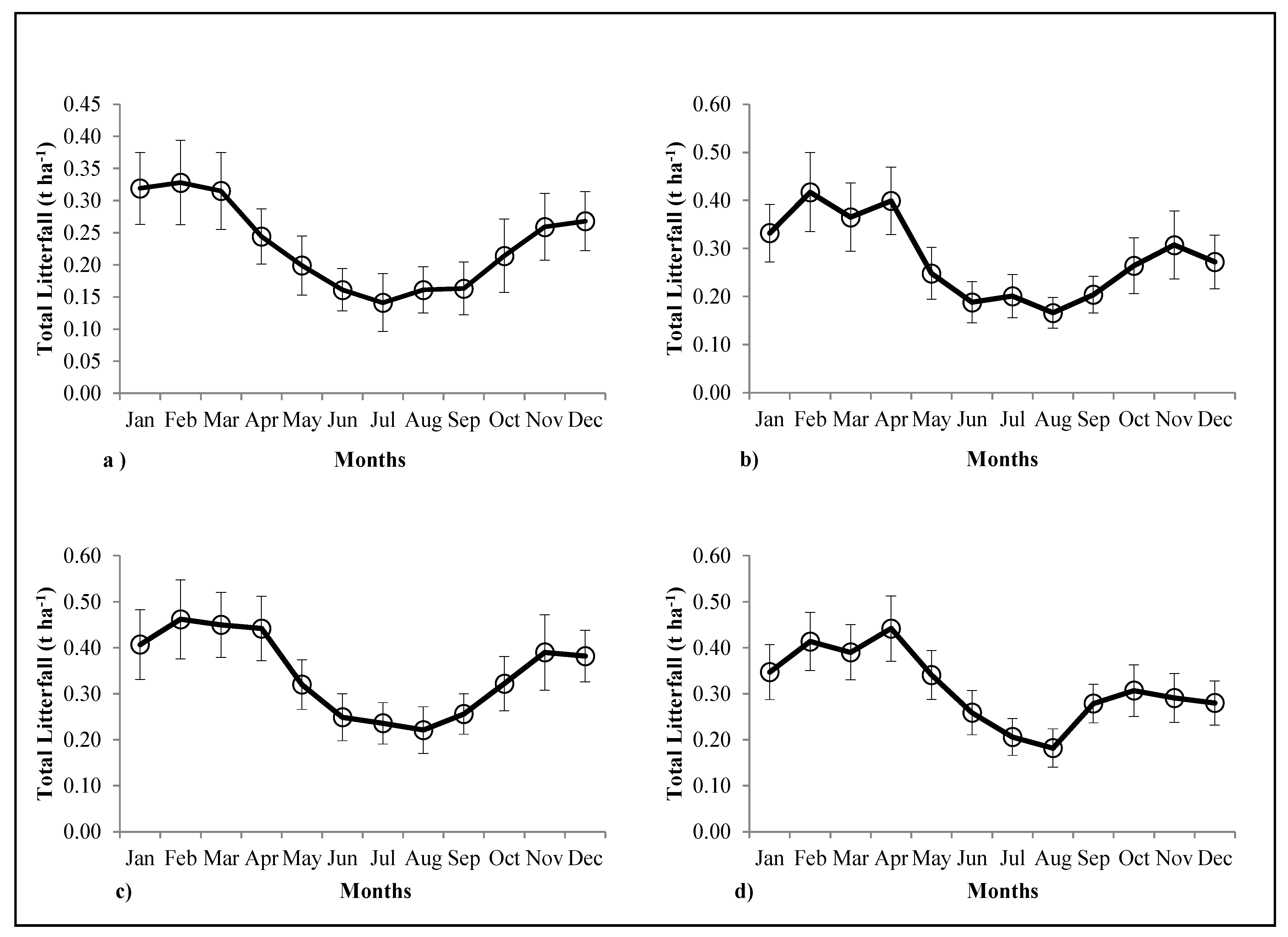
3.4. Forest Floor Biomass and Litter Inputs
3.5. Carbon Accumulation and Sequestration (Cseq)
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Species | Age (Years) | Density (Ind. ha−1) | Carbon Stock (t Cha−1) | C Seq. (t Cha−1yr−1) | Reference |
|---|---|---|---|---|---|---|
| India | Teak forest | 4 | 3490 | 12.61 | 12.80 | Karmacharya and Singh [68] |
| India | Poplar | 9 | 400 | 55 | 8 | Kaul et al. [69] |
| India | Eucalyptus | 9 | 2000 | 41 | 6 | Kaul et al. [69] |
| India | Mixed plantation (D. sissoo, A. catechu and A. lebbeck) | 5 | 1322 | 0.11 | 0.12 | Singh et al. [66] |
| India | Dry tropical | - | - | - | 2.4 | Chaturvedi et al. [70] |
| India | P. deltoides | 8 | 500 | 28.67 | 2.75 | Kanime et al. [60] |
| India | E. tereticornis | 10 | 120 | 10.52 | 0.84 | |
| India | D. sissoo | 10 | 1666 | 43.39 | 2.73 | |
| India India | Teak Teak | 5 11 | - - | 15.8 35.4 | 6.96 5.46 | Jha [30] |
| India | Teak plantation | - | - | 230.16 | Singh et al. [71] | |
| India | Areca catechu | - | 1320 | 36.48 | Dabi et al. [72] | |
| India | M. composita | 4–7 | 960 | 49.54–68.94 | 5.46–7.60 | Present study |
| India | D. latifolia | 4–7 | 880 | 32.71–42.95 | 2.66–4.33 | Present study |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goulder, L.; Pizer, W. The Economics of Climate Change; Discussion Paper RFF DP 06-06; Resources for the Future: Washington, DC, USA, 2006. [Google Scholar]
- Archer, D.; Brovkin, V. The millennial atmospheric lifetime of anthropogenic CO2. Clim. Chang. 2008, 90, 283–297. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Pandey, S.S.; Maraseni, T.N.; Cockfield, G. Carbon stock dynamics in different vegetation dominated community forests under REDD+: A case from Nepal. For. Ecol. Manag. 2014, 327, 40–47. [Google Scholar] [CrossRef]
- Yadav, R.P.; Bisht, J.K.; Bhatt, J.C. Biomass, carbon stock under different production systems in the mid hills of Indian Himalaya. Trop Ecol. 2017, 58, 15–21. [Google Scholar]
- Lewis, S.L.; Lopez-Gonzalez1, G.; Sonke, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L. Increasing carbon storage in intact African tropical forests. Nat. Letter. 2009, 457, 1003–1006. [Google Scholar] [CrossRef]
- FAO. The State of Food Insecurity in the World; Food and Agriculture Organization of the United Nations: Rome Italy, 2001. [Google Scholar]
- Lal, M.; Singh, R. Carbon sequestration potential of Indian forests. Environ. Monit Assess. 2000, 60, 315–327. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 310, 1628–1632, Green Version. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Global Forest Resources Assessment 2010-Main Report; FAO Forestry Paper 163; FAO: Rome, Italy, 2010; 340p. [Google Scholar]
- Amazonas, N.T.; Forrester, D.I.; Silva, C.C.; Almeida, D.R.A.; Rodrigues, R.R.; Brancalion, P.H.S. High diversity mixed plantations of Eucalyptus and native trees: An interface between production and restoration for the tropics. For. Ecol. Manag. 2018, 417, 247–256. [Google Scholar] [CrossRef]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, e00419. [Google Scholar] [CrossRef]
- Deng, L.; Han, Q.S.; Zhang, C.; Tang, Z.S.; Shangguan, Z.P. Above-ground and below-ground ecosystem biomass accumulation and carbon sequestration with Caragana korshinskii Kom plantation development. Land Degrad. Dev. 2017, 28, 906–917. [Google Scholar] [CrossRef]
- Zheng, D.; Rademacher, J.; Chen, J.; Crow, T.; Bresee, M.; Le Moine, J.; Ryu, S.R. Estimating Aboveground Biomass Using Landsat 7 ETM+ Data across a Managed Landscape in Northern Wisconsin, USA. Remote Sens. Environ. 2004, 93, 402–411. [Google Scholar] [CrossRef]
- Negi, J.D.S.; Chauhan, P.S. Greenhouse gases mitigation potential by Sal (Shorea robusta Gaertn. f.) forest in Doon Valley. Indian For. 2002, 128, 771–778. [Google Scholar]
- ITTO. Encouraging Industrial Forest Plantations in the Tropics; International Tropical Timber Organization: Yokohama, Japan, 2009. [Google Scholar]
- Krishnamurthy, Y.L.; Prakasha, H.M.; Nanda, A.; Krishnappa, M.; Dattaraja, H.S.; Suresh, H.S. Vegetation structure and floristic composition of a tropical dry deciduous forest in bhadra wildlife sanctuary, Karnataka, India. Trop. Ecol. 2010, 51, 235–246. [Google Scholar]
- Arunkumar, A.N.; Warrier, R.R.; Kher, M.M.; Teixeira da Silva, J.A. Indian rosewood (Dalbergia latifolia Roxb.): Biology, utilisation, and conservation practices. Trees 2022, 36, 883–898. [Google Scholar] [CrossRef]
- Röll, A.; Ramesha, M.N.; Link, R.M.; Hertel, D.; Schuldt, B.; Patil, S.L.; Hölscher, D. Water Availability Controls the Biomass Increment of Melia dubia in South India. Forests 2021, 12, 1675. [Google Scholar] [CrossRef]
- Sharma, S.K.; Shukla, S.R.; Sujatha, M.; Shashikala, S.; Kumar, P. Assessment of certain wood quality parameters of selected genotypes of Melia dubia Cav. grown in a seedling seed orchard. J. Indian Acad. Wood Sci. 2012, 9, 165–169. [Google Scholar] [CrossRef]
- Thakur, N.S.; Mohanty, S.; Gunaga, R.P.; Gajbhiye, N.A. Melia dubia Cav. spatial geometries influence the growth, yield and essential oil principles content of Cymbopogon flexuosus (Nees Ex Steud.) W. Watson. Agrofor. Syst. 2020, 94, 985–995. [Google Scholar] [CrossRef]
- Parthiban, K.T.; Bharathi, A.K.; Seenivasan, R.; Kamala, K.; Rao, M.G. Integrating Melia dubia in Agroforestry farms as an alternate pulpwood species. Asia-Pac. Agrofor. Newsl. 2009, 34, 3–4. [Google Scholar]
- Sinha, S.K.; Chaudhari, P.A.; Thakur, N.S.; Jha, S.K.; Patel, D.P.; Dhaka, R.K. Melia dubia Cav. wood properties vary with age and influence the pulp and paper quality. Int. Wood Prod. J. 2019, 10, 139–148. [Google Scholar] [CrossRef]
- Kirankumar, G.K.; Patil, H.Y. Growth and productivity of Melia dubia under different plant density. J. Farm Sci. 2017, 30, 70–73. [Google Scholar]
- Henry, M.; Besnard, A.; Asante, W.A. Wood density, phytomass variations within and among trees, and allometric equations in a tropical rainforest of Africa. For. Ecol. Manag. 2010, 260, 1375–1388. [Google Scholar] [CrossRef]
- Cifuentes Jara, M.; Henry, M.; Réjou-Méchain, M.; Wayson, C.; Zapata-Cuartas, M.; Piotto, D.; Alice Guier, F.; Castañeda Lombis, H.; Castellanos López, E.; Cuenca Lara, R.; et al. Guidelines for documenting and reporting tree allometric equations. Ann. For. Sci. 2015, 72, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Bargali, S.S.; Singh, S.P.; Singh, R.P. Structure and function of an age series of Eucalypts plantations in Central Himalaya. I. Dry matter dynamics. Ann. Bot. 1992, 69, 405–411. [Google Scholar] [CrossRef]
- Lodhiyal, L.S.; Singh, R.P.; Singh, S.P. Structure and function of an age series of poplar plantations in Central Himalaya. I. Dry matter dynamics. Ann. Bot. 1995, 76, 191–199. [Google Scholar] [CrossRef]
- Jha, K.K. Carbon storage and sequestration rate assessment and allometric model development in young Teak plantations of tropical moist deciduous forest. India. J. For. Res. 2015, 26, 589–604. [Google Scholar] [CrossRef]
- Lodhiyal, N.; Lodhiyal, L.S.; Pangtey, Y.P.S. Structure and function of Shisham forests in Central Himalaya, India: Dry matter dynamics. Ann. Bot. 2002, 89, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piper, C.S. Soil and Plant Analysis; Hans Publisher: Bombay, India, 1966. [Google Scholar]
- Misra, R. Ecology Work Book; Oxford and IBH Publishing Co.: Calcutta, India, 1968. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 3. Chemical Methods; Black, C.A., Ed.; Soil Science of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part 2 Chemical and Microbiological Properties; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Morwin, H.D.; Peach, P.M. Exchangeability of soil potassium in and, silt and clay fractions as influenced by the nature of complementary exchangeable cations. Proc. Natl. Acad. Sci. USA 1951, 15, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis, Part 3-Chemical Methods; Sparks, D.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. The Interrelations of Certain Analytic and Synthetic Phytosociological Characters. Ecology 1950, 31, 434–455. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fromard, F.; Higuchi, N.; Kira, T.; Lescure, J.P.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Picard, N.; Saint-André, L.; Henry, M. Manual for Building Tree Volume and Biomass Allometric Equations: From Field Measurement to Prediction; Food and Agricultural Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Rawat, Y.S.; Singh, J.S. Structure and function of oak forests in Central Himalaya. I. Dry matter dynamics. Ann. Bot. 1988, 62, 397–411. [Google Scholar] [CrossRef]
- MacDicken, K.G. A Guide to Mountain Carbon Storage in Forestry and Agroforestry Projects; Winrock International Institute for Agricultural Development, Forest Carbon Monitoring Programme: Arlington, VA, USA, 1997. [Google Scholar]
- IPCC. IPCC Guidelines for National Greenhouse Gas Inventories; Prepared by the National Greenhouse Gas Inventories Programme; IGES: Hayama, Japan, 2006. [Google Scholar]
- Brown, S.; Lugo, A.E. The storage and production of organic matter in tropical forests and their role in global carbon cycle. Biotropica 1982, 14, 161–187. [Google Scholar] [CrossRef]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; FAO Forestry paper 134; Food and Agriculture Organization: Rome, Italy, 1997; 55p. [Google Scholar]
- Preece, N.D.; van Oosterzee, P.; Unda, G.C.H.; Lawes, M.J. National Carbon Model Not Sensitive to Species, Families and Site Characteristics in a Young Tropical Reforestation Project. For. Ecol. Manag. 2017, 392, 115–124. [Google Scholar] [CrossRef]
- Dawit, S.; Fritzsche, F.; Tekalign, M.; Lehmann, J.; Zech, W. Phosphorus forms and Dynamics as influenced by land use changes in the sub-humid Ethiopian highlands. Geoderma. 2002, 105, 21–48. [Google Scholar]
- Li, Y.F.; Zhang, J.J.; Chang, S.X.; Jiang, P.K.; Zhou, G.M.; Shen, Z.M.; Wu, J.S.; Lin, L.; Wang, Z.S.; Shen, M.C. Converting native shrub forests to Chinese chestnut plantations and subsequent intensive management affected soil C and N pools. For. Ecol. Mgt. 2014, 312, 161–169. [Google Scholar] [CrossRef]
- Muche, M.; Kokeb, A.; Molla, E. Assessing the physicochemical properties of soil under different land use types. J. Environ. Anal. Toxicol. 2015, 5, 309. [Google Scholar]
- Singh, J.S.; Singh, S.P. Forest vegetation of the Himalaya. Bot. Rev. 1987, 53, 80–192. [Google Scholar] [CrossRef]
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D.; Havlin, J.L. Soil Fertility and Fertilizers; Macmillan: New York, NY, USA, 1993; pp. 176–229. [Google Scholar]
- Chauhan, S.K.; Singh, S.; Sharma, S.; Vashist, B.B.; Sharma, R.; Saralch, H.S. Soil health (physical, chemical and biological) status under short rotation tree plantations on riverain soils. J. Pharmacogn. Phytochem. 2018, 7, 1599–1605. [Google Scholar]
- Swamy, S.L.; Mishra, A.; Puri, S. Comparison of growth, biomass and nutrient distribution in five promising clones of Populus deltoides under an agri-silviculture system. Bioresource Tech. 2006, 97, 57–68. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, K.N. Tree growth and nutrient status of soil in a poplar (Populus deltoids Bartr.) based agroforestry system in Punjab, India. Agrofor. Syst. 2007, 70, 113–124. [Google Scholar] [CrossRef]
- Mehra, M.S.; Singh, J.S. Pattern of wood litterfall in five forest located along an altitudinal gradient in Central Himalayan forests. Vegetatio 1985, 63, 3–11. [Google Scholar] [CrossRef]
- Kale, M.; Singh, S.; Roy, P.S.; Deosthali, V.; Ghole, V.S. Biomass equations of dominant species of dry deciduous forest in Shivpuri district, Madhya Pradesh. Curr.Sci. 2004, 87, 683–687. [Google Scholar]
- Basuki, T.M.; van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. & Manag. 2009, 257, 1684–1694. [Google Scholar]
- Basuki, T.M.; Leksono, B.; Baral, H.; Andini, S.; Wahyuni, N.S.; Artati, Y.; Choi, E.; Shin, S.; Kim, R.; Yang, A.-R.; et al. Allometric Equations for the Biomass Estimation of Calophyllum inophyllum L. in Java, Indonesia. Forests 2022, 13, 1057. [Google Scholar] [CrossRef]
- Lima, A.J.N.; Suwa, R.; de Mello Ribeiro, G.H.P.; Kajimoto, T.; dos Santos, J.; da Silva, R.P.; de Souza, C.A.S.; de Barros, P.C.; Noguchi, H.; Ishizuka, M. Allometric Models for Estimating Above- and Below-Ground Biomass in Amazonian Forests at São Gabriel da Cachoeira in the Upper Rio Negro, Brazil. For. Ecol. Manag. 2012, 277, 163–172. [Google Scholar] [CrossRef]
- Kanime, N.; Kaushal, R.; Tewari, S.K.; Raverkar, K.P.; Chaturvedi, S.; Chaturvedi, O.P. Biomass production and carbon sequestration in different tree-based systems of Central Himalayan Tarai region. For. Trees Livelihoods. 2013, 22, 38–50. [Google Scholar] [CrossRef]
- Marziliano, P.A.; Lafortezza, R.; Medicamento, U.; Lorusso, L.; Giannico, V.; Colangelo, G.; Sanesi, G. Estimating Belowground Biomass and Root/Shoot Ratio of Phillyrea Latifolia L. in the Mediterranean Forest Landscapes. Ann. For. Sci. 2015, 72, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Manhas, R.K.; Negi, J.D.S.; Kumar, R.; Chauhan, P.S. Temporal assessment of growing stock, biomass and carbon stock of Indian forests. Clim. Chang. 2006, 74, 191–221. [Google Scholar] [CrossRef]
- Hooda, N.; Gera, M.; Andrasko, K.; Sathaye, J.A.; Gupta, M.K.; Vasistha, H.B.; Chandran, M.; Rassaily, S.S. Community and farm forestry climate mitigation projects: Case studies from Uttaranchal, India. Mitig. Adapt. Strateg. Glob. Chang. 2007, 12, 1099–1130. [Google Scholar] [CrossRef]
- Baishya, R.; Barik, S.K.; Upadhaya, K. Distribution Pattern of Aboveground Biomass in Natural and Plantation Forests of Humid Tropics in Northeast India. Trop. Ecol. 2009, 50, 295–304. [Google Scholar]
- Bhadwal, S.; Singh, R. Carbon sequestration estimates for forestry options under different land-use scenarios in India. Curr. Sci. 2002, 83, 1380–1386. [Google Scholar]
- Singh, V.; Tewari, A.; Kushwaha, S.P.S.; Dadhwal, V.K. Formulating allometric equations for estimating biomass and carbon stock in small diameter trees. For. Ecol. Manag. 2011, 261, 1945–1949. [Google Scholar] [CrossRef]
- Rana, B.S.; Singh, S.P.; Singh, R.P. Biomass and net primary productivity in Central Himalayan Forest along an altitudinal gradient. For. Ecol. Manag. 1989, 27, 199–218. [Google Scholar] [CrossRef]
- Karmacharya, S.B.; Singh, K.P. Biomass and net productivity of teak plantation in dry tropical region of India. For. Ecol. Manag. 1992, 55, 233–247. [Google Scholar] [CrossRef]
- Kaul, M.; Mohren, G.M.J.; Dadhwal, V.K. Carbon storage and sequestration potential of selected tree species in India. Mitig. Adapt. Strateg. Glob. Chang. 2010, 15, 489–510. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, R.K.; Raghubanshi, A.S.; Singh, J.S. Carbon density and accumulation in woody species of tropical dry forest in India. For. Ecol. Manag. 2011, 262, 1576–1588. [Google Scholar] [CrossRef]
- Singh, A.K.; Sahu, C.; Sahu, S.K. Carbon sequestration potential of a teak plantation forest in the Eastern Ghats of India. J. Environ. Biol. 2020, 41, 770–775. [Google Scholar] [CrossRef]
- Dabi, H.; Bordoloi, R.; Das, B.; Paul, A.; Tripathi, O.P.; Mishra, B.P. Biomass, carbon stock and soil physicochemical properties in plantation of East Siang district, Arunachal Pradesh, India. Environ. Chal. 2021, 4, 100191. [Google Scholar] [CrossRef]






| Plantations | ||
|---|---|---|
| Parameter | D. latifolia | M. composita |
| Altitude (m) | 242 | 242 |
| Tree density (trees ha−1) | 880 | 960 |
| Herb density (ind m−2) | 45.91 | 32.87 |
| Shrub density (ind ha−1) | 3240 | 3740 |
| Mean change dbh (4–5 years old)(cm) | 3.1 | 3.2 |
| Mean change dbh (6–7 years old)(cm) | 2.8 | 3.8 |
| Plantations/Tree Component | a (Intercept) | b (Slope) | r2 |
|---|---|---|---|
| D. latifolia | 4 Years | ||
| Bole | −0.8249 | 1.5256 | 0.985 |
| Branch | −0.5632 | 0.8967 | 0.932 |
| Twigs | −0.0356 | 0.5845 | 0.971 |
| Foliage | −0.0548 | 0.6495 | 0.939 |
| Stump root | −0.2451 | 0.8216 | 0.978 |
| Lateral roots | −0.0628 | 0.6371 | 0.979 |
| Fine roots | −0.0025 | 0.0416 | 0.827 |
| 6 Years | |||
| Bole | −2.3226 | 1.9436 | 0.993 |
| Branch | −0.7066 | 0.9375 | 0.968 |
| Twigs | −0.0513 | 0.6501 | 0.918 |
| Foliage | −0.0807 | 0.7031 | 0.836 |
| Stump root | −0.6195 | 0.9849 | 0.990 |
| Lateral roots | −0.1251 | 0.7215 | 0.996 |
| Fine roots | −0.0084 | 0.1293 | 0.876 |
| M. composita | 4 Years | ||
| Bole | −3.3641 | 2.4906 | 0.916 |
| Branch | −0.2293 | 0.9154 | 0.986 |
| Twigs | −0.0371 | 0.5243 | 0.978 |
| Foliage | −0.0676 | 0.7527 | 0.931 |
| Stump root | −0.3819 | 0.9573 | 0.911 |
| Lateral roots | 0.0727 | 0.6010 | 0.987 |
| Fine roots | −0.0026 | 0.0302 | 0.874 |
| 6 Years | |||
| Bole | −4.8561 | 2.7284 | 0.901 |
| Branch | −1.0862 | 1.1333 | 0.989 |
| Twigs | −0.0439 | 0.5908 | 0.964 |
| Foliage | −0.0838 | 0.7722 | 0.887 |
| Stump root | −0.9704 | 1.1868 | 0.936 |
| Lateral roots | −0.3232 | 0.7958 | 0.960 |
| Fine roots | −0.0048 | 0.2281 | 0.858 |
| Species/Components | Biomass (Mg ha−1) | ||||
|---|---|---|---|---|---|
| D. latifolia | 4 Years | 5 Years | 6 Years | 7 Years | Mean Change |
| Bole | 32.36 ± 12.34 (47.0) | 36.54 ± 10.45 (49.1) | 41.2 ± 14.57 (50.7) | 47.38 ± 11.73 (52.4) | 39.37 (49.8) |
| Branches | 8.77 ± 1.92 (12.7) | 9.18 ± 1.83 (12.3) | 9.72 ± 2.72 (12.0) | 10.43 ± 2.97 (11.5) | 9.52 (12.1) |
| Twigs | 5.41 ± 1.47 (7.9) | 5.54 ± 0.97 (7.4) | 5.75 ± 0.86 (7.1) | 6.07 ± 1.08 (6.7) | 5.69 (7.3) |
| Foliage | 6.55 ± 1.79 (9.5) | 6.72 ± 1.68 (9.0) | 6.93 ± 2.08 (8.5) | 7.28 ± 1.09 (8.1) | 6.87 (8.8) |
| Stump root | 9.65 ± 0.80 (14.0) | 10.1 ± 3.20 (13.6) | 10.74 ± 2.65 (13.2) | 11.62 ± 4.25 (12.9) | 10.53 (13.4) |
| Lateral roots | 5.24 ± 1.61 (7.6) | 5.48 ± 1.73 (7.4) | 5.94 ± 0.91 (7.3) | 6.55 ± 2.86 (7.2) | 5.80 (7.4) |
| Fine roots | 0.88 ± 0.15 (1.3) | 0.9 ± 0.56 (1.2) | 0.96 ± 0.34 (1.2) | 1.1 ± 0.57 (1.2) | 0.96 (1.2) |
| Total | 68.86 ± 20.08 | 74.46 ± 20.42 | 81.24 ± 24.13 | 90.43 ± 24.55 | 78.75 |
| M. composita | 4 Years | 5 Years | 6 Years | 7 Years | Mean Change |
| Bole | 58.66 ± 18.45 (56.3) | 68.43 ± 13.63 (59.1) | 78.8 ± 10.72 (61.0) | 91.24 ±12.77 (62.9) | 74.28 (59.8) |
| Branches | 13.13 ± 2.12 (12.6) | 13.86 ± 4.19 (12.0) | 14.86 ± 2.28 (11.5) | 16.01 ± 3.31 (11.0) | 14.46 (11.8) |
| Twigs | 4.74 ± 0.59 (4.6) | 4.81 ± 0.66 (4.2) | 5.12 ± 0.96 (4.0) | 5.42 ± 1.79 (3.7) | 5.02 (4.1) |
| Foliage | 6.34 ± 2.78 (6.1) | 6.51 ± 2.07 (5.6) | 6.97 ± 1.87 (5.4) | 7.53 ± 1.10 (5.2) | 6.84 (5.6) |
| Stump root | 13.02 ± 2.15 (12.5) | 13.56 ± 3.20 (11.7) | 14.42 ± 2.21 (11.2) | 15.57 ± 4.35 (10.7) | 14.14 (11.5) |
| Lateral roots | 7.04 ± 2.06 (6.8) | 7.23 ± 1.78 (6.2) | 7.55 ± 1.09 (5.9) | 7.9 ± 1.11 (5.4) | 7.43 (6.1) |
| Fine roots | 1.356 ± 0.24 (1.3) | 1.381 ± 0.46 (1.2) | 1.422 ± 0.37 (1.1) | 1.473 ± 0.65 (1.01) | 1.41 (1.2) |
| Total | 104.28 ± 28.69 | 115.78 ± 25.99 | 129.14 ± 19.50 | 145.14 ± 25.08 | 123.59 |
| Age (Years) | ||||
|---|---|---|---|---|
| 4 | 5 | 6 | 7 | |
| Tree | 68.86 ± 8.08 | 74.46 ± 8.42 | 81.24 ± 9.63 | 90.43 ± 10.95 |
| Shrub | 1.41 ± 0.13 | 1.29 ± 0.22 | 1.32 ± 0.46 | 2.08 ± 0.67 |
| Herb | 2.36 ± 0.41 | 3.05 ± 0.83 | 2.67 ± 0.86 | 3.68 ± 0.84 |
| Litter | 3.33 ± 0.63 | 3.11± 1.01 | 3.58 ± 1.53 | 4.25 ± 1.65 |
| Age (Years) | ||||
|---|---|---|---|---|
| 4 | 5 | 6 | 7 | |
| Tree | 104.28 ± 15.39 | 115.78 ± 14.59 | 129.14 ± 18.20 | 145.14 ± 19.57 |
| Shrub | 1.44 ± 0.21 | 2.02 ± 0.32 | 1.85 ± 0.37 | 2.41 ± 0.23 |
| Herb | 2.08 ± 0.94 | 1.14 ± 0.13 | 2.46 ± 0.78 | 1.83 ± 0.17 |
| Litter | 2.77 ± 0.67 | 3.38 ± 1.28 | 4.13 ± 1.38 | 3.72 ± 0.98 |
| Age (Years) | ||||
|---|---|---|---|---|
| D. latifolia | 4 | 5 | 6 | 7 |
| Leaf | 1.19 ± 0.17 (35.7) | 1.22 ± 0.19 (39.1) | 1.36 ± 0.18 (38) | 1.58 ± 0.20 (36.9) |
| Miscellaneous | 1.28 ± 0.16 (38.4) | 1.12 ± 0.17 (35.9) | 1.38 ± 0.17 (38.5) | 1.61 ± 0.17 (37.6) |
| Wood | 0.86 ± 0.13 (25.8) | 0.78 ± 0.13 (25.0) | 0.84 ± 0.13 (23.5) | 1.09 ± 0.14 (25.5) |
| Age (Years) | ||||
|---|---|---|---|---|
| M. composita | 4 | 5 | 6 | 7 |
| Leaf | 1.09 ± 0.185 (39.4) | 1.2 ± 0.194 (35.7) | 1.54 ± 0.219 (39.6) | 1.4 ± 0.196 (35.1) |
| Miscellaneous | 0.9 ± 0.164 (32.5) | 1.19 ± 0.204 (35.4) | 1.45 ± 0.207 (37.3) | 1.3 ± 0.174 (32.6) |
| Wood | 0.78 ± 0.127 (28.2) | 0.97 ± 0.148 (28.9) | 0.9 ± 0.126 (23.1) | 1.29 ± 0.141 (32.3) |
| Species/Components | Carbon Stock (Mg C ha−1) | |||
|---|---|---|---|---|
| D. latifolia | 4 Years | 5 Years | 6 Years | 7 Years |
| Bole | 15.37 ±1.21 (46.99) | 17.36 ±1.34 (49.07) | 19.57 ±1.28 (50.71) | 22.51 ±1.30 (52.39) |
| Branches | 4.17 ±0.1 (12.74) | 4.36 ±0.13 (12.33) | 4.62 ±0.17 (11.96) | 4.95 ±0.28 (11.53) |
| Twigs | 2.57 ±0.15 (7.86) | 2.63 ±0.1 (7.44) | 2.73 ±0.15 (7.08) | 2.88 ±0.11 (6.71) |
| Foliage | 3.11 ±0.18 (9.51) | 3.19 ±0.16 (9.02) | 3.29 ±0.32 (8.53) | 3.46 ±0.33 (8.05) |
| Stump root | 4.58 ±0.18 (14.01) | 4.8 ±0.18 (13.56) | 5.1 ±0.12 (13.22) | 5.52 ±0.40 (12.85) |
| Lateral roots | 2.49 ±0.29 (7.61) | 2.6 ±0.23 (7.36) | 2.82 ±0.28 (7.31) | 3.11 ±0.30 (7.24) |
| Fine roots | 0.42 ± 0.04 (1.28) | 0.43 ±0.03 (1.21) | 0.46 ±0.04 (1.18) | 0.52 ±0.06 (1.22) |
| Total | 32.71 ± 2.15 | 35.37 ± 2.28 | 38.59 ± 2.35 | 42.95 ± 2.77 |
| M. composita | 4 Years | 5 Years | 6 Years | 7 Years |
| Bole | 27.86 ±1.21 (56.25) | 32.5 ±1.32 (59.1) | 37.43 ±1.35 (61.02) | 43.34 ±1.44 (62.86) |
| Branches | 6.24 ±0.27 (12.59) | 6.58 ±0.30 (11.97) | 7.06 ±0.37 (11.51) | 7.6 ±0.40 (11.03) |
| Twigs | 2.25 ±0.12 (4.55) | 2.28 ±0.13 (4.15) | 2.43 ±0.21 (3.96) | 2.57 ±0.23 (3.73) |
| Foliage | 3.01 ±0.10 (6.08) | 3.09 ±0.14 (5.62) | 3.31 ±0.16 (5.4) | 3.58 ±0.18 (5.19) |
| Stump root | 6.18 ±0.14 (12.48) | 6.44 ±0.18 (11.71) | 6.85 ±0.14 (11.17) | 7.4 ±0.15 (10.73) |
| Lateral roots | 3.34 ±0.18 (6.75) | 3.43 ±0.19 (6.24) | 3.59 ±0.18 (5.85) | 3.75 ±0.24 (5.44) |
| Fine roots | 0.64 ±0.05 (1.3) | 0.66 ±0.04 (1.19) | 0.68 ±0.02 (1.1) | 0.7 ±0.03 (1.01) |
| Total | 49.54 ± 2.07 | 55 ± 2.29 | 61.34 ± 2.43 | 68.94 ± 2.67 |
| Tree Components | Carbon Sequestration (Mg C ha−1 yr−1) | |||||
|---|---|---|---|---|---|---|
| D. latifolia | M. composita | |||||
| 4 Years | 5 Years | 6 Years | 4 Years | 5 Years | 6 Years | |
| Bole | 1.99 ± 0.21 | 2.21 ± 0.28 | 2.93 ± 0.37 | 4.64 ± 0.45 | 4.93 ± 0.63 | 5.91 ± 0.72 |
| Branches | 0.2 ± 0.15 | 0.25 ± 0.17 | 0.33 ± 0.19 | 0.35 ± 0.22 | 0.48 ± 0.29 | 0.55 ± 0.28 |
| Twigs | 0.06 ± 0.03 | 0.1 ± 0.04 | 0.15 ± 0.05 | 0.03 ± 0.09 | 0.15 ± 0.06 | 0.14 ± 0.06 |
| Foliage | 0.08 ± 0.02 | 0.1 ± 0.05 | 0.16 ± 0.04 | 0.08 ± 0.08 | 0.22 ± 0.07 | 0.27 ± 0.07 |
| Stump root | 0.21 ± 0.09 | 0.31 ± 0.1 | 0.418 ± 0.12 | 0.26 ± 0.15 | 0.41 ± 0.20 | 0.55 ± 0.21 |
| Lateral roots | 0.11 ± 0.03 | 0.22 ± 0.07 | 0.28 ± 0.17 | 0.09 ± 0.06 | 0.15 ± 0.08 | 0.17 ± 0.09 |
| Fine roots | 0.01 ± 0.01 | 0.03 ± 0.02 | 0.06 ± 0.02 | 0.01 ± 0.04 | 0.02 ± 0.06 | 0.02 ± 0.07 |
| Total | 2.66 ± 0.54 | 3.22 ± 0.73 | 4.33 ± 0.86 | 5.46 ± 0.52 | 6.35 ± 0.84 | 7.6 ± 0.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chopra, N.; Tewari, L.M.; Tewari, A.; Wani, Z.A.; Asgher, M.; Pant, S.; Siddiqui, S.; Siddiqua, A. Estimation of Biomass and Carbon Sequestration Potential of Dalbergia latifolia Roxb. and Melia composita Willd. Plantations in the Tarai Region (India). Forests 2023, 14, 646. https://doi.org/10.3390/f14030646
Chopra N, Tewari LM, Tewari A, Wani ZA, Asgher M, Pant S, Siddiqui S, Siddiqua A. Estimation of Biomass and Carbon Sequestration Potential of Dalbergia latifolia Roxb. and Melia composita Willd. Plantations in the Tarai Region (India). Forests. 2023; 14(3):646. https://doi.org/10.3390/f14030646
Chicago/Turabian StyleChopra, Neha, Lalit Mohan Tewari, Ashish Tewari, Zishan Ahmad Wani, Mohd Asgher, Shreekar Pant, Sazada Siddiqui, and Ayesha Siddiqua. 2023. "Estimation of Biomass and Carbon Sequestration Potential of Dalbergia latifolia Roxb. and Melia composita Willd. Plantations in the Tarai Region (India)" Forests 14, no. 3: 646. https://doi.org/10.3390/f14030646
APA StyleChopra, N., Tewari, L. M., Tewari, A., Wani, Z. A., Asgher, M., Pant, S., Siddiqui, S., & Siddiqua, A. (2023). Estimation of Biomass and Carbon Sequestration Potential of Dalbergia latifolia Roxb. and Melia composita Willd. Plantations in the Tarai Region (India). Forests, 14(3), 646. https://doi.org/10.3390/f14030646