Distribution of Plant Hormones and Their Precursors in Cambial Region Tissues of Quercus myrsinifolia and Castanopsis cuspidata var.sieboldii after Bending Stems or Applying Ethylene precursor

Abstract
:1. Introduction
2. Materials and Methods
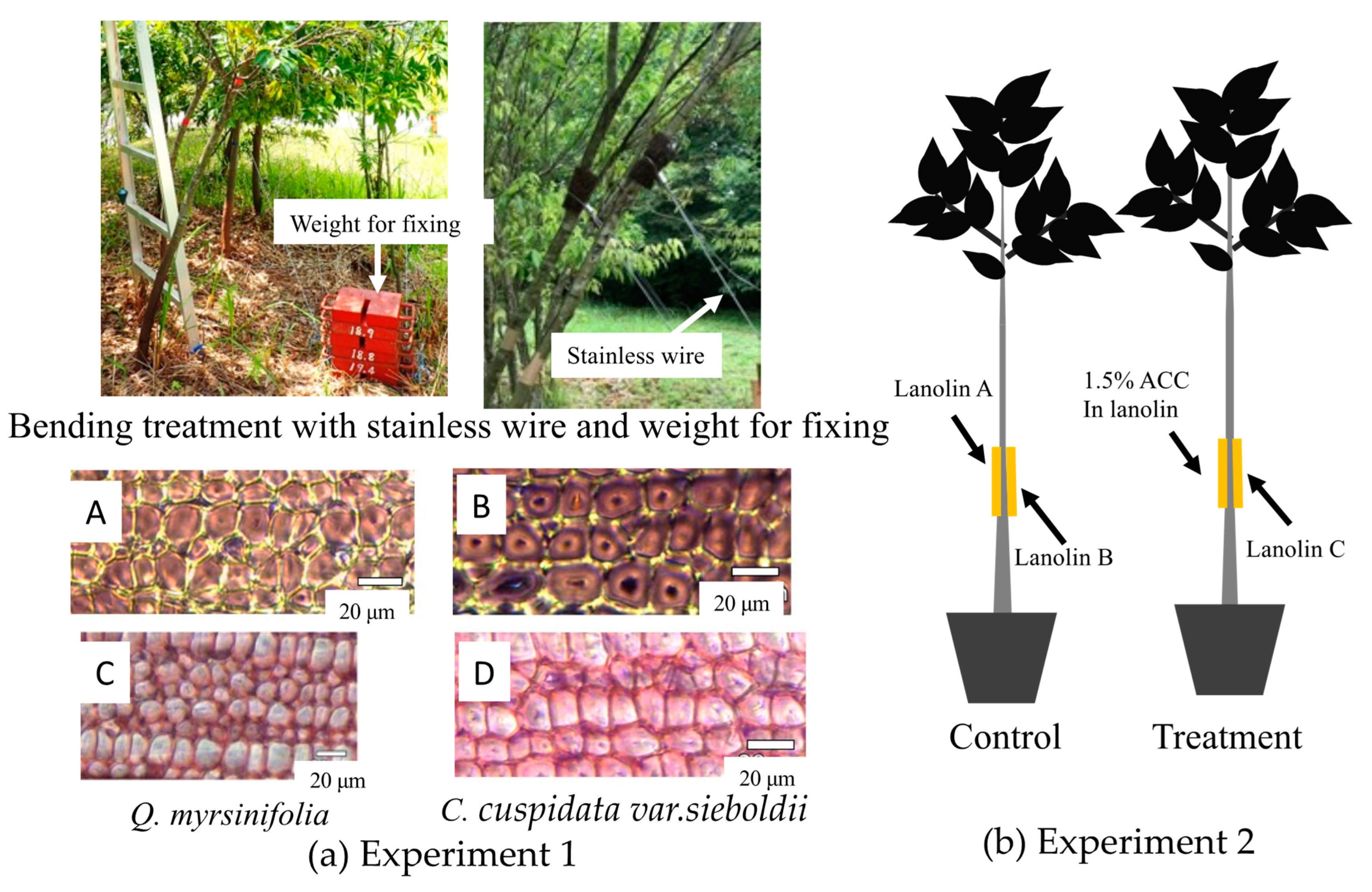
2.1. Sample Trees and Stem Bending Procedure
2.2. Sample Seedlings and Treatment Procedure for ACC Application
2.3. Observation of Wood Anatomy
2.4. Quantification of Plant Hormones and Its Precursor
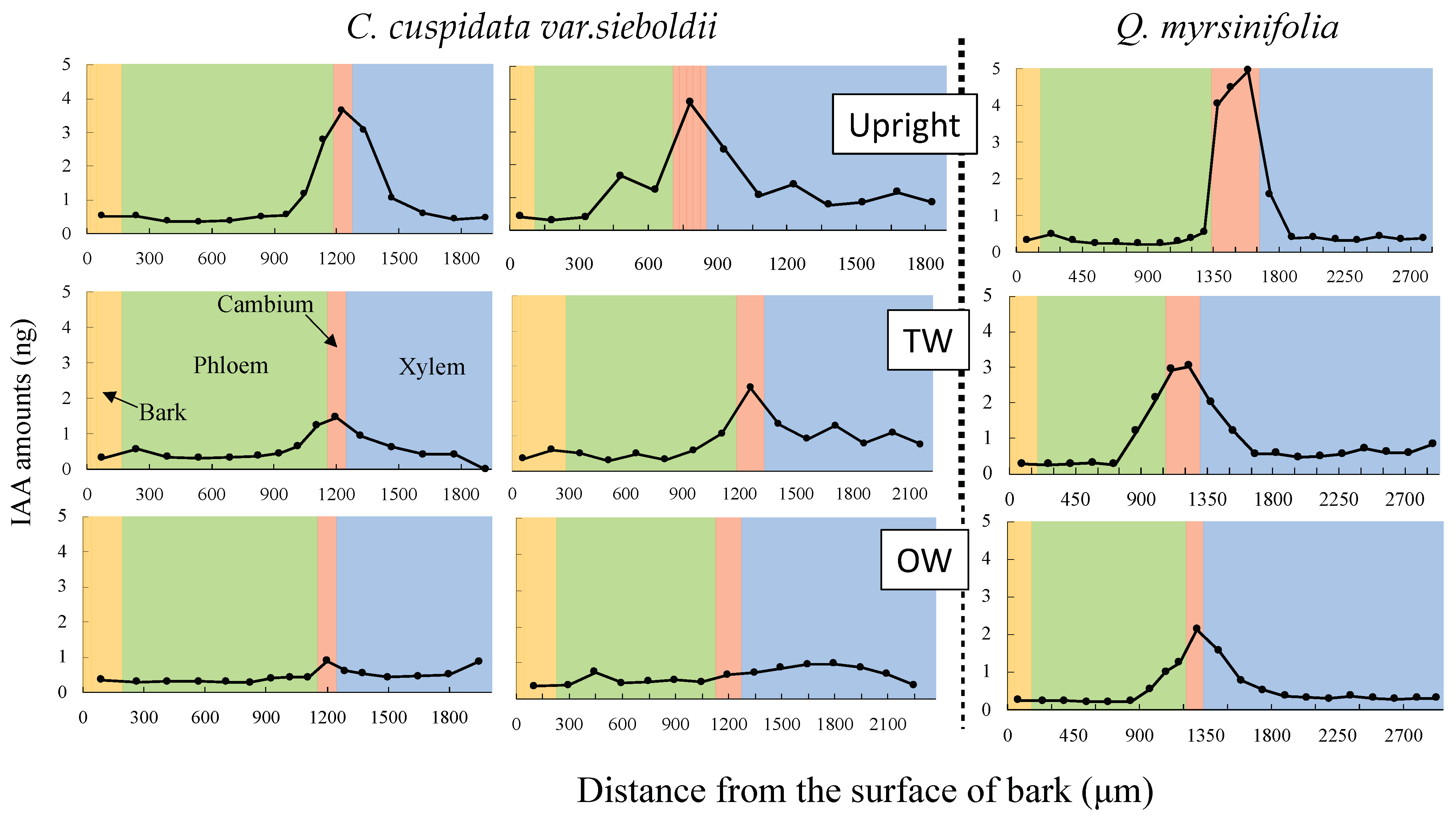
2.5. Analysis of Radial Distribution of IAA with Cryo-Sectioning
2.6. Statistical Analysis
3. Results
3.1. Wood Anatomy Formed after Bending Treatment
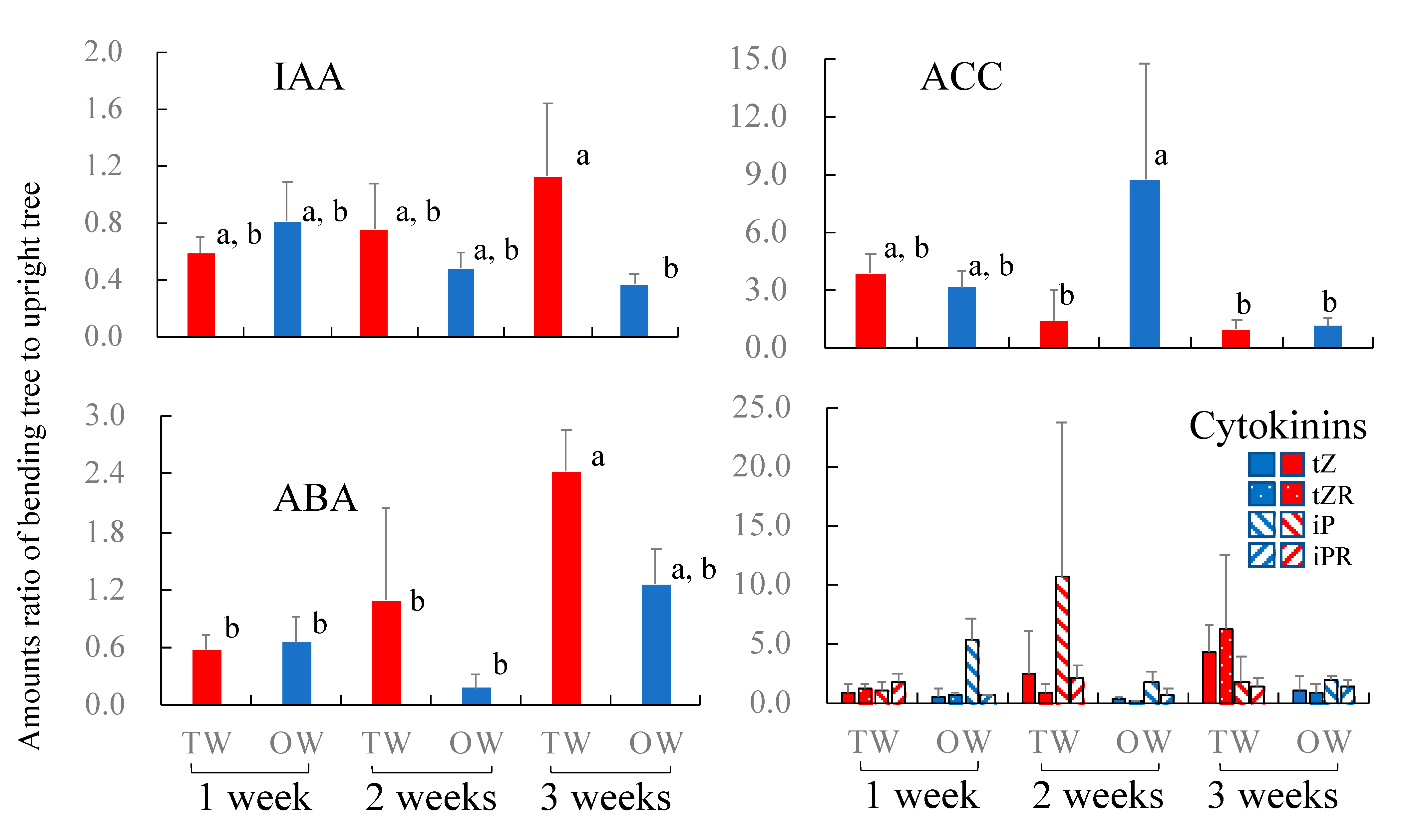
3.2. Distribution of Plant Hormones and Their Precursor after Bending Treatment
3.3. Peak Concentrations of IAA in Cambium after 2-Week Bending
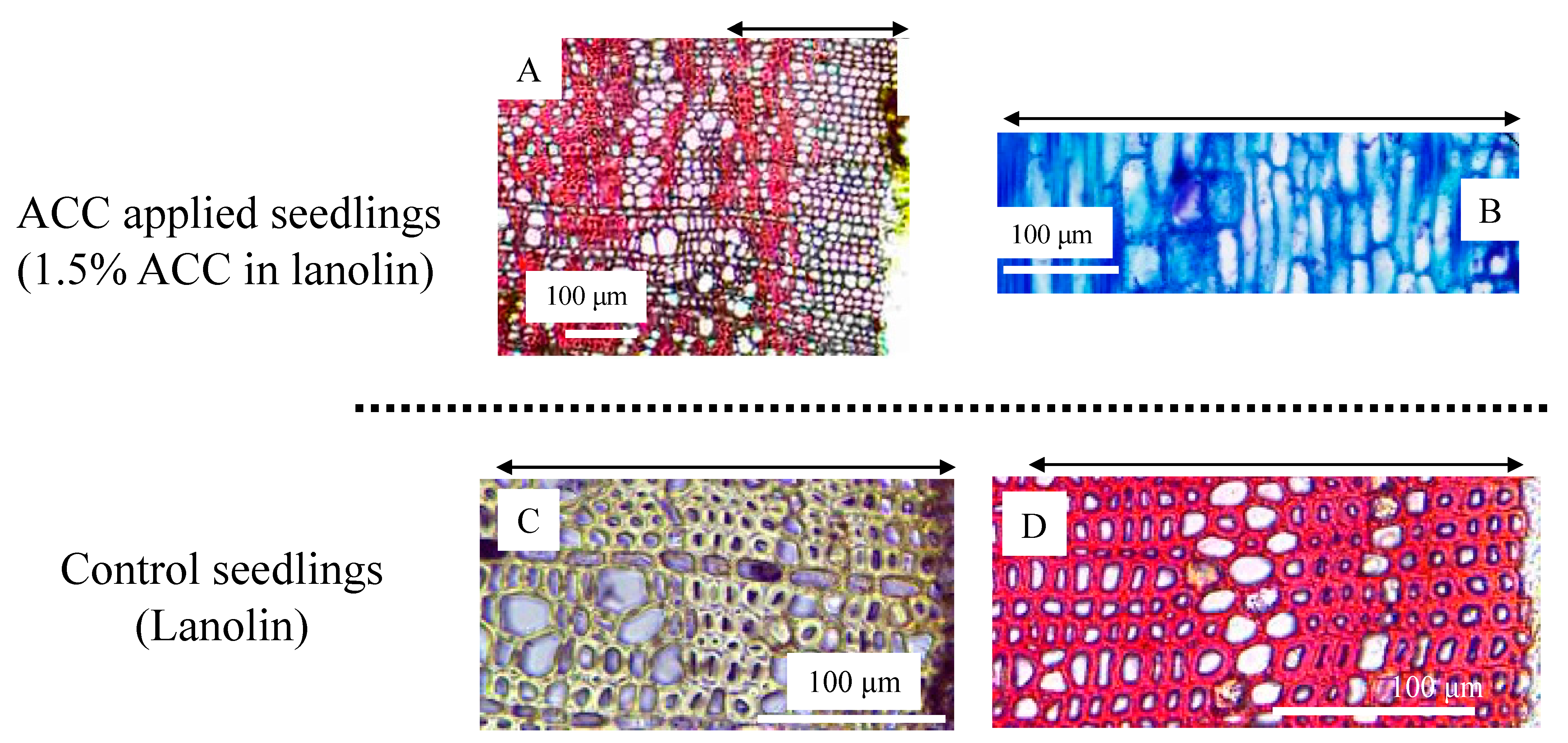
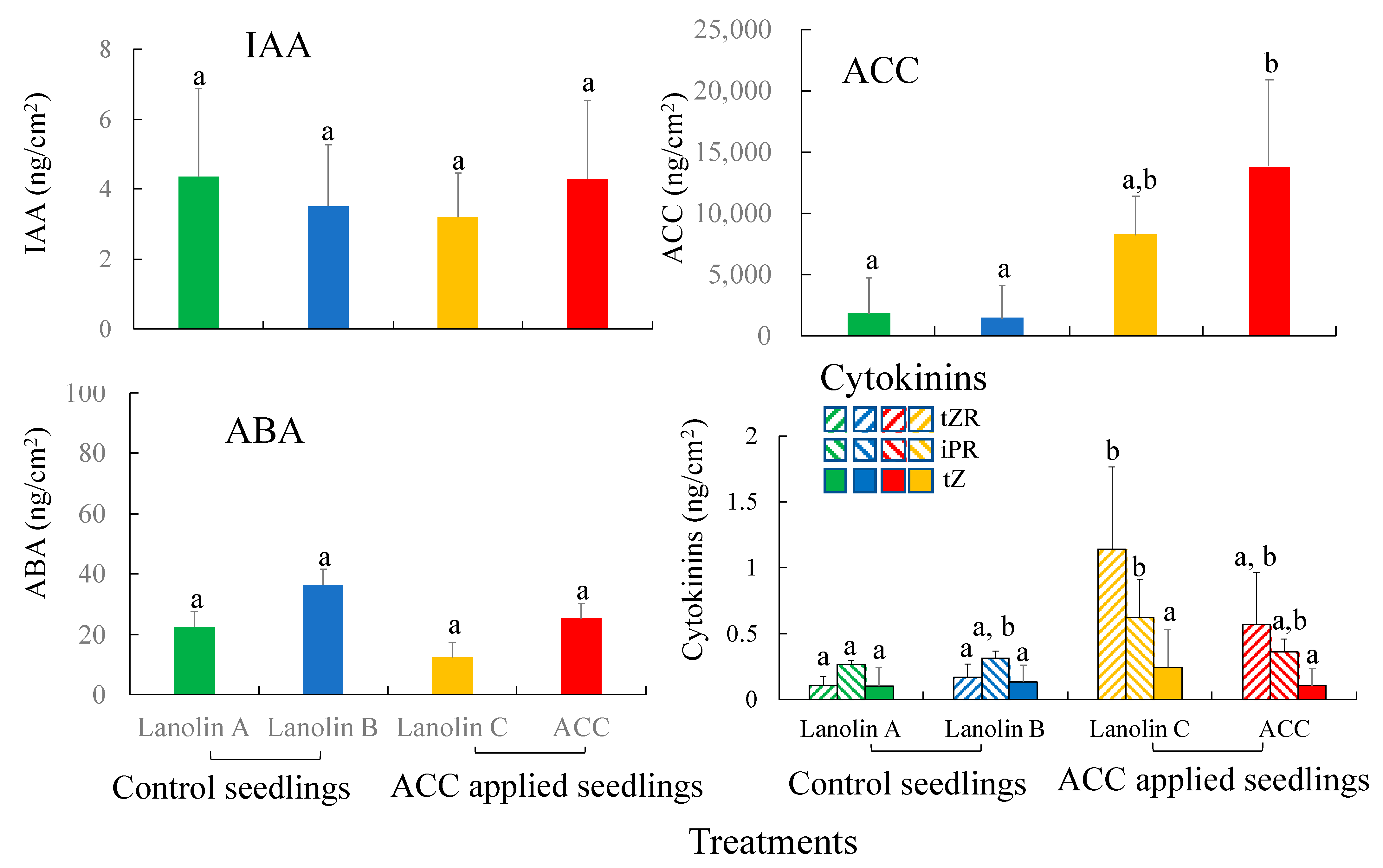
3.4. Wood Anatomy and the Distribution of Plant Hormones and Their Precursors after ACC Application in C. cuspidata var.sieboldii Seedlings
4. Discussion
4.1. The Role of IAA in TW Formation
4.2. The Role of ABA in TW Formation
4.3. The Roles of ACC and Cytokinins in TW Formation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, R.; Ilic, J. Rapid prediction of wood stiffness from microfibril angle and density. For. Prod. J. 2001, 51, 53–57. [Google Scholar]
- Yang, J.L.; Evans, R. Prediction of MOE of eucalypt wood from microfibril angle and density. Holz. Roh. Werkstoff. 2003, 61, 449–452. [Google Scholar] [CrossRef]
- Lachenbruch, B.; Johnson, G.R.; Downes, G.M.; Evans, R. Relationships of density, microfibril angle, and sound velocity with stiffness and strength in mature wood of Douglas-fir. Can. J. For. Res. 2010, 40, 55–64. [Google Scholar] [CrossRef]
- Mclean, J.P.; Evans, R.; Moore, J.R. Predicting the longitudinal modulus of elasticity of Sitka spruce from cellulose orientation and abundance. Holzforschung 2010, 64, 495–500. [Google Scholar] [CrossRef]
- Yamashita, K.; Hirakawa, Y.; Nakatani, H.; Ikeda, M. Tangential and radial shrinkage variation within trees in sugi (Cryptomeria japonica) cultivars. J. Wood Sci. 2009, 55, 161–168. [Google Scholar] [CrossRef]
- Cronshaw, J.; Morey, P.R. The effect of plant growth substances on the development of tension wood in horizontally inclined stems of Acer rubrum seedlings. Protoplasma 1968, 65, 379–391. [Google Scholar] [CrossRef]
- Casperson, G. On endogenous Factors of the Reaction Wood Formation. I. Comm. Auxin application on horse-chestnut epicotyls. Planta 1965, 64, 225–240. [Google Scholar] [CrossRef]
- Cronshaw, J.; Morey, P.R. Induction of Tension Wood by 2,3,5-Tri-iodobenzoic Acid. Nature 1965, 205, 816–818. [Google Scholar] [CrossRef]
- Kennedy, R.W.; Farrar, J.L. Induction of Tension Wood with the Antiauxin 2,3,5-Triiodobenzoic acid. Nature 1965, 208, 406–407. [Google Scholar] [CrossRef]
- Hellgren, J.M.; Olofsson, K.; Sundberg, B. Patterns of auxin distribution during gravitational induction of reaction wood in poplar and pine. Plant Physiol. 2004, 135, 212–220. [Google Scholar] [CrossRef]
- Moyle, R.; Schrader, J.; Stenberg, A.; Olsson, O.; Saxena, S.; Sandberg, G.; Bhalerao, R.P. Environmental and auxin regulation of wood formation involves members of the Aux/IAA gene family in hybrid aspen. Plant J. 2002, 31, 675–685. [Google Scholar] [CrossRef]
- Nugroho, W.D.; Yamagishi, Y.; Nakaba, S.; Fukuhara, S.; Begum, S.; Marsoem, S.N.; Ko, J.; Jin, H.; Funada, R. Gibberellin is required for the formation of tension wood and stem gravitropism in Acacia mangium seedlings. Ann. Bot. 2012, 110, 887–895. [Google Scholar] [CrossRef]
- Wang, H.; Jin, Y.; Wang, C.; Li, B.; Jiang, C.; Sun, Z.; Zhang, Z.; Kong, F.; Zhang, H. Fasciclin-like arabinogalactan proteins, PtFLAs, play important roles in GA-mediated tension wood formation in Populus. Sci. Rep. 2017, 7, 6182. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Honma, T.; Nakamura, T.; Furukawa, I.; Yamamoto, F. Regulation by uniconazole-P and gibberellins of morphological and anatomical responses of Fraxinus mandshurica seedlings to gravity. IAWA J. 1998, 19, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, M.; Zhu, S.; Zhou, L.; Liu, S. Effects of exogenous 24-epibrassinolide and brassinazole on negative gravitropism and tension wood formation in hybrid poplar (Populus deltoids × Populus nigra). Planta 2019, 249, 1449–1463. [Google Scholar] [CrossRef]
- Jin, Y.; Yu, C.; Jiang, C.; Guo, X.; Li, B.; Wang, C.; Kong, F.; Zhang, H.; Wang, H. PtiCYP85A3, a BR C-6 oxidase gene, plays a critical role in brassinosteroid-mediated tension wood formation in poplar. Front. Plant Sci. 2020, 11, 468. [Google Scholar] [CrossRef]
- Du, S.; Yamamoto, F. Ethylene evolution changes in the stems of Metasequoia glyptostroboides and Aesculus turbinate seedlings in relation to gravity-induced reaction wood formation. Trees 2003, 17, 522–528. [Google Scholar] [CrossRef]
- Yamamoto, F.; Kozlowski, T.T. Effects of flooding, tilting of stems, and ethrel application on growth, stem anatomy, and ethylene production of Acer platanoides seedlings. Scand. J. For. 1987, 2, 141–156. [Google Scholar]
- Andersson-Gunneras, S.; Hellgren, J.M.; Bjorklund, S.; Regan, S.; Moritz, T.; Sundberg, B. Asymmetric expression of a poplar ACC oxidase controls ethylene production during gravitational induction of tension wood. Plant J. 2003, 34, 339–349. [Google Scholar] [CrossRef]
- Felten, J.; Vahala, J.; Love, J.; Gorzsas, A.; Ruggeberg, M.; Delhomme, N.; Lesniewska, J.; Kangasjarvi, J.; Hvidsten, T.R.; Mellerowicz, E.J.; et al. Ethylene signaling induces gelatinous layers with typical features of tension wood in hybrid aspen. New Phytol. 2018, 218, 999–1014. [Google Scholar] [CrossRef]
- Seyfferth, C.; Wessels, B.A.; Gorzsas, A.; Love, J.W.; Ruggeberg, M.; Delhomme, N.; Vain, T.; Antos, K.; Tuominen, H.; Sundberg, B.; et al. Ethylene signaling is required for fully functional tension wood in hybrid aspen. Front. Plant Sci. 2019, 10, 1101. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Hayashi, S.; Itoh, T.; Shimaji, K. Studies on the improvement of the pinning method for marking xylem growth, I. minute examination of minute examination of Pin Marks in Taeda Pine and other species. Wood Res. 1981, 67, 1–16. [Google Scholar]
- Aiso, H.; Ishiguri, F.; Toyoizumi, T.; Ohshima, J.; Iizuka, K.; Priadi, D.; Yokota, S. Anatomical, chemical, and physical characteristics of tension wood in two tropical fast-growing species, Falcataria moluccana and Acacia auriculiformis. Tropics 2016, 25, 33–41. [Google Scholar] [CrossRef]
- Kijidani, Y.; Nagai, T.; Suwashita, T.; Tsuyama, T. Seasonal variations of tracheid formation and amount of auxin (IAA) and gibberellin A4 (GA4) in cambial-region tissues of mature sugi (Cryptomeria japonica) cultivar grown in a Nelder plot with different tree densities. J. Wood Sci. 2017, 63, 315–321. [Google Scholar] [CrossRef]
- Uggla, C.; Mellerowicz, E.J.; Sundberg, B. Indole-3-acetic acid controls cambial growth in Scots pine by positional signalling. Plant Physiol. 1998, 117, 113–121. [Google Scholar] [CrossRef]
- Kijidani, Y.; Tsuyama, T.; Takata, K. Seasonal variations of auxin and gibberellin A4 levels in cambial-region tissues of three conifers (Pinus elliottii, Chamaecyparis obtusa, and Cryptomeria japonica) with inherently different wood densities. J. Wood Sci. 2021, 67, 46. [Google Scholar] [CrossRef]
- Takata, N.; Tsuyama, T.; Nagano, S.; Baba, K.; Yasuda, Y.; Sakamoto, S.; Mitsuda, N.; Taniguch, T. Prior secondary cell wall formation is required for gelatinous layer deposition and posture control in gravi-stimulated aspen. Plant J. 2021, 108, 725–736. [Google Scholar] [CrossRef]
- Du, S.; Yamamoto, F. An Overview of the Biology of Reaction Wood Formation. J. Integr. Plant Biol. 2007, 49, 131–143. [Google Scholar] [CrossRef]
- Peuke, A.D.; Schraml, C.; Hartung, W.; Rennenberg, H. Identification of drought-sensitive beech ecotypes by physiological parameters. New Phytol. 2002, 154, 373–387. [Google Scholar] [CrossRef]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolezi, P.; Azeeze, A.; Singh, R.K.; Immanen, J.; Mahler, N.; Hvidsten, T.R.; Eklund, D.M.; et al. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef]
- Covington, M.F.; Maloof, J.N.; Straume, M.; Kay, S.A.; Harmer, S.L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol. 2008, 9, R130. [Google Scholar] [CrossRef] [PubMed]
- Mwange, K.N.; Hou, H.W.; Wang, Y.Q.; He, X.Q.; Cui, K.M. Opposite patterns in the annual distribution and time-course of endogenous abscisic acid and indole-3-acetic acid in relation to the periodicity of cambial activity in Eucommia ulmoides Oliv. J. Exp. Bot. 2005, 56, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, B.; Zhang, D. Transcript profiling of Populus tomentosa genes in normal, tension, and opposite wood by RNA-seq. BMC Genom. 2015, 16, 164. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Hormones | No. | Effects on TW Formation |
|---|---|---|
| IAA | [6,7,8,9] | Negative effects |
| [10,11] | No or small effects | |
| GA3 | [12,13] | Positive effects |
| [14] | No or small effects | |
| BRs | [15] | Negative effects |
| Ethylene | [16,20] | Positive effects |
| [17,18] | No or small effects | |
| ACC | [19,20] | Positive effects |
| [21] | No or small effects |
| Species | Age | n | H (m) | D (cm) | MBA (degree) | Treatment-Period | Experiment |
|---|---|---|---|---|---|---|---|
| Q. myrsinifolia (Radial-porous wood) | 6 | 3 | 4.26 | 4.10 | 34.2 | Bending_1 W | Analysis of plant hormones |
| 2 | 4.00 | 3.65 | Upright_1 W | ||||
| 5 | 2 | 2.65 | 5.25 | 37.8 | Bending_2 W | ||
| 2 | 2.75 | 4.25 | Upright_2 W | ||||
| 6 | 3 | 5.13 | 4.83 | 30.7 | Bending_3 W | ||
| 2 | 3.82 | 3.70 | Upright_3 W | ||||
| 5 | 1 | 3.23 | 3.0 | 42.6 | Bending | Anatomy | |
| 1 | 2.7 | 3.0 | Upright | ||||
| C. cuspidata var.sieboldii (Ring-porous wood) | 5 | 2 | 3.10 | 4.00 | 43.6 | Bending_2 W | Analysis of plant hormones |
| 2 | 3.75 | 6.25 | Upright_2 W | ||||
| 5 | 1 | 3.14 | 6.0 | 40.4 | Bending | Anatomy | |
| 1 | 3.60 | 6.0 | Upright |
| Species | Age | n | H (m) | D (cm) | Treatment-Period | Experiment |
|---|---|---|---|---|---|---|
| C. cuspidata var.sieboldii (Ring-porous wood) | 2 | 5 | 0.65 | 0.76 | ACC/lanoline_1 W | Analysis of plant hormones |
| 2 | 5 | 0.65 | 0.78 | Lanolin_1 W | ||
| 2 | 2 | 0.66 | 0.90 | ACC/lanoline | Anatomy | |
| 2 | 2 | 0.60 | 0.73 | Lanolin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kijidani, Y.; Tsuyama, T.; Tokumoto, Y. Distribution of Plant Hormones and Their Precursors in Cambial Region Tissues of Quercus myrsinifolia and Castanopsis cuspidata var.sieboldii after Bending Stems or Applying Ethylene precursor. Forests 2023, 14, 813. https://doi.org/10.3390/f14040813
Kijidani Y, Tsuyama T, Tokumoto Y. Distribution of Plant Hormones and Their Precursors in Cambial Region Tissues of Quercus myrsinifolia and Castanopsis cuspidata var.sieboldii after Bending Stems or Applying Ethylene precursor. Forests. 2023; 14(4):813. https://doi.org/10.3390/f14040813
Chicago/Turabian StyleKijidani, Yoshio, Taku Tsuyama, and Yuji Tokumoto. 2023. "Distribution of Plant Hormones and Their Precursors in Cambial Region Tissues of Quercus myrsinifolia and Castanopsis cuspidata var.sieboldii after Bending Stems or Applying Ethylene precursor" Forests 14, no. 4: 813. https://doi.org/10.3390/f14040813
APA StyleKijidani, Y., Tsuyama, T., & Tokumoto, Y. (2023). Distribution of Plant Hormones and Their Precursors in Cambial Region Tissues of Quercus myrsinifolia and Castanopsis cuspidata var.sieboldii after Bending Stems or Applying Ethylene precursor. Forests, 14(4), 813. https://doi.org/10.3390/f14040813