
Mixed-Species Acacia Plantation Decreases Soil Organic Carbon and Total Nitrogen Concentrations but Favors Species Regeneration and Tree Growth over Monoculture: A Thirty-Three-Year Field Experiment in Southern China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
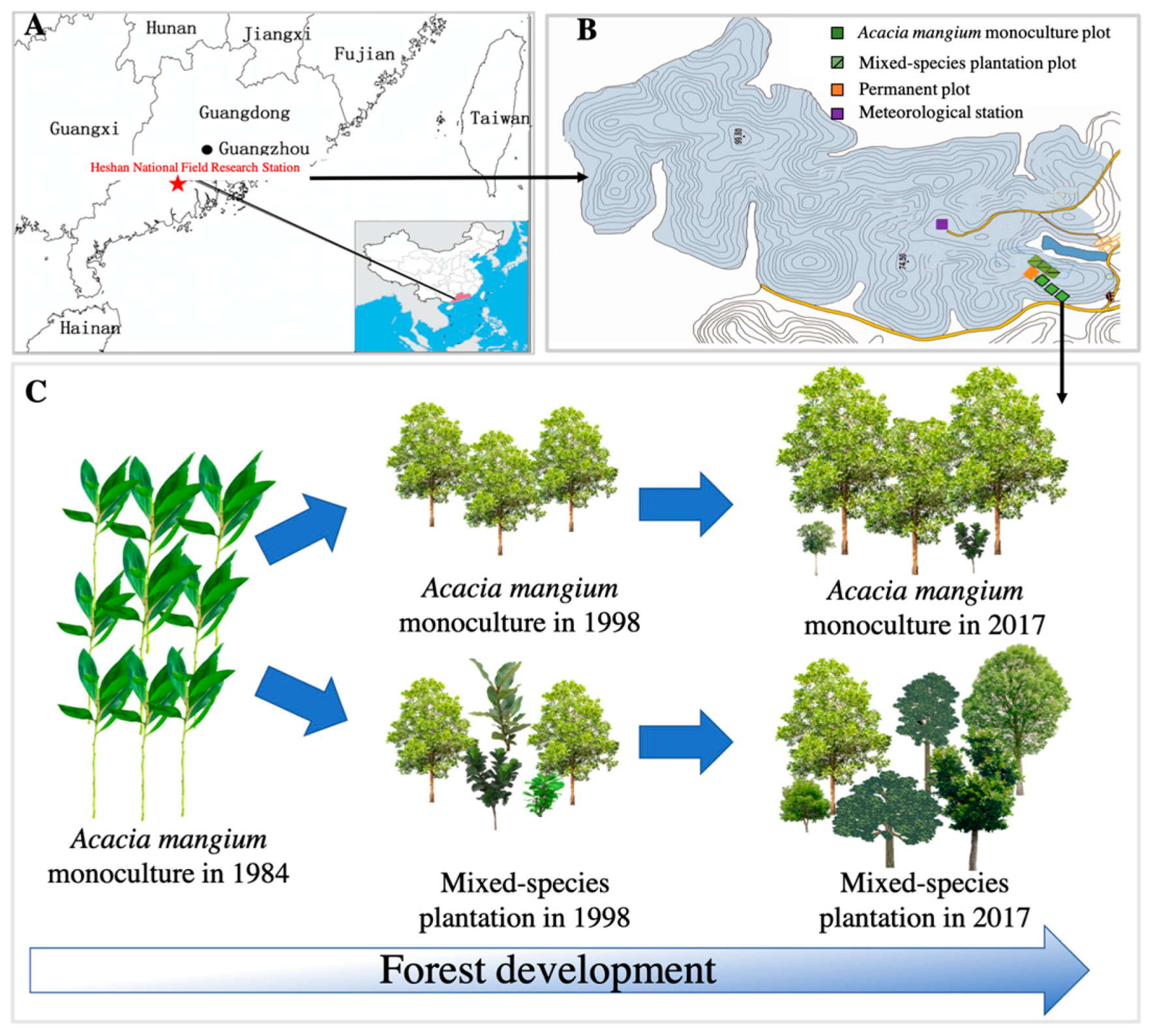
2.1. Study Site
2.2. Experimental Design
2.2.1. Plot Design
2.2.2. Soil and Leaf Sample Collections
2.2.3. Soil and Leaf Sample Property Analyses
2.3. Statistical Analyses
3. Results
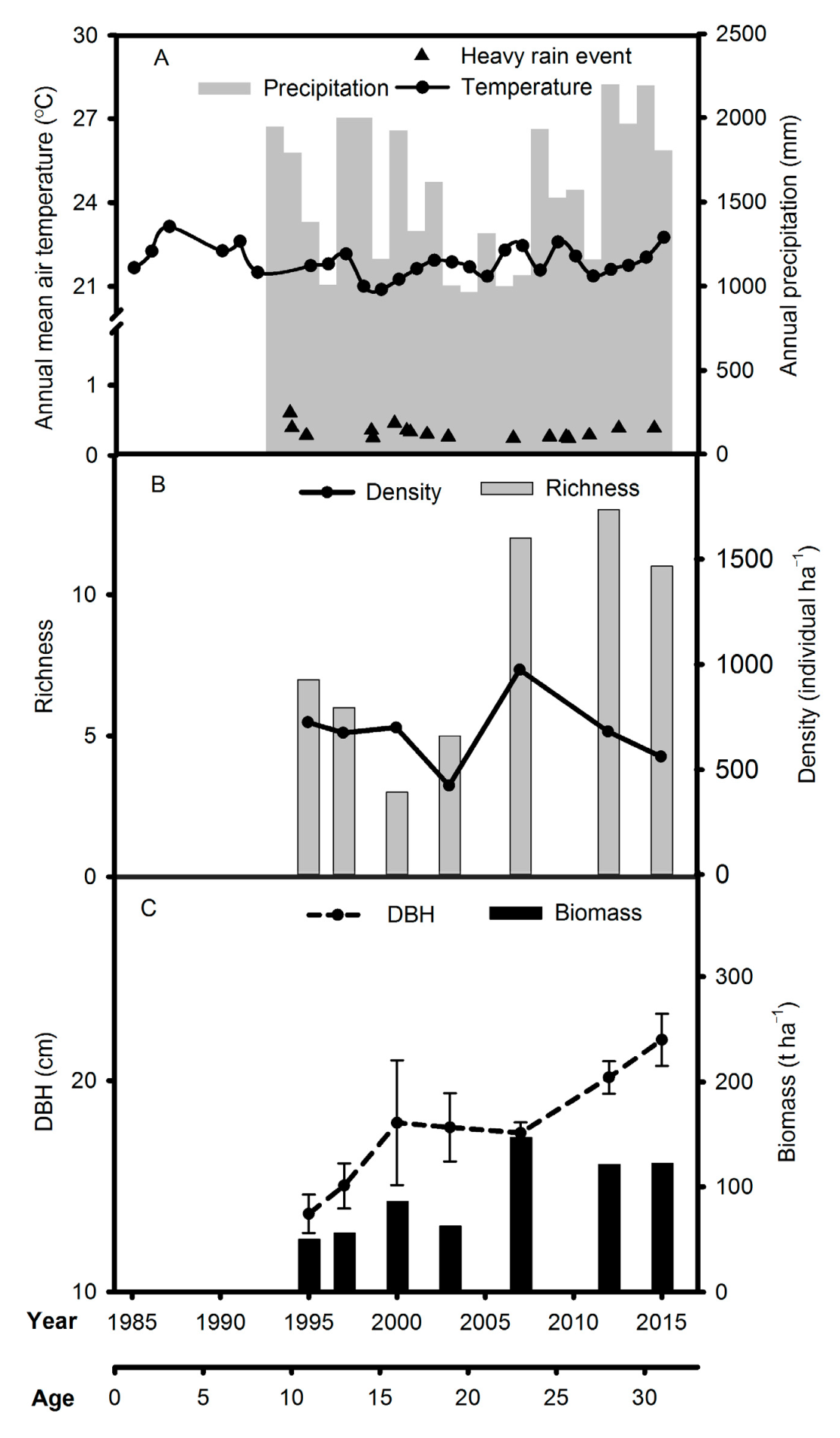
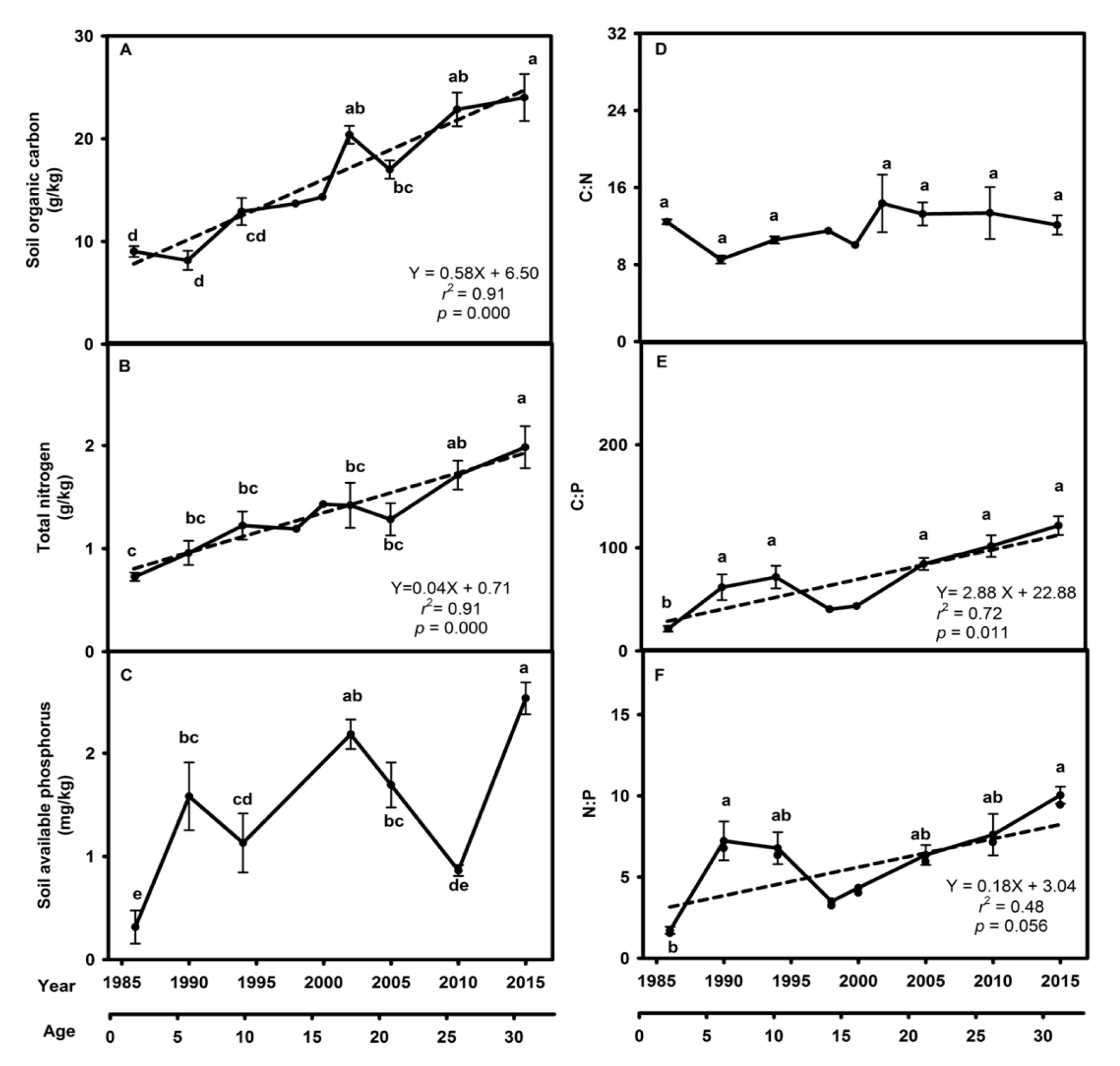
3.1. Tree Community Composition, Vegetation Biomass, and Soil and Foliar Properties in the A. mangium Monoculture
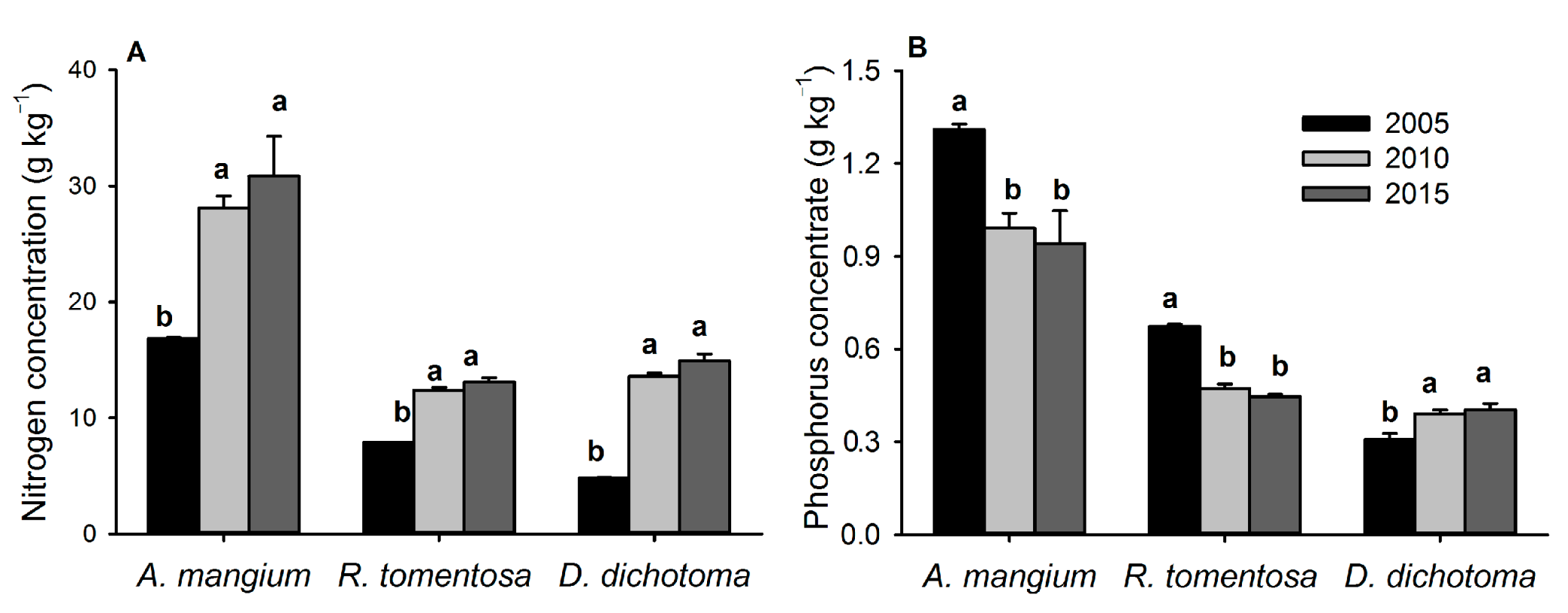
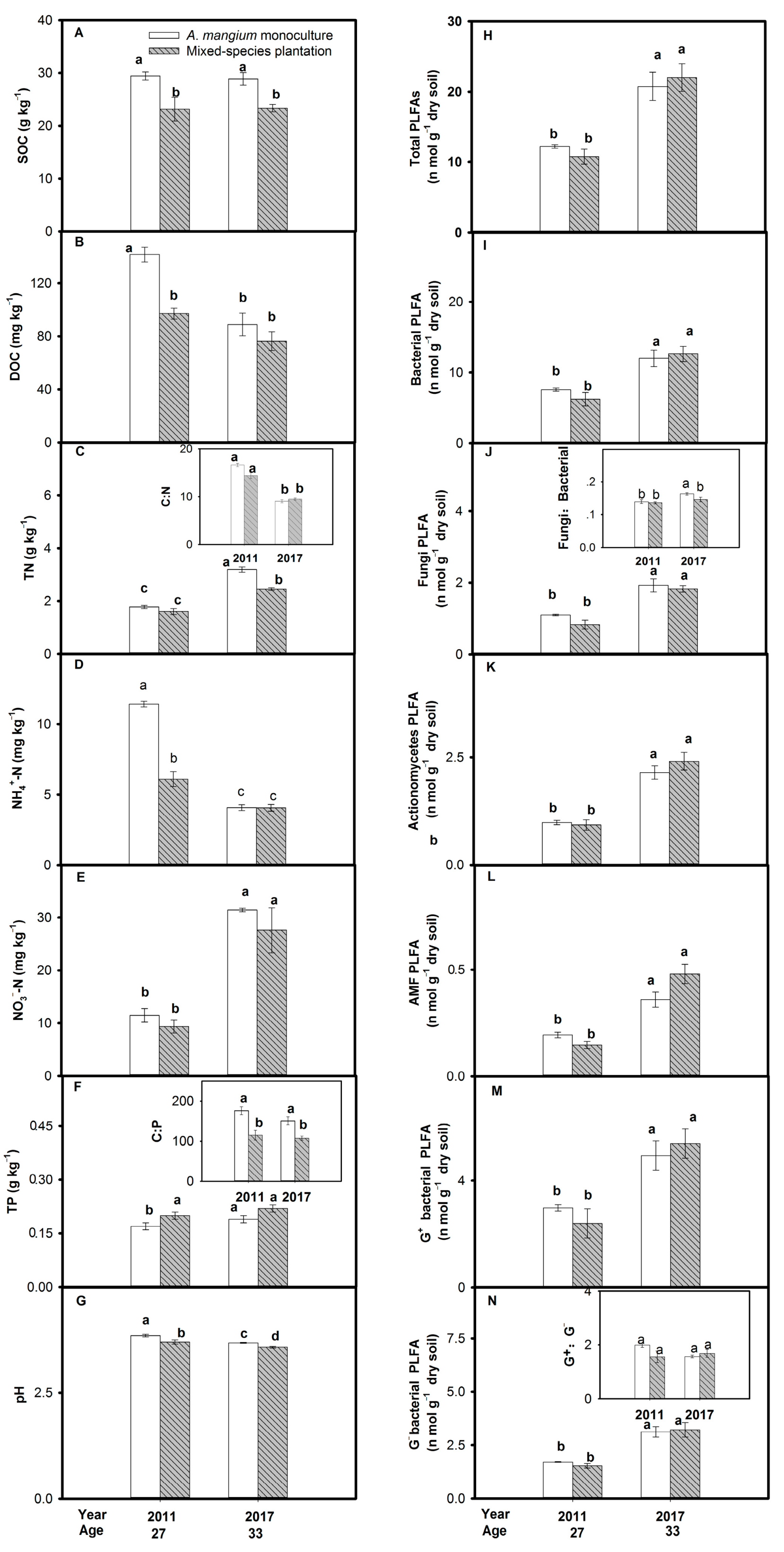
3.2. Comparison between the A. mangium Monoculture and the A. mangium Mixed-Species Plantation
4. Discussion
4.1. Dynamics of the Tree Community Composition, Vegetation Biomass, and Soil and Leaf Properties in the A. mangium Monoculture
4.2. Effects of Mixed-Species Plantation on Forest Stand Development
4.2.1. Changes in Tree Community Composition and Vegetation Biomass
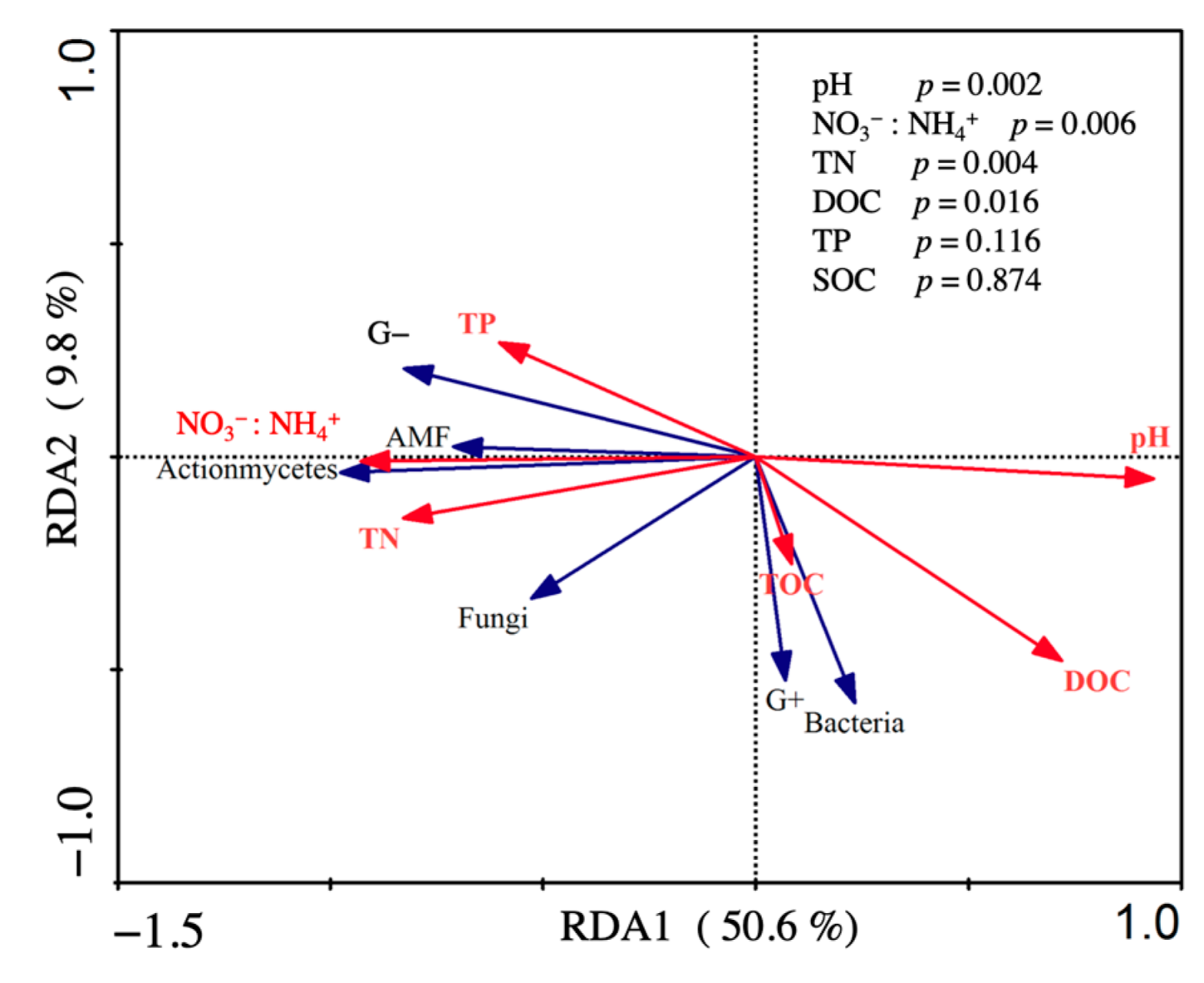
4.2.2. Changes in Soil Physiochemical Properties and Microbial Community Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World’s Forests: Forests, Biodiversity and People; FAO: Rome, Italy, 2020. [Google Scholar]
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, B.-J.; Stafford Smith, M.; Wang, S. Improve forest restoration initiatives to meet Sustainable Development Goal 15. Nat. Ecol. Evol. 2020, 5, 10–13. [Google Scholar] [CrossRef]
- Bauhus, J.; Forrester, D.I.; Pretzsch, H. From observations to evidence about effects of mixed-species stands. In Mixed-Species Forests; Springer: Berlin/Heidelberg, Germany, 2017; pp. 27–71. [Google Scholar]
- Messier, C.; Bauhus, J.; Sousa-Silva, R.; Auge, H.; Baeten, L.; Barsoum, N.; Bruelheide, H.; Caldwell, B.; Cavender-Bares, J.; Dhiedt, E. For the sake of resilience and multifunctionality, let’s diversify planted forests! Conserv. Lett. 2022, 15, e12829. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E.; Arneth, A.; Bai, X.; Barnes, D.; Burrows, M.; Chan, L.; Cheung, W.L.W.; et al. Scientific Outcome of the IPBES-IPCC Co-Sponsored Workshop on Biodiversity and Climate Change; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES): Bonn, Germany, 2021. [Google Scholar]
- Forrester, D.I.; Bauhus, J.; Khanna, P.K. Growth dynamics in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2004, 193, 81–95. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Bauhus, J. Mixed-species forests. In Ecology and management; Springer: Berlin, Germany, 2017. [Google Scholar]
- Erskine, P.D.; Lamb, D.; Bristow, M. Tree species diversity and ecosystem function: Can tropical multi-species plantations generate greater productivity? For. Ecol. Manag. 2006, 233, 205–210. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P.; et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Laclau, J.-P.; Gonçalves, J.L.d.M.; Voigtlaender, M.; Gava, J.L.; Leite, F.P.; Hakamada, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. On the success and failure of mixed-species tree plantations: Lessons learned from a model system of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]
- Voigtlaender, M.; Laclau, J.-P.; de Moraes Goncalves, J.L.; Piccolo, M.d.C.; Moreira, M.Z.; Nouvellon, Y.; Ranger, J.; Bouillet, J.-P. Introducing Acacia mangium trees in Eucalyptus grandis plantations: Consequences for soil organic matter stocks and nitrogen mineralization. Plant Soil 2012, 352, 99–111. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [PubMed]
- Amazonas, N.T.; Forrester, D.I.; Silva, C.C.; Almeida, D.R.A.; Rodrigues, R.R.; Brancalion, P.H.S. High diversity mixed plantations of Eucalyptus and native trees: An interface between production and restoration for the tropics. For. Ecol. Manag. 2018, 417, 247–256. [Google Scholar] [CrossRef]
- Manson, D.G.; Schmidt, S.; Bristow, M.; Erskine, P.D.; Vanclay, J.K. Species-site matching in mixed species plantations of native trees in tropical Australia. Agrofor. Syst. 2012, 87, 233–250. [Google Scholar] [CrossRef]
- Amazonas, N.T.; Forrester, D.I.; Silva, C.C.; de Almeida, D.R.A.; Oliveira, R.S.; Rodrigues, R.R.; Brancalion, P.H.S. Light- and nutrient-related relationships in mixed plantations of Eucalyptus and a high diversity of native tree species. New For. 2021, 52, 807–828. [Google Scholar] [CrossRef]
- Yu, Z.; Peng, S. The artificial and natural restoration of tropical and subtropical forest. Acta Ecol. Sin. 1995, 15 (Suppl. A), 1–17. [Google Scholar]
- Zhou, G.; Morris, J.; Yan, J.; Yu, Z.; Peng, S. Hydrological impacts of reafforestation with eucalypts and indigenous species: A case study in southern China. For. Ecol. Manag. 2002, 167, 209–222. [Google Scholar] [CrossRef]
- Peng, S.L. Study and Application of Restoration Ecology in Tropical and Subtropical China; Science Press: Beijing, China, 2003. [Google Scholar]
- Ren, H.; Shen, W.J.; Lu, H.F.; Wen, X.Y.; Jian, S.G. Degraded ecosystems in China: Status, causes, and restoration efforts. Landsc. Ecol. Eng. 2007, 3, 1–13. [Google Scholar] [CrossRef]
- Duan, W.; Ren, H.; Fu, S.; Wang, J.; Zhang, J.; Yang, L.; Huang, C. Community Comparison and Determinant Analysis of Understory Vegetation in Six Plantations in South China. Restor. Ecol. 2010, 18, 206–214. [Google Scholar] [CrossRef]
- Wang, J.; Zou, C.; Ren, H.; Duan, W. Absence of tree seeds impedes shrubland succession in southern China. J. Trop. For. Sci. 2009, 21, 210–217. [Google Scholar]
- Peng, S.L.; Liu, J.; Lu, H.F. Characteristics and role of Acacia auriculiformis on vegetation restoration in lower subtropics of China. J. Trop. For. Sci. 2005, 17, 508–525. [Google Scholar]
- Forrester, D.I.; Pares, A.; O’Hara, C.; Khanna, P.K.; Bauhus, J. Soil Organic Carbon is Increased in Mixed-Species Plantations of Eucalyptus and Nitrogen-Fixing Acacia. Ecosystems 2013, 16, 123–132. [Google Scholar] [CrossRef]
- Kaye, J.P.; Resh, S.C.; Kaye, M.W.; Chimner, R.A. Nutrient and Carbon Dynamics in a Replacement Series of Eucalyptus and Albizia Trees. Ecology 2000, 81, 3267–3273. [Google Scholar] [CrossRef]
- Forrester, D.I.; Pretzsch, H. Tamm Review: On the strength of evidence when comparing ecosystem functions of mixtures with monocultures. For. Ecol. Manag. 2015, 356, 41–53. [Google Scholar] [CrossRef]
- World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports 2014, Update 2015; FAO: Rome, Italy, 2015. [Google Scholar]
- Kalembasa, S.J.; Jenkinson, D.S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Liu, G.; Jiang, N.; Zhang, L.; Liu, Z. Soil Physical and Chemical Analysis and Description of Soil Profiles; China Standard Methods Press: Beijing, China, 1996. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Li, Z.; Peng, S. Nutrient structure of several man-made forests in tropical and subtropical of China. Chin. J. Ecol. 2001, 20, 1–4. [Google Scholar]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Methods 1991, 14, 151–163. [Google Scholar] [CrossRef]
- Frostegård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Johansen, A.; Olsson, S. Using Phospholipid Fatty Acid Technique to Study Short-Term Effects of the Biological Control Agent Pseudomonas fluorescens DR54 on the Microbial Microbiota in Barley Rhizosphere. Microb. Ecol. 2005, 49, 272–281. [Google Scholar] [CrossRef]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps in the forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Banerjee, S.; Siciliano, S.D.; Grogan, P. The seasonal pattern of soil microbial community structure in mesic low arctic tundra. Soil Biol. Biochem. 2013, 65, 338–347. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Experimental analysis of the effect of exotic and native plant species on the structure and function of soil microbial communities. Soil Biol. Biochem. 2003, 35, 895–905. [Google Scholar] [CrossRef]
- Kulmatiski, A.; Beard, K.H. Long-term plant growth legacies overwhelm short-term plant growth effects on soil microbial community structure. Soil Biol. Biochem. 2011, 43, 823–830. [Google Scholar] [CrossRef]
- McIntosh, A.C.S.; Macdonald, S.E.; Quideau, S.A. Linkages between the forest floor microbial community and resource heterogeneity within mature lodgepole pine forests. Soil Biol. Biochem. 2013, 63, 61–72. [Google Scholar] [CrossRef]
- Li, Z.A.; Peng, S.L.; Rae, D.J.; Zhou, G.Y. Litter decomposition and nitrogen mineralization of soils in subtropical plantation forests of southern China, with special attention to comparisons between legumes and non-legumes. Plant Soil 2001, 229, 105–116. [Google Scholar] [CrossRef]
- Zhou, G.; Wei, X.; Wu, Y.; Liu, S.; Huang, Y.; Yan, J.; Zhang, D.; Zhang, Q.; Liu, J.; Meng, Z.; et al. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China. Glob. Chang. Biol. 2011, 17, 3736–3746. [Google Scholar] [CrossRef]
- Laclau, J.P.; Bouillet, J.P.; Gonçalves, J.L.M.; Silva, E.V.; Jourdan, C.; Cunha, M.C.S.; Moreira, M.R.; Saint-André, L.; Maquère, V.; Nouvellon, Y.; et al. Mixed-species plantations of Acacia mangium and Eucalyptus grandis in Brazil. For. Ecol. Manag. 2008, 255, 3905–3917. [Google Scholar] [CrossRef]
- Santos, F.M.; Chaer, G.M.; Diniz, A.R.; Balieiro, F.d.C. Nutrient cycling over five years of mixed-species plantations of Eucalyptus and Acacia on a sandy tropical soil. For. Ecol. Manag. 2017, 384, 110–121. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Management of soil, nutrients and water in tropical plantation forests. In Nitrogen Fixation in Tropical Forest Plantations; Nambiar, E.K.S., Brown, A.G., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1997; pp. 297–337. [Google Scholar]
- Forrester, D.I.; Schortemeyer, M.; Stock, W.D.; Bauhus, J.; Khanna, P.K.; Cowie, A.L. Assessing nitrogen fixation in mixed- and single-species plantations of Eucalyptus globulus and Acacia mearnsii. Tree Physiol. 2007, 27, 1319–1328. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Deng, M.; Liu, L.; Jiang, L.; Liu, W.; Wang, X.; Li, S.; Yang, S.; Wang, B. Ecosystem scale trade-off in nitrogen acquisition pathways. Nat. Ecol. Evol. 2018, 2, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liu, J.; Wang, Y.P.; Zhou, G.; Han, T.; Li, Y. Increasing phosphorus limitation along three successional forests in southern China. Plant Soil 2013, 364, 181–191. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, G.; Luo, X.; Hou, E.; Zheng, M.; Zhang, L.; He, X.; Shen, W.; Wen, D. Mycorrhizal fungi and phosphatase involvement in rhizosphere phosphorus transformations improves plant nutrition during subtropical forest succession. Soil Biol. Biochem. 2021, 153, 108099. [Google Scholar] [CrossRef]
- Sardans, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Peñuelas, J. Recent advances and future research in ecological stoichiometry. Perspect. Plant Ecol. Evol. Syst. 2021, 50, 125611. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of phosphatase enzymes in soil. In Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar]
- Zhang, L.; Xu, M.; Liu, Y.; Zhang, F.; Hodge, A.; Feng, G. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 2016, 210, 1022–1032. [Google Scholar] [CrossRef]
- Yang, L.; Liu, N.; Ren, H.; Wang, J. Facilitation by two exotic Acacia: Acacia auriculiformis and Acacia mangium as nurse plants in South China. For. Ecol. Manag. 2009, 257, 1786–1793. [Google Scholar] [CrossRef]
- Norisada, M.; Hitsuma, G.; Kuroda, K.; Yamanoshita, T.; Masumori, M.; Tange, T.; Yagi, H.; Nuyim, T.; Sasaki, S.; Kojima, K. Acacia mangium, a Nurse Tree Candidate for Reforestation on Degraded Sandy Soils in the Malay Peninsula. For. Sci. 2005, 51, 498–510. [Google Scholar]
- Jones, E.R.; Wishnie, M.H.; Deago, J.; Sautu, A.; Cerezo, A. Facilitating natural regeneration in Saccharum spontaneum (L.) grasslands within the Panama Canal Watershed: Effects of tree species and tree structure on vegetation recruitment patterns. For. Ecol. Manag. 2004, 191, 171–183. [Google Scholar] [CrossRef]
- Wang, X.; Hua, F.; Wang, L.; Wilcove, D.S.; Yu, D.W. The biodiversity benefit of native forests and mixed-species plantations over monoculture plantations. Divers. Distrib. 2019, 25, 1721–1735. [Google Scholar] [CrossRef]
- Feng, Y.; Schmid, B.; Loreau, M.; Forrester, D.I.; Fei, S.; Zhu, J.; Tang, Z.; Zhu, J.; Hong, P.; Ji, C. Multispecies forest plantations outyield monocultures across a broad range of conditions. Science 2022, 376, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Wallander, H.; Ekblad, A.; Godbold, D.; Johnson, D.; Bahr, A.; Baldrian, P.; Björk, R.; Kieliszewska-Rokicka, B.; Kjøller, R.; Kraigher, H. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils–A review. Soil Biol. Biochem. 2013, 57, 1034–1047. [Google Scholar] [CrossRef]
- Frostegård, Å.; Bååth, E.; Tunlio, A. Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol. Biochem. 1993, 25, 723–730. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, T.; Gilliam, F.S.; Gundersen, P.; Zhang, W.; Chen, H.; Mo, J. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest. PLoS ONE 2013, 8, e61188. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.L.; Lin, Y.B.; Rao, X.Q.; Liu, S.P. Forest Ecosystem: Heshan Filed Station in Guangdong; Chinese Forestry Press: Beijing, China, 2008. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | A. mangium Monoculture Plots | Mixed-Species Plantation of A. mangium Plots | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | A. mangium | Regenerated Species | Total | A. mangium | Planted Native Species | Regenerated Species | ||||||||
| 2011 | 2017 | 2011 | 2017 | 2011 | 2017 | 2011 | 2017 | 2011 | 2017 | 2011 | 2017 | 2011 | 2017 | |
| Density (individual ha−1) | 833 | 634 | 700 | 467 | 133 | 167 | 1287 | 1233 | 125 | 108 | 775 | 900 | 387 | 225 |
| Biomass (t ha−1) | 118.87 | 108.69 | 118.76 | 108.58 | 0.10 | 0.10 | 50.06 | 103.17 | 24.73 | 46.04 | 9.06 | 38.29 | 16.81 | 18.77 |
| Tree species richness | 6 | 7 | 1 | 1 | 5 | 6 | 39 | 38 | 1 | 1 | 30 | 30 | 8 | 7 |
| Mean tree DBH (cm) | 14.87 ± 0.89 | 17.54 ± 1.44 | 17.14 ± 0.86 | 21.19 ± 1.39 | 2.98 ± 0.40 | 2.93 ± 0.50 | 7.83 ± 0.64 | 10.00 ± 0.82 | 20.21 ± 2.25 | 31.44 ± 3.35 | 6.66 ± 0.67 | 9.31 ± 0.38 | 2.17 ± 0.24 | 2.45 ± 0.32 |
| Mean tree height (m) | 14.23 ± 0.65 | 12.67 ± 0.79 | 16.39 ± 0.49 | 15.14 ± 0.65 | 2.86 ± 0.30 | 2.81 ± 0.36 | 7.67 ± 0.53 | 8.52 ± 0.46 | 15.40 ± 1.59 | 17.00 ± 0.74 | 7.28 ± 0.65 | 8.67 ± 0.27 | 3.64 ± 0.16 | 3.47 ± 0.33 |
| Shrub and herb richness | 20 | 13 | 28 | 21 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, S.; Tie, L.; Rao, X.; Cai, X.; Liu, S.; Vitali, V.; Wei, L.; Yu, Q.; Sun, D.; Lin, Y.; et al. Mixed-Species Acacia Plantation Decreases Soil Organic Carbon and Total Nitrogen Concentrations but Favors Species Regeneration and Tree Growth over Monoculture: A Thirty-Three-Year Field Experiment in Southern China. Forests 2023, 14, 968. https://doi.org/10.3390/f14050968
Ouyang S, Tie L, Rao X, Cai X, Liu S, Vitali V, Wei L, Yu Q, Sun D, Lin Y, et al. Mixed-Species Acacia Plantation Decreases Soil Organic Carbon and Total Nitrogen Concentrations but Favors Species Regeneration and Tree Growth over Monoculture: A Thirty-Three-Year Field Experiment in Southern China. Forests. 2023; 14(5):968. https://doi.org/10.3390/f14050968
Chicago/Turabian StyleOuyang, Shengnan, Liehua Tie, Xingquan Rao, Xi’an Cai, Suping Liu, Valentina Vitali, Lanying Wei, Qingshui Yu, Dan Sun, Yongbiao Lin, and et al. 2023. "Mixed-Species Acacia Plantation Decreases Soil Organic Carbon and Total Nitrogen Concentrations but Favors Species Regeneration and Tree Growth over Monoculture: A Thirty-Three-Year Field Experiment in Southern China" Forests 14, no. 5: 968. https://doi.org/10.3390/f14050968
APA StyleOuyang, S., Tie, L., Rao, X., Cai, X., Liu, S., Vitali, V., Wei, L., Yu, Q., Sun, D., Lin, Y., Bose, A. K., Gessler, A., & Shen, W. (2023). Mixed-Species Acacia Plantation Decreases Soil Organic Carbon and Total Nitrogen Concentrations but Favors Species Regeneration and Tree Growth over Monoculture: A Thirty-Three-Year Field Experiment in Southern China. Forests, 14(5), 968. https://doi.org/10.3390/f14050968