
Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection and Chronology Development
2.3. Climate Data
2.4. Data Analysis
3. Results
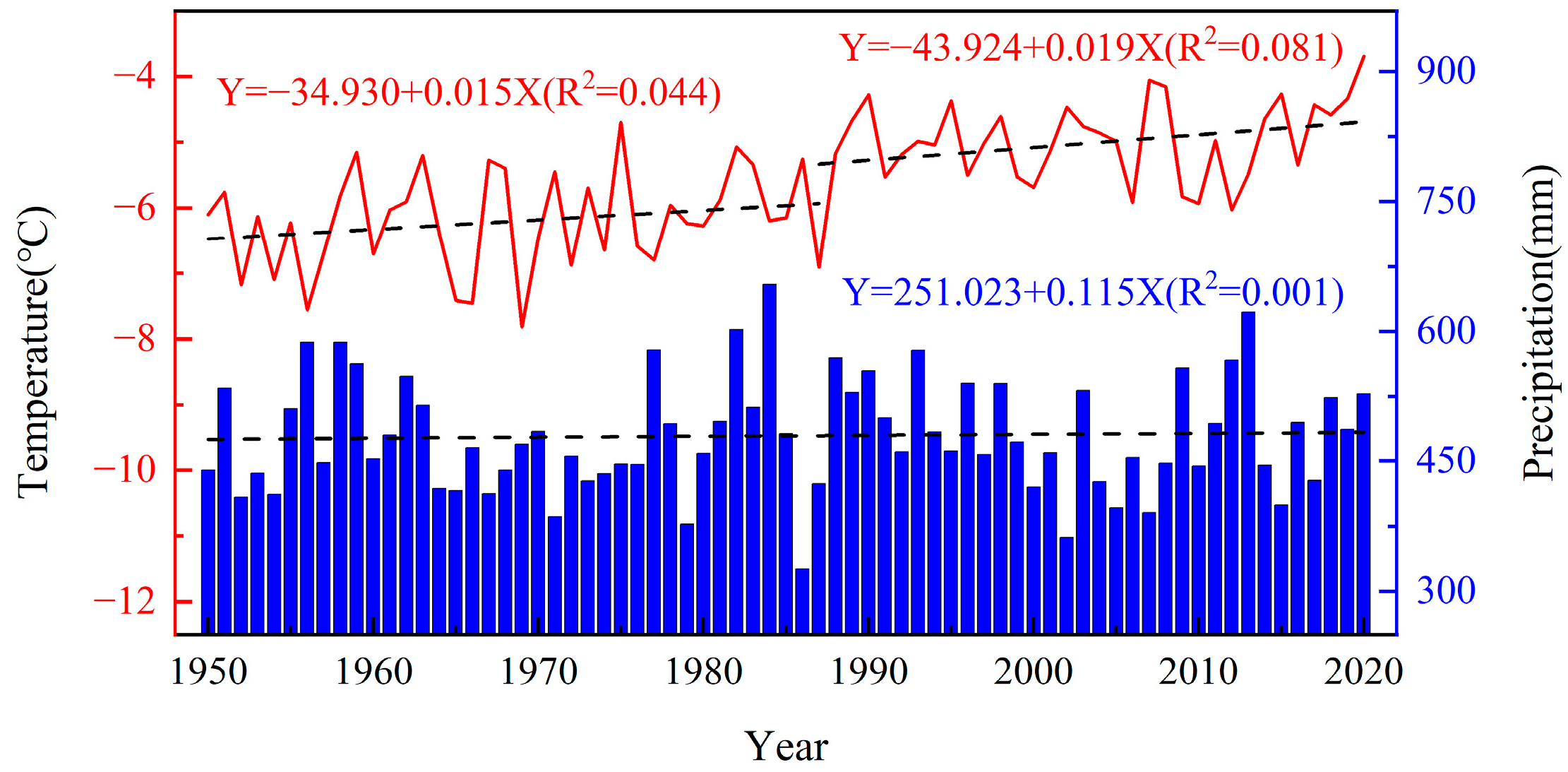
3.1. Winter Climatic Characteristics
3.2. Radial Growth Characteristics
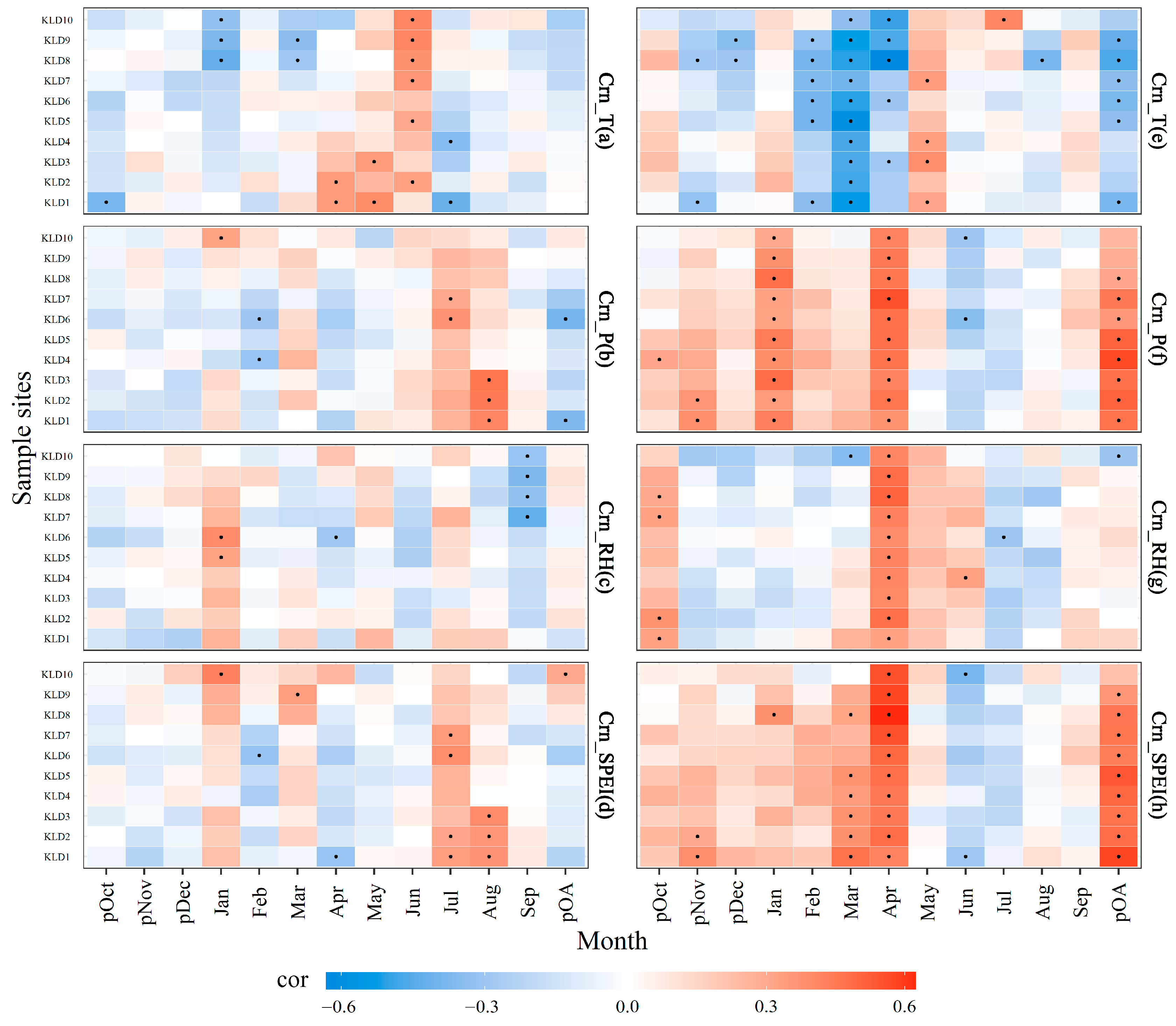
3.3. Growth–Climate Relationships
3.4. Altitudinal Divergence of Growth–Winter Drought Associations
4. Discussion
4.1. Temporal–Spatial Differences in Radial Growth
4.2. Temporal–Spatial Differences in Growth–Climate Associations
4.3. Future Winter Warming-Drying Trends and Dahurian larch Distribution
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Press: Cambridge, UK, 2021. [Google Scholar]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Press: Cambridge, UK, 2022. [Google Scholar]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, C.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368, eaaz7005. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wu, C.; Acil, N.; Carvalhais, N.; Pugh, T.A.M.; Sadler, J.P.; Seidl, R. A climate risk analysis of Earth’s forests in the 21st century. Science 2022, 377, 1099–1103. [Google Scholar] [CrossRef]
- Du, H.; Liu, J.; Li, M.H.; Buntgen, U.; Yang, Y.; Wang, L.; Wu, Z.; He, H.S. Warming-induced upward migration of the alpine treeline in the Changbai Mountains, northeast China. Glob. Chang. Biol. 2018, 24, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Gaire, N.P.; Fan, Z.X.; Bräuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Q.; Xie, F.; He, N.; He, R.; Zhang, K.; Zhang, Q.; Dang, H. Disparity in elevational shifts of upper species limits in response to recent climate warming in the Qinling Mountains, North-central China. Sci. Total Environ. 2020, 706, 135718. [Google Scholar] [CrossRef]
- Zheng, L.; Gaire, N.P.; Shi, P.; Schöb, C. High-altitude tree growth responses to climate change across the Hindu Kush Himalaya. J. Plant Ecol. 2021, 14, 829–842. [Google Scholar] [CrossRef]
- Yuan, S.; Jiang, Y.; Zhao, Z.; Cui, M.; Shi, D.; Wang, S.; Kang, M. Different trends and divergent responses to climate factors in the radial growth of Abies georgei along elevations in the central Hengduan Mountains. Dendrochronologia 2023, 80, 126114. [Google Scholar] [CrossRef]
- Panthi, S.; Fan, Z.X.; van der Sleen, P.; Zuidema, P.A. Long-term physiological and growth responses of Himalayan fir to environmental change are mediated by mean climate. Glob. Chang. Biol. 2020, 26, 1778–1794. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Zhang, W.T.; Wang, M.C.; Zhang, L.N.; Zhao, S.D. Divergent responses to climate factors in the radial growth of Larix sibirica in the eastern Tianshan Mountains, northwest China. Trees 2015, 29, 1673–1686. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, Q.B.; Pellatt, M.G.; Büntgen, U.; Li, M.H.; Cherubini, P. Drought limitation on tree growth at the Northern Hemisphere’s highest tree line. Dendrochronologia 2019, 53, 40–47. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Y. Climate warming and land use change in Heilongjiang Province, Northeast China. Appl. Geogr. 2011, 31, 476–482. [Google Scholar] [CrossRef]
- Yasmeen, S.; Wang, X.; Zhao, H.; Zhu, L.; Yuan, D.; Li, Z.; Zhang, Y.; Ahmad, S.; Han, S. Contrasting climate-growth relationship between Larix gmelinii and Pinus sylvestris var. mongolica along a latitudinal gradient in Daxing’an Mountains, China. Dendrochronologia 2019, 58, 125645. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhang, Q.; Zeng, X.; Xu, G.; Wu, G.; Wang, W. Species-specific tree growth and intrinsic water-use efficiency of Dahurian larch (Larix gmelinii) and Mongolian pine (Pinus sylvestris var. mongolica) growing in a boreal permafrost region of the Greater Hinggan Mountains, Northeastern China. Agric. For. Meteorol. 2018, 248, 145–155. [Google Scholar] [CrossRef]
- Zhang, X.; Bai, X.; Hou, M.; Chen, Z.; Manzanedo, R.D. Warmer Winter Ground Temperatures Trigger Rapid Growth of Dahurian Larch in the Permafrost Forests of Northeast China. J. Geophys. Res. Biogeosci. 2019, 124, 1088–1097. [Google Scholar] [CrossRef]
- Bai, X.P.; Zhang, X.L.; Li, J.X.; Duan, X.Y.; Jin, Y.T.; Chen, Z.J. Altitudinal disparity in growth of Dahurian larch (Larix gmelinii Rupr.) in response to recent climate change in northeast China. Sci. Total Environ. 2019, 670, 466–477. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Zhu, X.; Yue, Y.; Xue, J.; Shi, L. How permafrost degradation threatens boreal forest growth on its southern margin? Sci. Total Environ. 2021, 762, 143154. [Google Scholar] [CrossRef]
- Pithan, F.; Mauritsen, T. Arctic amplification dominated by temperature feedbacks in contemporary climate models. Nat. Geosci. 2014, 7, 181–184. [Google Scholar] [CrossRef]
- Gao, M.; Meng, F. On the delineation of forest vegetation zones in the Oakleigh Pile Mountains. Inn. Mong. For. Investig. Des. 1999, 4, 141–143. [Google Scholar]
- Zhang, L. The Research on the East Slope of Middle Region of Daxing’an Mountains Vertical Zone. Master’s Thesis, Northeast Normal University, Changchun, China, 2007. [Google Scholar]
- Huang, X.; Zhang, X.; Yuan, L. Vegetation Types and Vertical Distribution Patterns of Oakley Mountain. High. Educ. Forum. 2009. Available online: https://www.docin.com/p-236712927.html (accessed on 20 February 2024).
- Yang, Z.; Zhou, G.; Yin, X.; Jia, B. Geographic distribution of Larix gmelinii natural forest in China and its climatic suitability. Chin. J. Ecol. 2014, 33, 1429–1436. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, S.; Song, W.; Tigabu, M.; Fu, M.; Xue, H.; Sun, A.; Zhao, M.; Cai, K.; Li, Y.; et al. Climate response of radial growth and early selection of Larix olgensis at four trials in northeast China. Dendrochronologia 2022, 73, 125955. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Cook, E.R. A Time Series Approach to Tree-Ring Standardization. Ph.D. Thesis, The University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Holmes, R.L.; Cook, E.R. Guide for Computer Program ARSTAN; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1994. [Google Scholar]
- Peixoto, J.; Oort, A.H. The Climatology of Relative Humidity in the Atmosphere. J. Clim. 1996, 9, 3443–3463. [Google Scholar] [CrossRef]
- Liu, J.; Guo, H.; Li, Y.; Liu, H.; Wu, B. Fundamentals of Physical Quantity Calculation in Weather Analysis and Forecasting; China Meteorological Press: Beijing, China, 2005. [Google Scholar]
- Kendall, M.G. Rank Correlation Methods. Biometrika 1957, 44, 146–163. [Google Scholar] [CrossRef]
- Team R Core. R: A language and environment for statistical computing. MSOR Connect. 2014, 1, 12–21. [Google Scholar]
- Hepting, G.H. Climate and Forest diseases. Annu. Rev. Phytopathol. 1963, 60, 133–149. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, H.; Liu, X.; Liang, E.; Grießinger, J.; Wu, G.; Li, X.; Bräuning, A. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural alpine timberline on the southeastern Tibetan Plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- Su, J.; Gou, X.; Deng, Y.; Zhang, R.; Liu, W.; Zhang, F.; Lu, M.; Chen, Y.; Zheng, W. Tree growth response of Fokienia hodginsii to recent climate warming and drought in southwest China. Int. J. Biometeorol. 2017, 61, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Bogachev, M.I.; Gafurov, A.M.; Iskandirov, P.Y.; Kaplun, D.I.; Kayumov, A.R.; Lyanova, A.I.; Pyko, N.S.; Pyko, S.A.; Safonova, A.N.; Sinitca, A.M.; et al. Reversal in the drought stress response of the Scots pine forest ecosystem: Local soil water regime as a key to improving climate change resilience. Heliyon 2023, 9, e21574. [Google Scholar] [CrossRef]
- Freeland, R.O. Apparent Photosynthesis in Some Conifers During Winter. Plant Physiol. 1944, 19, 179–185. [Google Scholar] [CrossRef]
- Iversen, C.M.; Childs, J.; Norby, R.J.; Ontl, T.A.; Kolka, R.K.; Brice, D.J.; McFarlane, K.J.; Hanson, P.J. Fine-root growth in a forested bog is seasonally dynamic, but shallowly distributed in nutrient-poor peat. Plant Soil 2017, 424, 123–143. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Jiang, Y.; Wang, B.; Jiao, L.; Wang, M.c. Seasonal water use by Larix principis-rupprechtii in an alpine habitat. For. Ecol. Manag. 2018, 409, 47–55. [Google Scholar] [CrossRef]
- Sugimoto, A.; Naito, D.; Yanagisawa, N.; Ichiyanagi, K.; Kurita, N.; Kubota, J.; Kotake, T.; Ohata, T.; Maximov, T.C.; Fedorov, A.N. Characteristics of soil moisture in permafrost observed in East Siberian taiga with stable isotopes of water. Hydrol. Process. 2002, 17, 1073–1092. [Google Scholar] [CrossRef]
- Pompa García, M.; Cerano Paredes, J.; Fulé, P.Z. Variation in radial growth of Pinus cooperi in response to climatic signals across an elevational gradient. Dendrochronologia 2013, 31, 198–204. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Penuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Sun, Y. Effects of altitude and slope on the climate–radial growth relationships of Larix olgensis A. Henry in the southern Lesser Khingan Mountains, Northeast China. Ecol. Process. 2022, 11, 46. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Liu, C.; Guo, D.; Zhang, Q. Response of Chinese pine regeneration density to forest gap and slope aspect in northern China: A meta-analysis. Sci. Total Environ. 2023, 873, 162428. [Google Scholar] [CrossRef] [PubMed]
- Gaire, N.P.; Zaw, Z.; Bräuning, A.; Grießinger, J.; Sharma, B.; Rana, P.; Bhandari, S.; Basnet, S.; Fan, Z.-X. The impact of warming climate on Himalayan silver fir growth along an elevation gradient in the Mt. Everest region. Agric. For. Meteorol. 2023, 339, 109575. [Google Scholar] [CrossRef]
- Zhang, W.; Gou, X.; Zhang, F.; Liu, W.; Zhang, Y.; Gao, L. Divergent responses of Qinghai spruce (Picea crassifolia) to recent warming along elevational gradients in the central Qilian Mountains, Northwest China. J. Geogr. Sci. 2023, 33, 151–168. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Altitudinal Classes | Site ID | Time Span | Trees (Cores) | MS | SD | EPS | SNR |
|---|---|---|---|---|---|---|---|
| 970–1009 | KLD1 | 1880–2021 | 7 (14) | 0.267 | 0.507 | 0.970 | 31.907 |
| 1010–1049 | KLD2 | 1849–2021 | 13 (26) | 0.181 | 0.312 | 0.934 | 14.205 |
| 1050–1089 | KLD3 | 1849–2021 | 8 (16) | 0.231 | 0.416 | 0.859 | 6.112 |
| 1090–1129 | KLD4 | 1845–2021 | 13 (26) | 0.194 | 0.366 | 0.920 | 11.491 |
| 1130–1169 | KLD5 | 1873–2021 | 8 (16) | 0.343 | 0.223 | 0.851 | 5.695 |
| 1170–1209 | KLD6 | 1880–2021 | 5 (11) | 0.252 | 0.377 | 0.914 | 10.644 |
| 1210–1249 | KLD7 | 1794–2021 | 10 (20) | 0.258 | 0.439 | 0.925 | 12.346 |
| 1250–1289 | KLD8 | 1745–2021 | 12 (24) | 0.220 | 0.282 | 0.928 | 12.795 |
| 1290–1329 | KLD9 | 1811–2021 | 8 (16) | 0.251 | 0.314 | 0.920 | 11.484 |
| 1330–1409 | KLD10 | 1769–2021 | 3 (5) | 0.292 | 0.270 | 0.675 | 2.073 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Bai, X.; Tian, X.; Chen, Z.; Zhang, H.; Liu, H. Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China. Forests 2024, 15, 565. https://doi.org/10.3390/f15030565
Zhang R, Bai X, Tian X, Chen Z, Zhang H, Liu H. Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China. Forests. 2024; 15(3):565. https://doi.org/10.3390/f15030565
Chicago/Turabian StyleZhang, Rui, Xueping Bai, Xun Tian, Zhenju Chen, Haiyue Zhang, and Haiting Liu. 2024. "Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China" Forests 15, no. 3: 565. https://doi.org/10.3390/f15030565
APA StyleZhang, R., Bai, X., Tian, X., Chen, Z., Zhang, H., & Liu, H. (2024). Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China. Forests, 15(3), 565. https://doi.org/10.3390/f15030565