
Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Treatments
2.2. Determination of Pigment Content in Leaves
2.3. Determination of Endogenous ALA Content and ALA Biosynthesis and Metabolism in Leaves
2.4. Determination of Photosynthetic Gas Exchange Parameters
2.5. Determination of Chlorophyll Fast Fluorescence
2.6. RNA Extraction and RT-qPCR Analysis
2.7. DNA Extraction and Promoter Sequence Cloning from Plant Materials
2.8. Construction of Plant Gene Expression Vectors
2.9. Agrobacterium Transformation, Tobacco Infection and GUS Qualitative and Quantitative Experiments
2.10. Data Analysis
3. Results
3.1. Effects of Exogenous ALA Treatment on Photosynthetic Gas Exchange Parameters in Pear Leaves
3.2. Effects of Exogenous ALA Treatment on Chlorophyll Fluorescence Characteristics of Pear Leaves
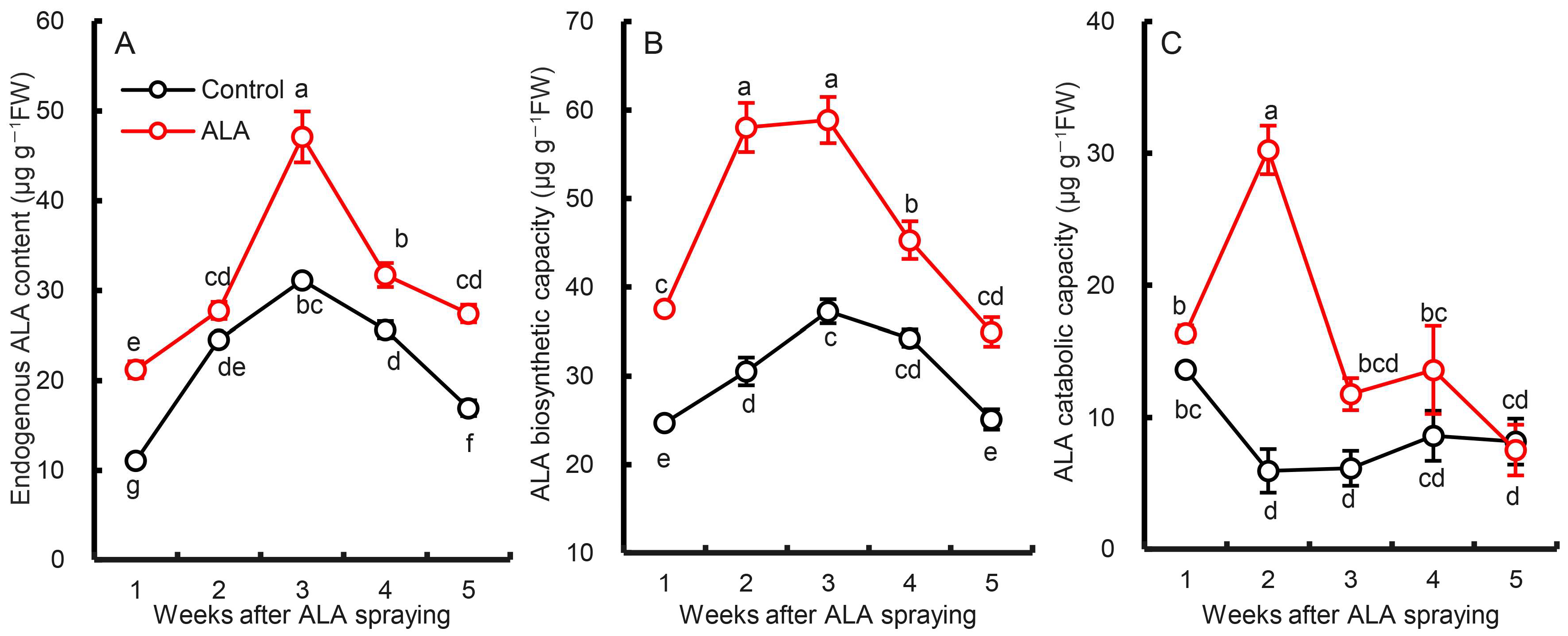
3.3. Effects of Exogenous ALA Treatment on Endogenous ALA Biosynthesis and Metabolism in Pear Leaves
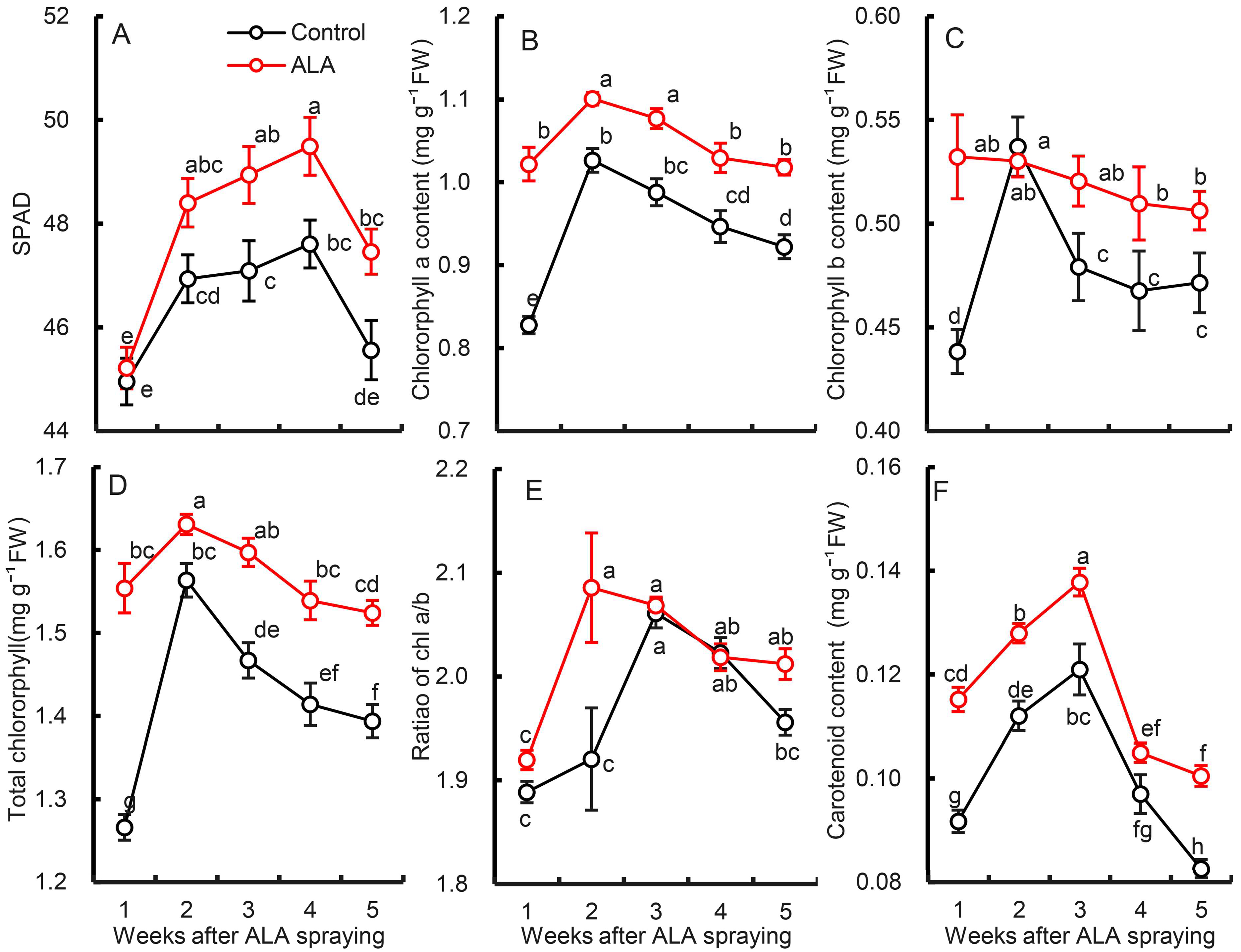
3.4. Effects of Exogenous ALA Treatment on Photosynthetic Pigment Content in Pear Leaves
3.5. Effects of Exogenous ALA Treatment on the Gene Expressions Related with Chlorophyll Synthesis
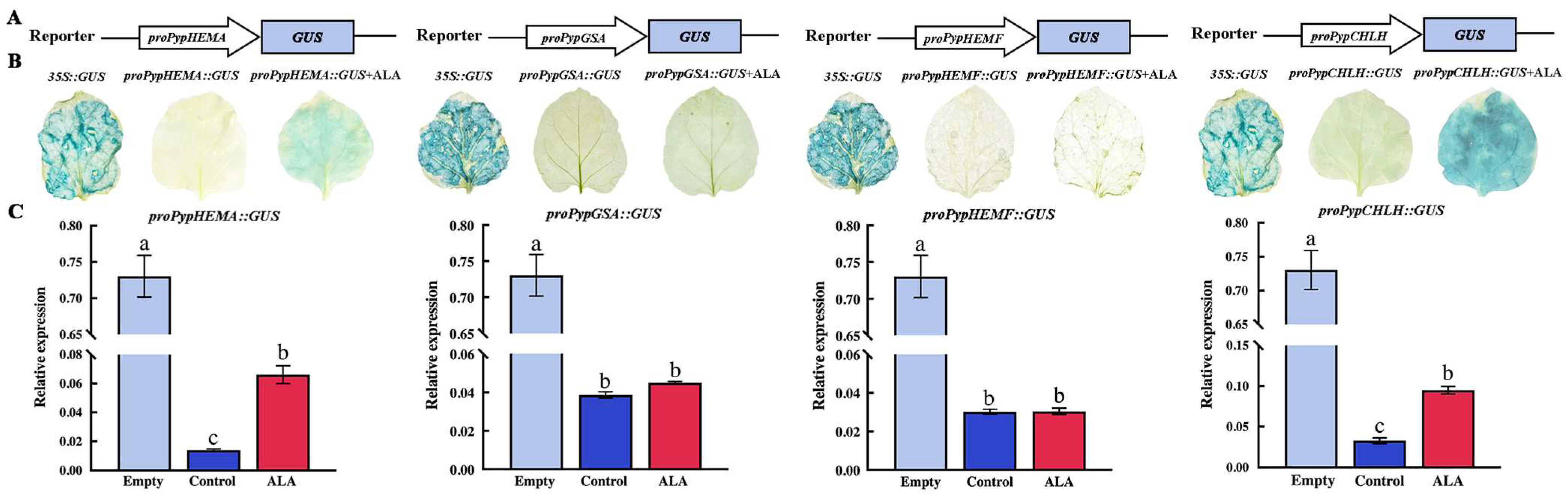
3.6. Effect of Exogenous ALA Treatment on Promoter Activity of Genes Related to ALA and Chlorophyll Synthesis
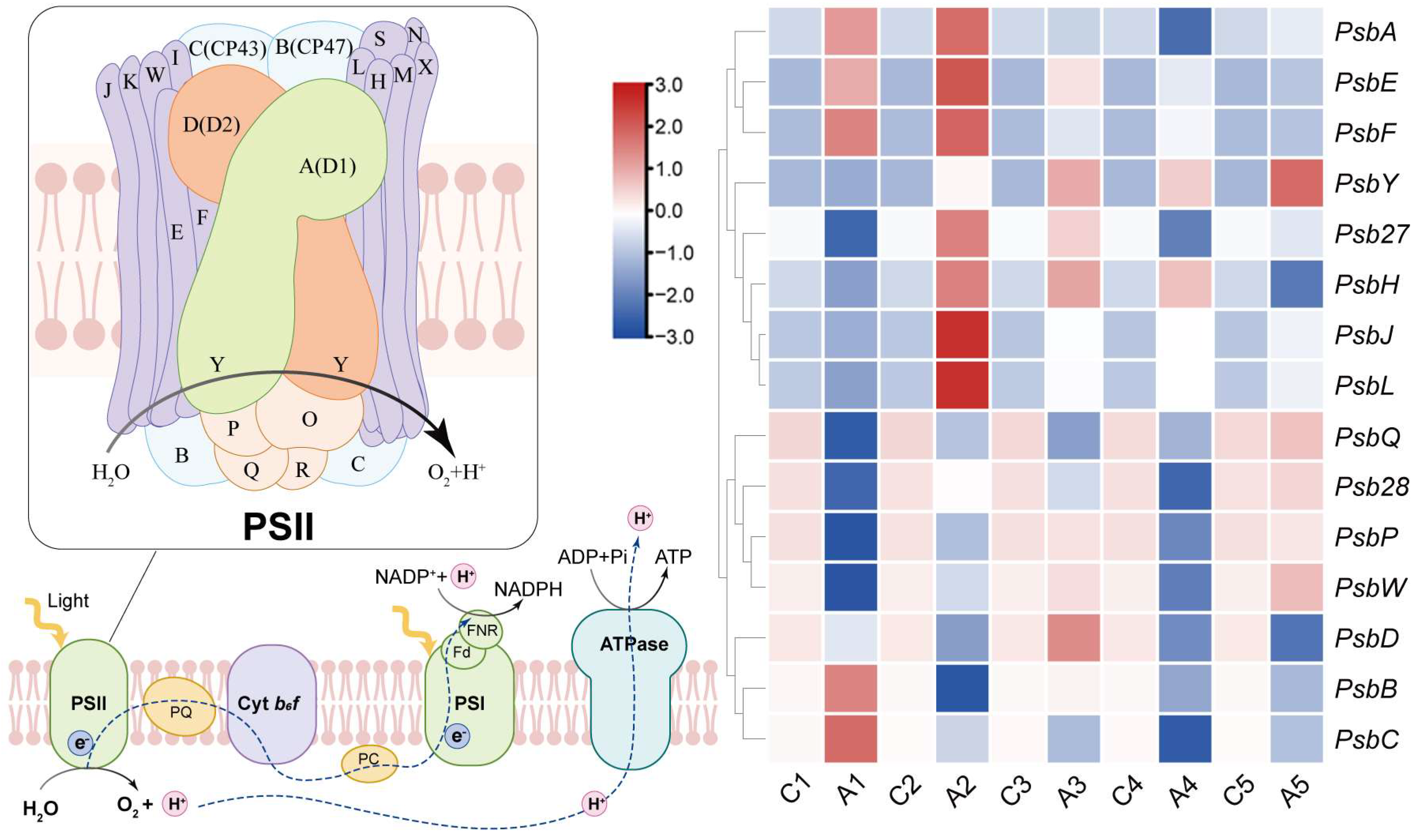
3.7. Expression Analysis of PSII-Related Genes in Pear Leaves Treated with Exogenous ALA
3.8. Effect of Exogenous ALA Treatment on Promoter Activity of PSII-Related Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Z.; Zhang, J.; Chen, Z.G. Analysis of the value chain of China’s pear fruit industry. Chin. Rural. Econ. 2008, 1, 63–72. [Google Scholar]
- Xu, D. Photosynthetic rate, photosynthetic efficiency, and crop yield. Bull. Biol. 1999, 1, 11–13. [Google Scholar]
- Peng, L.L.; Jiang, W.B.; Han, J. Effects of source-sink relationship change on yield and quality in fruit tree. Nonwood For. Res. 2012, 30, 134–140. [Google Scholar]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Wang, L.J.; Jiang, W.B.; Liu, H.; Liu, W.Q.; Kagn, L.; Hou, X.L. Promotion by 5-aminolevulinic acid of germination of pakchoi (Brassica campestris ssp. chinensis var. communis Tsen et Lee) seeds under salt stress. J. Integra Plant Biol. 2005, 47, 1084–1091. [Google Scholar] [CrossRef]
- Lukšienė, Ž.; Danilčenko, H.; Tarasevičienė, Ž.; Anusevičius, Ž.; Marozienė, A.; Nivinskas, H. New approach to the fungal decontamination of wheat used for wheat sprouts: Effects of aminolevulinic acid. Int. J. Food Microbiol. 2007, 116, 153–158. [Google Scholar] [CrossRef]
- Mai, H.X.; Wang, H.X.; Hu, Y.P.; Han, W.Y.; Li, Z.M.; Pang, Z.P.; Zhou, H.G. Effects of growth regulator ALA on growth and physiology traits of potted lily. Jiangsu Agric. Sci. 2021, 49, 9–103. [Google Scholar]
- Kong, L.G.; Wang, Y.H.; Han, X.D.; Bo, H.X.; Xun, J.; Wang, L.J. Effect of Cropspring foliar fertilizer on growth and quality of rice under different nitrogen supplies. Jiangsu J. Agric. Sci. 2018, 34, 90–798. [Google Scholar]
- Li, M.A.; Ma, L.; Hao, Q.; An, Y.Y.; Wang, L.J. Effect of 5-aminolevulinic acid on leaf photosynthetic characteristics, yield, and quality of potato. China Veg. 2020, 1, 43–52. [Google Scholar]
- Sheng, Y.F.; Yang, L.L.; Wang, J.K.; Chen, Y. Effects of 5-aminolevulinic acid on yield and benefit of carrot. Hortic. Seed 2020, 40, 8–9+11. [Google Scholar]
- Gao, J.J.; Feng, X.X.; Duan, C.H.; Li, J.H.; Shi, Z.X.; Gao, C.Y.; Wang, L.J. Effects of 5-aminolevulinic acid (ALA) on leaf photosynthesis and fruit quality of apples. J. Fruit Sci. 2013, 30, 944–951. [Google Scholar]
- Xie, L.; Cheng, X.H.; Feng, X.X.; Yang, T.; Zhang, Z.P.; Wang, L.J. Effects of an amino acid fertilizer on the leaf photosynthesis and fruit quality of ‘Summer black’ grape. J. Nanjing Agric. Univ. 2013, 36, 31–37. [Google Scholar]
- Wei, J.G.; Yu, H.; Zhang, X.; Jiang, Y.Q.; Zeng, Q.L. Effects of 5-aminolevulinic acid on photosynthetic capacity, yield, and fruit quality of Vaccinium ashei Reade. North. Hortic. 2014, 16, 9–12. [Google Scholar]
- Fang, X.; Zhang, L.Z.; Wang, L.J. The transcription factor MdERF78 is involved in ALA-induced anthocyanin accumulation in apples. Front. Plant Sci. 2022, 13, 915197. [Google Scholar] [CrossRef]
- Liang, R.L.; Wang, L.J.; Wang, X.Q.; Zhang, J.T.; Gan, X. Effects of exogenous ALA on leaf photosynthesis, photosynthate transport, and sugar accumulation in Prunus persica L. Forests 2023, 14, 723. [Google Scholar] [CrossRef]
- Liu, H.; Kang, L.; Wang, L.J. Promotion of 5-aminolevulinic acid on seed germination of watermelon (Citrullus lanatus) under salt stress. J. Fruit Sci. 2006, 1, 854–859. [Google Scholar]
- Wang, L.J.; Jiang, W.B.; Huang, B.J. Promotion of photosynthesis by 5-aminolevulinic acid (ALA) during and after chilling stress in melon seedlings grown under low light condition. Acta Hortic. Sin. 2004, 3, 321–326. [Google Scholar]
- Yin, L.L.; Yu, X.C.; Wang, Y.H.; Xu, Z.H.; Li, K.; Han, D.J. Effect of 5-aminolevulinic acid on chilling tolerance in cucumber seedlings. Acta Agric. Boreali-Occident. Sin. 2007, 16, 166–169. [Google Scholar]
- Liu, T.; Xua, J.J.; Li, J.M.; Hu, X.H. NO is involved in JA- and H2O2-mediated ALA-induced oxidative stress tolerance at low temperatures in tomato. Environ. Exp. Bot. 2019, 161, 334–343. [Google Scholar] [CrossRef]
- Ma, N.; Qi, L.; Gao, J.J.; Chao, K.C.; Hu, Q.F.; Jiang, H.G.; Wang, L.G. Effects of 5-aminolevulinic acid on cutting growth under high temperature condition and leaf chlorophyll fast fluorescence characteristics of Ficus carica L. J. Nanjing Agric. Univ. 2015, 38, 546–553. [Google Scholar]
- An, Y.Y.; Zhang, L.Y.; Feng, X.X.; Tian, F.; Li, J.; Wang, L.J. Effects of 5-aminolevulinic acid on low light tolerance of apple leaves. Acta Bot. Boreal-Occident. Sin. 2016, 36, 987–995. [Google Scholar]
- Long, S.; Liu, B.; Gong, J.; Wang, R.; Gao, S.; Zhu, T.; Guo, H.; Liu, T.; Xu, Y. 5-Aminolevulinic acid promotes low-light tolerance by regulating chloroplast ultrastructure, photosynthesis, and antioxidant capacity in tall fescue. Plant Physiol. Biochem. 2022, 190, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, T.; Gao, W.R.; Guo, S.R.; Li, D.C.; Sun, Y.J. Effects of 5-aminolevulinic (ALA) on growth and photosynthesis of pepper under low temperature stress. Jiangsu J. Agric. Sci. 2011, 27, 612–616. [Google Scholar]
- Wu, Y.; Jin, X.; Liao, W.B.; Hu, L.L.; Dawuda, M.M.; Zhao, X.J.; Tang, Z.Q.; Gong, T.Y.; Yu, J.H. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.P.; Wang, L.J. Effects of 5-aminolevulinic (ALA) on chlorophyll fluorescence dynamics of watermelon seedlings under shade condition. Acta Hortic. Sin. 2007, 34, 901–908. [Google Scholar]
- Zhao, Y.Y.; Yan, F.; Hu, L.P.; Zhou, X.T.; Zou, Z.R. Effects of 5-aminolevulinic acid on photosynthetic characteristics of tomato seedlings under NaCl stress. Chin. J. Appl. Ecol. 2014, 25, 2919–2926. [Google Scholar]
- Wang, Y.X.; Chen, X.Y.; Li, X.Y.; Song, Y.H.; Wang, J.A.; Huang, J.; Xue, R.L. Exogenous application of 5-aminolevulinic acid alleviated damage to wheat chloroplast ultrastructure under drought stress by transcriptionally regulating genes correlated with photosynthesis and chlorophyll biosynthesis. Acta Physiol. Plant. 2022, 44, 12. [Google Scholar] [CrossRef]
- Chen, Z.; An, Y.; Wang, L. ALA reverses ABA-induced stomatal closure by modulating PP2AC and SnRK2.6 activity in apple leaves. Hortic. Res. 2023, 10, 37287446. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, J.T.; Wang, L.J. ALA induces stomatal opening through regulation among PTPA, PP2AC, and SnRK2.6. Front. Plant Sci. 2023, 14, 1206728. [Google Scholar] [CrossRef]
- Shen, M.; Duan, C.H.; Zhang, Y.P.; Cheng, Y.; Wang, L.J.; Li, B.J. Effects of exogenous ALA treatment on thinning and fruit quality in ‘Hosui’ pear (Pyrus pyrifolia). Acta Hortic. Sin. 2011, 38, 1515–1522. [Google Scholar]
- Shen, M.; Chen, X.H.; Xie, L.; Li, C.; Sun, X.E.; Zhang, Z.P.; Wang, L.J. Effects of amino-acid fertilizer on photosynthesis in leaves of pear cultivars. J. Nanjing Agric. Univ. 2012, 35, 81–86. [Google Scholar]
- Shen, M.; Zhang, Z.P.; Wang, L.J. Effect of 5-aminolevulinic acid (ALA) on leaf diurnal photosynthetic characteristics and antioxidant activity in pear (Pyrus pyrifolia Nakai). In Artificial Photosynthesis; Najafpour, M.M., Ed.; InTech: London, UK, 2011; pp. 239–265. [Google Scholar]
- Wang, P.F.; Zhao, J.X.; Li, T.; Yu, L.C.; Li, Y.F.; Jia, Y.L.; Zhang, Y.X.; Zhang, H.X.; Quan, C.; Xu, J.F.; et al. Study on the effect of exogenous ALA to alleviate low temperature injury in pear flower organs. China Fruits 2023, 1, 16–21. [Google Scholar]
- Xiao, C.Z.; Zhang, S.L.; Hu, H.J.; Tian, R.; Wu, J.; Yang, Z.J.; Ma, C.Y. Effects of bagging and exogenous 5-aminolevulinic acid treatment on coloration of ‘Yunhongli 2’. J. Nanjing Agric. Univ. 2012, 35, 25–29. [Google Scholar]
- Li, W.X.; Cao, X.Y.; Wang, X.Q. Effect of ALA combined with calcium chloride and sucrose on fruit quality and color of ‘Nanhong’ pear. China Fruits 2022, 1, 35–38. [Google Scholar]
- T/JAASS 88-2023; 5-Aminolevulic Acid Technical Procedures for Application in Peach. Jiangsu Association of Agricultural Science Societies: Nanjing, China, 2023.
- Wang, X.K. Experimental Principles and Techniques of Plant Physiology and Biochemistry; Higher Education Press: Beijing, China, 2006; pp. 130–134. [Google Scholar]
- Harel, E.; Klein, S. Light dependent formation of δ-aminolevulinic acid in etiolated of higher plants. Biochem. Biophys. Res. Commun. 1972, 49, 364–370. [Google Scholar] [CrossRef]
- Li, P.M.; Gao, H.Y. Application of rapid chlorophyll fluorescence induction kinetics analysis in the study of photosynthesis. J. Plant Physiol. Mol. Biol. 2005, 1, 559–566. [Google Scholar]
- Wang, D.Y.; Li, X.; Xu, L.; An, Y.Y.; Wang, L.J. 5-Aminolevulinic acid (ALA) improved the heat resistance of leaves of two garden plants. J. Bot. Res. 2018, 73, 350–365. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Y.J.; Jiang, B.; Zhang, Y.; Li, Y.B.; He, L.; Wang, Y.C. Transient expression in tobacco by Agrobacterium mediated transformmation. J. Northeast. For. Univ. 2010, 38, 110–112. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Hotta, Y.; Tanaka, H.; Takaoka, Y.; Takeuchi, Y.; Konnai, M. Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant Growth Regul. 1997, 22, 109–114. [Google Scholar] [CrossRef]
- Liu, W.Q.; Kang, L.; Wang, L.J. Effects on strawberry photosynthesis and relations to anti-oxidant enzymes of ALA. Acta Bot. Boreali-Occident. Sin. 2006, 1, 57–62. [Google Scholar]
- Wang, J.N.; Li, X.Y.; Wei, S.M.; Zhao, H.J.; Zhao, M.Q.; Wang, Y.X. Regulation of exogenous 5-aminolevulinic acid on photosynthesis and D1 protein of wheat seedlings under drought stress. Crop J. 2018, 1, 121–126. [Google Scholar]
- Wang, L.J.; Zhang, J.T.; Zhong, Y.; Zhang, L.Z.; Yang, H.; Liu, L.B.; Zhou, J.Y.; Iqbal, M.M.; Gan, X. Regulation of 5-aminolevunilic acid and its application in agroforestry. Forests 2023, 14, 1857. [Google Scholar] [CrossRef]
- Chen, L.H.; Liu, L.B.; An, Y.Y.; Zhang, Z.P.; Wang, L.J. Preliminary studies on the possible mechanism underlying 5-aminolevulinic acid-induced stomatal opening in apple leaves. Acta Hortic. Sin. 2014, 41, 1965–1974. [Google Scholar]
- Liu, L.B.; An, Y.Y.; Xiong, L.J.; Wang, L.J. Flavonols induced by 5-aminolevulinic acid are involved in regulation of stomatal opening in apple leaves. Acta Hortic. Sin. 2016, 43, 817–828. [Google Scholar] [CrossRef]
- Xiong, L.J.; An, Y.Y.; Wang, L.J. The role of microtubule skeleton and PP1/PP2A protein phosphatase in ALA-ABA regulating stomatal movement in apple leaves. Acta Hortic. Sin. 2018, 45, 2073–2088. [Google Scholar]
- Yang, Y.X.; Xia, J.X.; Fang, X.; Jia, H.R.; Wang, X.C.; Lin, Y.L.; Liu, S.Y.; Ge, M.Q.; Pu, Y.F.; Fang, J.G.; et al. Drought stress in ‘Shine Muscat’ grapevine: Consequences and a novel mitigation strategy-5-aminolevulinic acid. Front. Plant Sci. 2023, 14, 1129114. [Google Scholar] [CrossRef]
- Fromme, P.; Melkozernov, A.; Jordan, P.; Krauss, N. Structure and function of photosystem I: Interaction with its soluble electron carriers and external antenna systems. FEBS Lett. 2003, 555, 40–44. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tanaka, A.; Tsuji, H. Effects of 5-aminolevulinic acid on the accumulation of chlorophyll-b and apoproteins of the light-harvesting chlorophyll a/b-protein complex of photosystem II. Plant Cell Physiol. 1993, 34, 465–472. [Google Scholar]
- Wang, L.J.; Jiang, W.B.; Zhang, Z.; Yao, Q.H.; Matsui, H.; Ohara, H. Biosynthesis and physiological activity of 5-aminolevulinic acid and its potential application in agriculture. Plant Physiol. Commun. 2003, 3, 185–192. [Google Scholar]
- Zhong, Y.; Liu, C.; Wei, B.; Zhang, J.; An, Y.; Wang, L. Exogenous 5-aminolevulinic acid promotes osmotic stress tolerance of walnuts by modulating photosynthesis, osmotic adjustment, and antioxidant systems. Forests 2023, 14, 789. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Li, J.; Dawuda, M.M.; Ali, B.; Wu, Y.; Yu, J.; Tang, Z.; Lyu, J.; Xiao, X.; et al. Exogenous application of 5-aminolevulinic acid promotes coloration and improves the quality of tomato fruit by regulating carotenoid metabolism. Front. Plant Sci. 2021, 12, 683868. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, J.W.; Li, R.R.; Tian, Q.; Wu, Y.; You, J.H. The mitigation effect of exogenous ALA on salt stress of tomato seedlings. China Cucurbits Veg. 2023, 36, 56–64. [Google Scholar]
- Wang, H.; Liu, Z.; Luo, S.; Li, J.; Zhang, J.; Li, L.; Xie, J. 5-Aminolevulinic acid and hydrogen sulphide alleviate chilling stress in pepper (Capsicum annuum L.) seedlings by enhancing chlorophyll synthesis pathway. Plant Physiol. Biochem. 2021, 167, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.R.; Zhang, F.T.; Gao, J.X.; Sun, X.Q.; Deng, X.J. An overview of chlorophyll biosynthesis in higher plants. Acta Bot. Boreali-Occident. Sin. 2009, 29, 629–636. [Google Scholar]
- Wang, R.J.; Jiang, Y.; Zhu, F.; Tang, Z.; Liu, X.S.; Jiang, Y.; Wang, Y. Application of 820 nm optical absorption curve technology to study the performance of leaf optical system I under stress conditions. Guangxi For. Sci. 2020, 49, 152–155. [Google Scholar]
- Sun, Y.P.; Liu, J.; Cao, X.; Huang, Y.J.; Hall, A.M.; Guo, C.B.; Wang, L.J. Effects of 5-aminolevulinic acid treatment on photosynthesis of strawberry. Photosynthetica 2017, 55, 276–284. [Google Scholar] [CrossRef]
- Song, Q.N.; Feng, P.W.; Liu, Y.; Yang, X.H. The research progress of the mechanism on PSII assemble and repair circulation. Plant Physiol. J. 2019, 55, 133–140. [Google Scholar]
- Hou, P.F.; Ma, J.Q.; Zhao, P.F.; Zhang, H.L.; Zhao, H.J.; Liu, H.S.; Zhao, Y.D.; Wang, Y.X. Effects of betaine on chloroplast protective enzymes and psbA gene expression in wheat seedlings under drought stress. Acta Agron. Sin. 2013, 39, 1319–1324. [Google Scholar] [CrossRef]
- Wang, M.; Shan, J.X.; Zhong, Z.P.; Li, L.B.; Kang, T.Y. Advances in the research of structure and function of photosystem ii core antenna complexes CP43 and CP47. Chin. Bull. Bot. 2000, 17, 141–149. [Google Scholar]
- Jia, H.S.; Li, D.Q.; Ham, Y.Q. Advances in studies on photoinhibition in photosynthesis of higher plants. Chin. Bull. Bot. 2000, 17, 218–224. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Gene ID) | Primer | Amplicon Size |
|---|---|---|
| Actin (LOC103931062) | Fw:cccagaagtgctcttccaac Rv:ttgatcttcatgctgcttgg | 1550 |
| PsbA (ON478189.1:308-1369) | Fw:cattttctgtggtttccctgat Rv:cgcccttggattgctgtt | 1061 |
| PsbB (ON478189.1:75738-77264) | Fw:tttcgggcttgtgcttttt Rv:cccacgctggatttacgg | 1526 |
| PsbC (ON478189.1:36669-38088) | Fw:ggcgggggagatgtaagaa Rv:ccaaacaaagcaacaagcaata | 1419 |
| PsbD (ON478189.1:35660-36721) | Fw:atgggagttgctggggtg Rv:ccaaaaacggttagcggtg | 1061 |
| PsbE (ON478189.1:67489-67740) | Fw:gaattccttgtcggctttctg Rv:ccttatttattgcgggttggt | 251 |
| PsbF (ON478189.1:c67479-67360) | Fw:gtacctaccgtttcttttttaggg Rv:ttggatgaactgcattgctga | 119 |
| PsbH (ON478189.1:77867-78091) | Fw:gatctggccgcccaaaa Rv:tgcgagacccatcaaagga | 224 |
| PsbJ (ON478189.1:c67094-66972) | Fw:acgacacaatcaaacccgaa Rv:caaaaatgagtaataacccccag | 122 |
| PsbL (ON478189.1:c67337-67221) | Fw:acgacacaatcaaacccgaa Rv:caaaaatgagtaataacccccag | 116 |
| PsbP (LOC103965639) | Fw:ctgagtccgagggtggctt Rv:tccttcatctccgtctgctgt | 807 |
| PsbQ (LOC103933887) | Fw:cttgctgtctgaaacaaccctc Rv:ttctctccacgactcgaactct | 674 |
| PsbW (LOC103946003) | Fw:tggaaggaaaaaaggagagca Rv:cgagggaggaagtatagataaagtaga | 623 |
| PsbY (LOC103949857) | Fw:caaccgcatgacaaacaagag Rv:ccaccacaaacagcaacaact | 887 |
| Psb27 (LOC103947836) | Fw:ccacccccacaatccca Rv:ccttatttatcacctcctccgtt | 874 |
| Psb28 (LOC103959875) | Fw:tcaggcttgttatgttgggg Rv:tctggtattgtctgttcatcggt | 987 |
| HEMA (LOC103957838) | Fw:acaaggaggataggctaaggaag Rv:ccattggaccatgaaggagtt | 2115 |
| GSA (LOC103936176) | Fw:taatggtatccgcaagctcac Rv:aacttcctggctccctcaac | 1716 |
| HEMB (LOC125472965) | Fw:gttgaggcaaatgaagacgagt Rv:tgcagctataggcagaggagag | 1537 |
| HEMD (LOC103957047) | Fw:tgctatgacaggagcggtga Rv:cctaagaatgggattgaaaaatg | 1367 |
| HEMF (LOC125474519) | Fw:caccgccccgatacctt Rv:tgacaccagccttctccca | 1485 |
| HEMG (LOC103967516) | Fw:atgatggtggactgtggtttga Rv:gacctggaggaggcgga | 2010 |
| CHLH (LOC103946503) | Fw:ggaggcaagaggggcg Rv:gggcaaaaccagtaagcga | 4452 |
| CHLM (LOC103931332) | Fw:ccaagaccgttgagaatgtga Rv:gccctgccgagaagcc | 1288 |
| CRD1 (LOC103938880) | Fw:agacaaatcgcactacagcca Rv:ttcaagtttttcaccacggg | 1424 |
| DVR (LOC103958557) | Fw:cgtggcaaaaacccaaaag Rv:aacacccaaatcagtcacatcc | 1531 |
| POR (LOC103949278) | Fw:ccctccgactgcctctca Rv:tactggcttccacttctgcttat | 1843 |
| CHLG (LOC103965133) | Fw:ttgaaggagatagagcaatggg Rv:tggaaaaagacttggggagc | 1470 |
| CAO (LOC103940306) | Fw:cccttctcccagcacttctac Rv:gacacatccaggtttcccatc | 2157 |
| NtActin (LOC107830629) | Fw:cctgaggtccttttccaacca Rv:ggattccggcagcttccatt | 1574 |
| GUS (LOC103930738) | Fw:cctgaggtccttttccaacca Rv:tcattgtttgcctccctgct | 2038 |
| Promoter of PsbA | Fw:accatgattacgccaagcttgatatccaccatcttgacttggtgatgg Rv:accacccggggatcctctagaatggagagagagagatattggagttgaagg | 254 |
| Promoter of PsbB | Fw:accatgattacgccaagcttctaattgctacacgcattgcagaatg Rv:accacccggggatcctctagatgccaatcagcttctctccacc | 502 |
| Promoter of PsbJ | Fw:accatgattacgccaagcttcgtaccaataacgtccaaatgattggtaca Rv:accacccggggatcctctagaaccaggactttagacaacggattacgtag | 1032 |
| Promoter of HEMA | Fw:accatgattacgccaagcttggacttttttttttttgggagaggagct Rv:accacccggggatcctctagaagggagggatatggaattgggataagg | 2000 |
| Promoter of GSA | Fw:accatgattacgccaagcttccccctgtagtattacaaattaaaaatttt Rv:accacccggggatcctctagaagactttggtcttagtatatatgagttctt | 2000 |
| Promoter of HEMF | Fw:accatgattacgccaagcttgtactatatctcactttgatgtgaaagcg Rv:accacccggggatcctctagatggaaatacccctttatcaatgaaaaatag | 2000 |
| Promoter of CHLH | Fw:accatgattacgccaagcttgatctgcggatttatcaacatcaagctc Rv:accacccggggatcctctagaataatcggatgaactgagttgtagacatga | 2000 |
| Weeks after Foliar Spraying of ALA | Treatment | Wk | Mo | Sm | φo | φPo | φEo | φDo | RC/CS (×103) | PIABS | PItotal | VPSI (×10−4) | VPSII-PSI (×10−5) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Control | 0.37 ± 0.01 ab | 0.53 ± 0.01 a | 28.21 ± 0.74 b | 0.63 ± 0.01 f | 0.80 ± 0.01 d | 0.54 ± 0.00 f | 0.16 ± 0.00 a | 4.48 ± 0.06 bc | 5.77 ± 0.15 c | 4.37 ± 0.23 c | 20.27 ± 0.19 a | 3.52 ± 0.10 abc |
| ALA | 0.35 ± 0.00 d | 0.49 ± 0.01 bc | 30.71 ± 0.69 a | 0.65 ± 0.01 de | 0.85 ± 0.00 b | 0.56 ± 0.01 de | 0.15 ± 0.00 bc | 4.73 ± 0.03 a | 6.18 ± 0.13 abc | 5.04 ± 0.17 ab | 19.90 ± 0.31 ab | 3.86 ± 0.16 a | |
| 2 | Control | 0.37 ± 0.00 b | 0.50 ± 0.01 b | 27.25 ± 0.64 b | 0.66 ± 0.01 cd | 0.80 ± 0.01 d | 0.56 ± 0.00 cd | 0.15 ± 0.00 bc | 4.35 ± 0.03 c | 5.90 ± 0.22 bc | 4.53 ± 0.13 c | 19.13 ± 0.33 bc | 3.11 ± 0.70 d |
| ALA | 0.35 ± 0.00 cd | 0.46 ± 0.01 cd | 30.81 ± 0.71 a | 0.69 ± 0.01 a | 0.85 ± 0.00 ab | 0.59 ± 0.00 a | 0.14 ± 0.00 d | 4.54 ± 0.05 b | 6.61 ± 0.21 a | 5.22 ± 0.13 a | 19.85 ± 0.26 ab | 3.57 ± 0.14 ab | |
| 3 | Control | 0.35 ± 0.01 d | 0.49 ± 0.01 bc | 28.23 ± 0.64 b | 0.67 ± 0.01 bc | 0.81 ± 0.01 d | 0.58 ± 0.00 bc | 0.14 ± 0.00 d | 4.49 ± 0.02 bc | 5.75 ± 0.17 c | 4.67 ± 0.12 bc | 18.17 ± 0.37 de | 3.18 ± 0.19 bcd |
| ALA | 0.33 ± 0.00 e | 0.45 ± 0.01 d | 30.26 ± 0.42 a | 0.69 ± 0.01 a | 0.86 ± 0.00 a | 0.59 ± 0.00 a | 0.14 ± 0.00 e | 4.70 ± 0.07 a | 6.49 ± 0.18 a | 5.25 ± 0.15 a | 18.34 ± 0.27 cde | 3.45 ± 0.12 bcd | |
| 4 | Control | 0.39 ± 0.00 a | 0.50 ± 0.01 b | 23.70 ± 0.50 c | 0.66 ± 0.01 cd | 0.83 ± 0.00 c | 0.56 ± 0.01 d | 0.16 ± 0.00 b | 4.11 ± 0.05 d | 5.95 ± 0.18 bc | 4.97 ± 0.13 ab | 17.56 ± 0.43 e | 2.04 ± 0.10 e |
| ALA | 0.37 ± 0.00 b | 0.48 ± 0.01 bcd | 24.69 ± 0.60 c | 0.68 ± 0.01 ab | 0.85 ± 0.00 ab | 0.58 ± 0.00 ab | 0.15 ± 0.00 c | 4.35 ± 0.07 c | 6.34 ± 0.18 ab | 5.41 ± 0.09 a | 18.35 ± 0.22 cde | 2.33 ± 0.09 e | |
| 5 | Control | 0.36 ± 0.01 bc | 0.50 ± 0.01 b | 26.70 ± 0.68 b | 0.64 ± 0.01 ef | 0.82 ± 0.01 c | 0.55 ± 0.01 ef | 0.15 ± 0.00 bc | 4.51 ± 0.07 b | 5.75 ± 0.16 c | 4.75 ± 0.08 bc | 18.29 ± 0.24 cde | 3.04 ± 0.15 d |
| ALA | 0.35 ± 0.01 cd | 0.48 ± 0.01 bc | 27.86 ± 0.62 b | 0.66 ± 0.01 cd | 0.85 ± 0.01 ab | 0.56 ± 0.01 bcd | 0.14 ± 0.00 d | 4.69 ± 0.05 a | 6.09 ± 0.17 abc | 5.04 ± 0.13 ab | 18.58 ± 0.25 cd | 3.13 ± 0.13 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, J.; Wang, L. Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves. Forests 2024, 15, 769. https://doi.org/10.3390/f15050769
Wang X, Zhang J, Wang L. Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves. Forests. 2024; 15(5):769. https://doi.org/10.3390/f15050769
Chicago/Turabian StyleWang, Xinqing, Jianting Zhang, and Liangju Wang. 2024. "Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves" Forests 15, no. 5: 769. https://doi.org/10.3390/f15050769
APA StyleWang, X., Zhang, J., & Wang, L. (2024). Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves. Forests, 15(5), 769. https://doi.org/10.3390/f15050769