The Importance of Maintaining Upland Forest Habitat Surrounding Salamander Breeding Ponds: Case Study of the Eastern Tiger Salamander in New York, USA

Abstract
:1. Introduction
2. Methods
2.1. Study Site


2.2. Radiotelemetry Equipment
2.3. Trapping and Surgical Procedures
2.4. Tracking Methods
2.5. Data Analysis
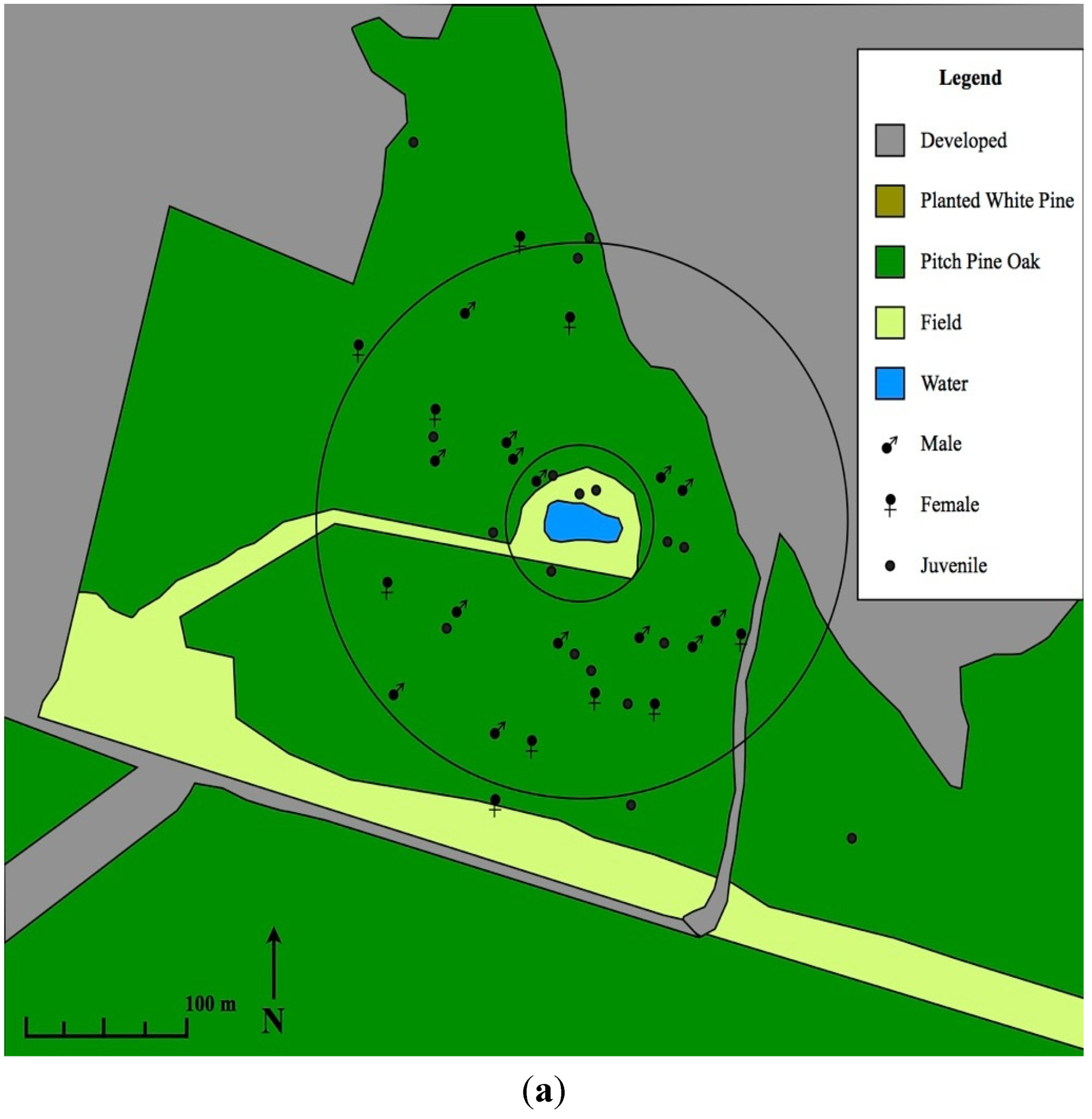
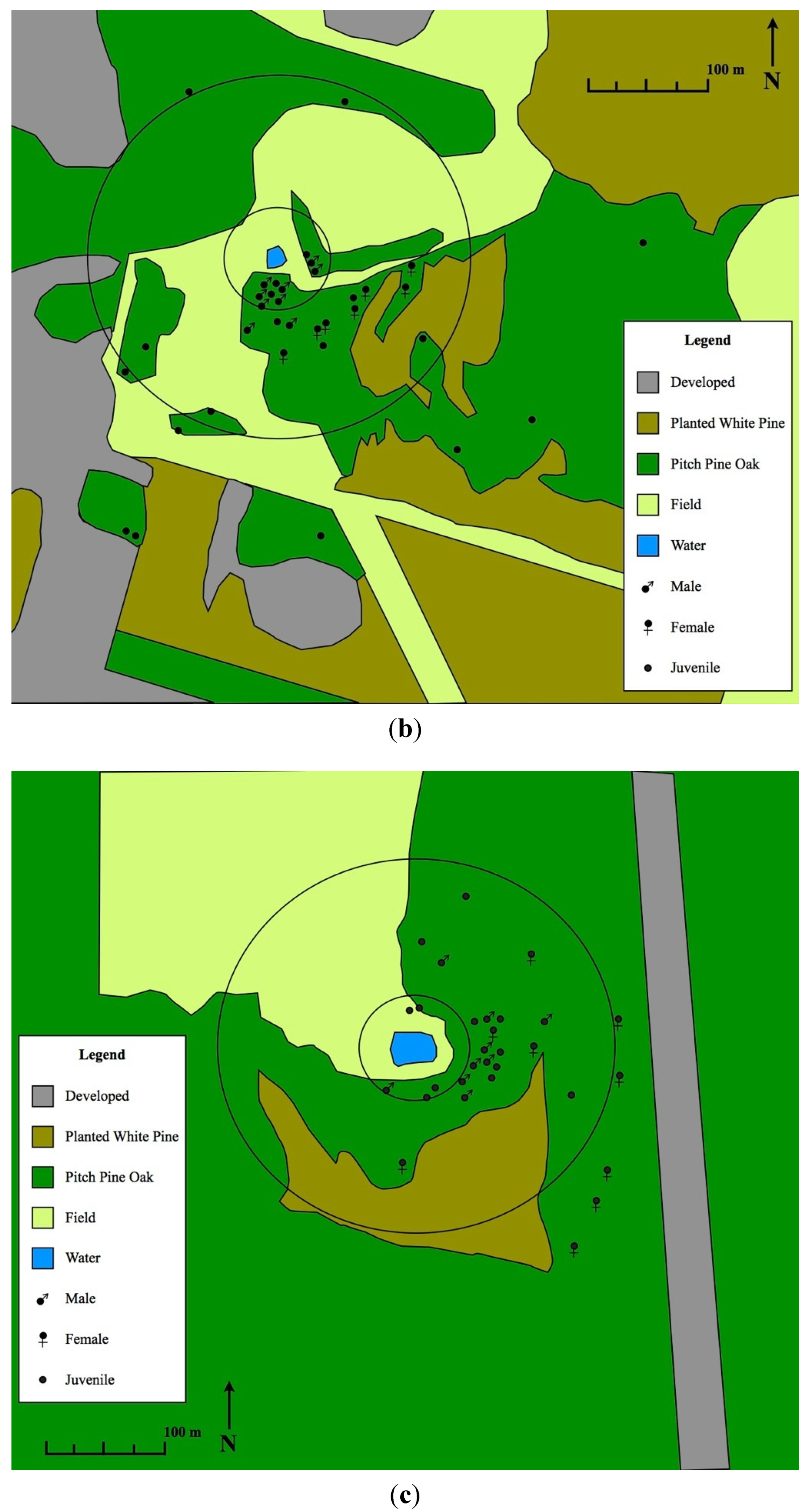
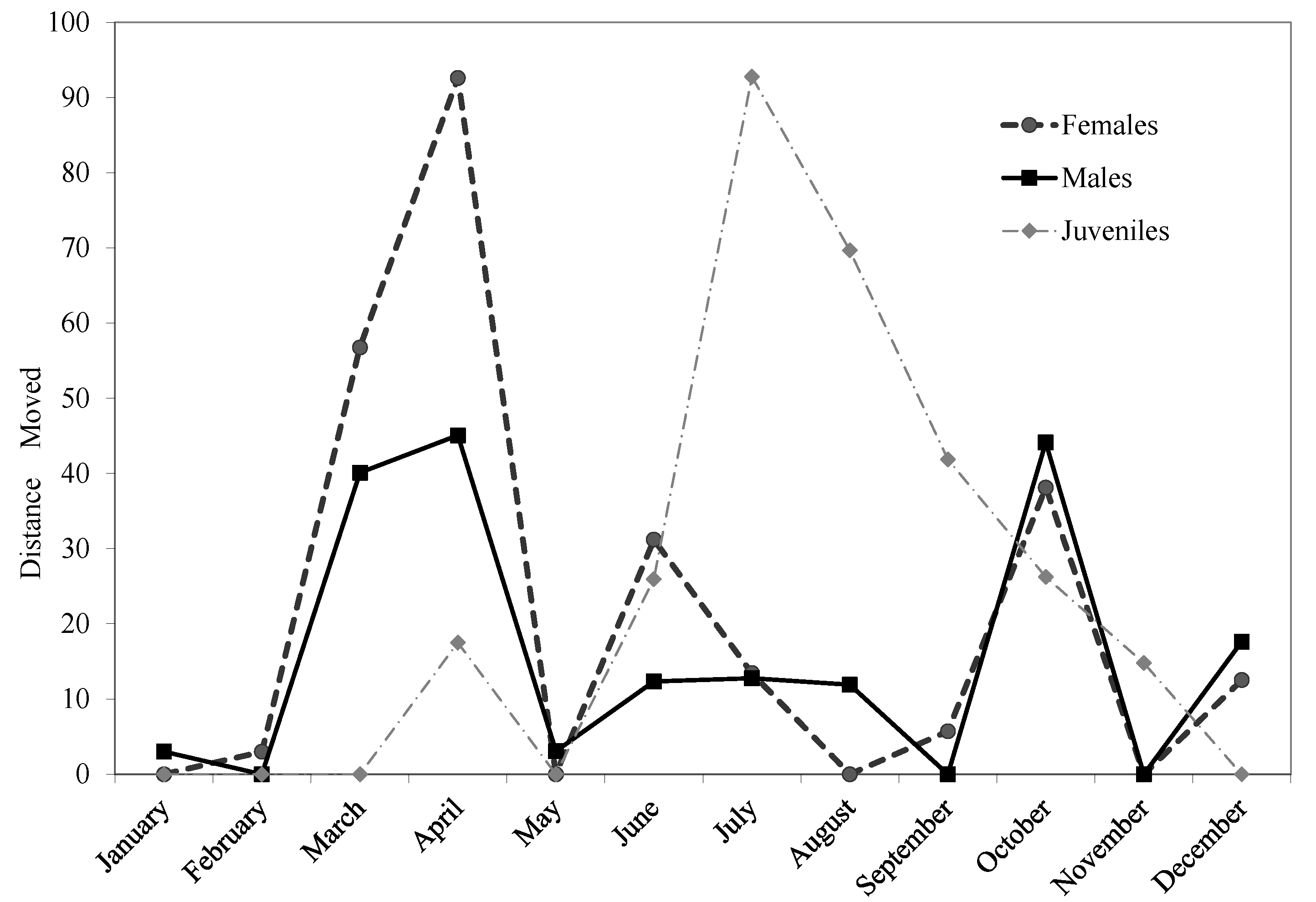
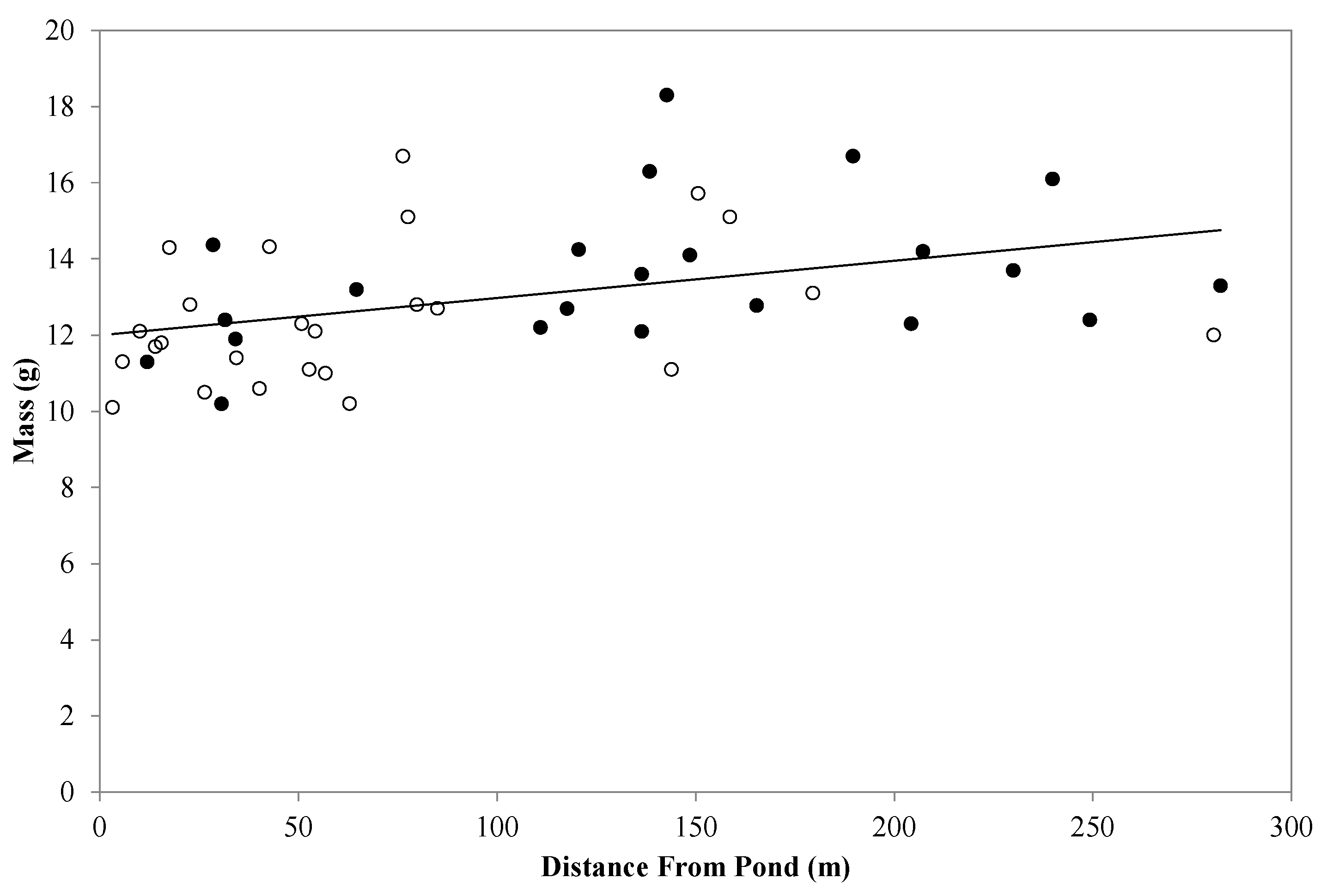
3. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Days Tracked and Distance From Pond | Mean ± SD | Range | |||||
|---|---|---|---|---|---|---|---|
| Pond | Males | Females | Juveniles | Males | Females | Juveniles | |
| L1 | n = 15 | n = 10 | n = 16 | ||||
| # Days Tracked | 90.5 ± 88.4 | 147.4 ± 186.6 | 34.1 ± 28.8 | 26–394 | 28–402 | 5–196 | |
| Distance from Pond (m) | 65.4 ± 42.3 | 106.9 ± 28.0 | 85.9 ± 75.0 | 9.1–153.4 | 68.4–145.6 | 3.2–239.8 | |
| L3 | n = 9 | n = 7 | n = 19 | ||||
| # Days Tracked | 35.6 ± 28.5 | 44.0 ± 35.7 | 59.9 ± 60.0 | 18–97 | 18–84 | 12–304 | |
| Distance from Pond (m) | 28.9 ± 13.9 | 84.0 ± 16.4 | 141.5 ± 86.0 | 12.6–59.3 | 72.2–108.5 | 5.7–282.1 | |
| L7 | n = 9 | n = 9 | n = 12 | ||||
| # Days Tracked | 70.1 ± 79.4 | 116.2 ± 98.8 | 39.1 ± 29.8 | 3–371 | 6–401 | 10–96 | |
| Distance from Pond (m) | 49.7 ± 22.9 | 148.4 ± 72.0 | 58.3 ± 42.0 | 20.1–99.8 | 31.1–249.3 | 17.5–142.7 | |
| Combined | n = 33 | n = 26 | n = 47 | ||||
| # Days Tracked | 68.6 ± 75.7 | 108.8 ± 133.2 | 45.8 ± 45.1 | 3–394 | 6–402 | 5–304 | |
| Distance from Pond (m) | 51.9 ± 34.9 | 115.1 ± 52.0 | 101.3 ± 79.7 | 9.1–153.4 | 31.1–249.3 | 3.2–282.1 | |

| Movement Distance Comparisons Between Sex and Age Classes | Pond | ||
|---|---|---|---|
| L1 | L3 | L7 | |
| Males vs. Females | Z = 2.946 * | Z = 3.334 * | Z = 2.782 * |
| P = 0.013 | P = 0.001 | P = 0.005 | |
| Males vs. Juveniles | Z = 0.376 | Z = 3.025 ** | Z = 0.071 |
| P = 0.707 | P = 0.002 | P = 0.943 | |
| Females vs. Juveniles | Z = 1.265 | Z = 1.937 ** | Z = 2.772 * |
| P = 0.206 | P = 0.052 | P = 0.006 | |

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Knutson, M.G.; Sauer, J.R.; Olsen, D.A.; Mossman, M.J.; Hemesath, L.M.; Lanoo, M.J. Effects of landscape composition and wetland fragmentation on frog and toad abundance and species richness in Iowa and Wisconsin, U.S.A. Conserv. Biol. 1999, 13, 1437–1446. [Google Scholar] [CrossRef]
- Weyrauch, S.L.; Grubb, T.C. Patch and landscape characteristics associated with the distribution of woodland amphibians in an agricultural fragmented landscape: An information-theoretic approach. Biol. Conserv. 2004, 111, 443–450. [Google Scholar] [CrossRef]
- Rittenhouse, T.A.G.; Semlitsch, R.D. Grasslands as movement barriers for a forest-associated salamander: Migration behavior of adult and juvenile salamanders as a distinct habitat edge. Biol. Conserv. 2006, 131, 14–22. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Biological delineation of terrestrial buffer zones for pond-breeding salamanders. Conserv. Biol. 1998, 12, 1113–1119. [Google Scholar] [CrossRef]
- Gibbs, J.P. Wetland loss and biodiversity conservation. Conserv. Biol. 2000, 14, 314–317. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Critical elements for biologically based recovery plans of aquatic-breeding amphibians. Conserv. Biol. 2002, 16, 619–629. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Bodie, J.R. Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conserv. Biol. 2003, 17, 1219–1228. [Google Scholar] [CrossRef]
- Harper, E.B.; Rittenhouse, T.A.G.; Semlitsch, R.D. Demographic consequences of terrestrial habitat loss for pool-breeding amphibians: Predicting extinction risks associated with inadequate size of buffer zones. Conserv. Biol. 2008, 22, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Burke, V.J.; Gibbons, J.W. Terrestrial buffer zones and wetland conservation: A case study of freshwater turtles in a Carolina bay. Conserv. Biol. 1995, 9, 1365–1369. [Google Scholar] [CrossRef]
- Gibbons, J.W. Terrestrial habitat: A vital component for herpetofauna of isolated wetlands. Wetlands 2003, 23, 630–635. [Google Scholar] [CrossRef]
- Roth, E.D. Buffer zone applications in snake ecology: A case study using cottonmouths (Agkistrodon piscivorus). Copeia 2005, 2005, 399–402. [Google Scholar] [CrossRef]
- Veysey, J.S.; Babbitt, K.J.; Cooper, A. An experimental assessment of buffer width: Implications for salamander migratory behavior. Biol. Conserv. 2009, 142, 2227–2239. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Differentiating migration and dispersal processes for pond-breeding amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Madison, D.; Farrand, L. Habitat use during breeding and emigration in radio-implanted tiger salamanders, Ambystoma tigrinum. Copeia 1998, 2, 402–410. [Google Scholar] [CrossRef]
- Anderson, J.D.; Hassinger, D.D.; Dalrymple, G.H. Natural mortality of eggs and larvae of Ambystoma t tigrinum. Ecology 1971, 52, 1107–1112. [Google Scholar] [CrossRef]
- Madison, D.M. The emigration of radio-implanted spotted salamanders, Ambystoma maculatum. J. Herpetol. 1997, 31, 542–551. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Biek, R.; Funk, W.C.; Maxell, B.A.; Mills, L.S. What is missing in amphibian decline research: Insights from ecological sensitivity analysis. Conserv. Biol. 2002, 16, 728–734. [Google Scholar] [CrossRef]
- Vonesh, J.R.; de la Cruz, O. Complex life cycles and density dependence: Assessing the contribution of egg mortality to amphibian declines. Oecologia 2002, 133, 325–333. [Google Scholar] [CrossRef]
- Trenham, P.C.; Shaffer, H.B. Amphibian upland habitat use and its consequences for population viability. Ecol. Appl. 2005, 15, 1158–1168. [Google Scholar] [CrossRef]
- Orloff, S.G. Movement pattern and migration distances in an upland population of California tiger salamander (Ambystoma californiense). Herpetol. Conserv. 2011, 6, 266–276. [Google Scholar]
- Calhoun, A.J.K.; Klemens, M.W. Best Development Practices: Conserving Pool-Breeding Amphibians in Residential Developments in the Northeastern United States; MCA Technical paper No. 5; Metroplitan Conservation Alliance, Wildlife Conservation Society: Bronx, NY, USA, 2002; p. 57. [Google Scholar]
- Richter, S.C.; Young, J.E.; Siegel, R.A.; Johnson, G.N. Postbreeding movements of the dark gopher frog, Rana sevosa Goin and Netting: Implications for conservation and management. J. Herpetol. 2001, 35, 316–321. [Google Scholar] [CrossRef]
- Trenham, P.C. Terrestrial habitat use by adult California tiger salamanders. J. Herpetol. 2001, 35, 343–346. [Google Scholar] [CrossRef]
- Faccio, S.D. Postbreeding emigration and habitat use by Jefferson and spotted salamanders in Vermont. J. Herpetol. 2003, 37, 479–489. [Google Scholar] [CrossRef]
- Porej, D.; Micacchion, M.; Hetherington, T.E. Core terrestrial habitat for conservation of local populations of salamanders and woo frogs in agricultural landscapes. Biol. Conserv. 2004, 120, 399–409. [Google Scholar]
- Searcy, C.A.; Shaffer, H.B. Calculating biologically accurate mitigation credits: Insights from the California tiger salamander. Conserv. Biol. 2008, 22, 997–1005. [Google Scholar] [PubMed]
- Castelle, A.J.; Johnson, A.W.; Conolly, C. Wetland and stream buffer size requirements—A review. J. Environ. Qual. 1994, 23, 878–882. [Google Scholar] [CrossRef]
- Pittman, S.E.; Osbourn, M.S.; Semlitsch, R.D. Movement ecology of amphibians: A missing component for understanding population declines. Biol. Conserv. 2014, 169, 44–53. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Peterson, C.R.; Ritson, P.I. Seasonal migration of Columbia spotted frogs (Rana lutiventris) among complementary resources in a high mountain basin. Can. J. Zool. 2002, 80, 1849–1862. [Google Scholar] [CrossRef]
- Regosin, J.V.; Windmiller, B.S.; Reed, J.M. Terrestrial habitat use and winter densities of the wood frog (Rana sylvatica). J. Herpetol. 2003, 37, 390–394. [Google Scholar] [CrossRef]
- Bartelt, P.E.; Peterson, C.R.; Klaver, R.W. Sexual Differences in the Post-Breeding Movements and Habitats Selected by Western Toads (Bufo boreas) in Southeastern Idaho. Herpetologica 2004, 60, 455–467. [Google Scholar] [CrossRef]
- Schabetsberger, R.; Jehle, R.; Maletzky, A.; Pesta, J.; Sztatecsny, M. Delineation of terrestrial reserves for amphibians: Post-breeding migrations of Italian crested newts (Triturus c carnifex) at high altitude. Biol. Conserv. 2004, 117, 95–104. [Google Scholar] [CrossRef]
- Steen, D.A.; Smith, L.L.; Miller, G.J.; Sterrett, S.C. Post-breeding terrestrial movements of Ambystoma tigrinum (eastern tiger salamander). Southeast. Nat. 2006, 5, 285–288. [Google Scholar] [CrossRef]
- Fellers, G.M.; Kleeman, P.M. California red-legged frog (Rana draytonii) movement and habitat use: Implications for conservation. J. Herpetol. 2007, 41, 276–286. [Google Scholar] [CrossRef]
- McDonough, C.; Paton, P.W.C. Salamander dispersal across a forested landscape fragmented by a golf course. J. Wildl. Manag. 2007, 71, 1163–1169. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Terrestrial movements of an eastern tiger salamander, Ambystoma tigrinum. Herpetol. Rev. 1983, 14, 112–113. [Google Scholar]
- Madison, D.M. Habitat-contingent reproductive behavior in radio-implanted salamanders: A model and test. Anim. Behav. 1998, 55, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Loredo, I.; van Vuren, D.; Morrison, M.L. Habitat use and migration behavior of the California tiger salamander. J. Herpetol. 1996, 30, 282–285. [Google Scholar] [CrossRef]
- Vasconcelos, D.; Calhoun, J.K. Movement patterns of adult and juvenile Rana sylvatica (LeConte) and Ambystoma maculatum (Shaw) in three restored seasonal pools in Maine. J. Herpetol. 2004, 38, 551–561. [Google Scholar] [CrossRef]
- Connete, G.M.; Semlitsch, R.D. Successful use of a passive integrated transponder (PIT) system for below-ground detection of plethodontid salamanders. Wildl. Res. 2012, 39, 1–6. [Google Scholar] [CrossRef]
- Pittman, S.E.; Semlitsch, R.D. Habitat type and distance to edge effect movement behavior of juvenile pond-breeding salamanders. J. Zool. 2013, 291, 154–162. [Google Scholar]
- Ousterhout, B.; Ousterhout, R.; Semlitsch, R.D. Measuring terrestrial movement behavior using passive integrated transponder (PIT) tags: Effects of tag size on detection, movement, survival, and growth. Behav. Ecol. Sociobiol. 2014, 68, 343–350. [Google Scholar] [CrossRef]
- Gahl, M.; Bates College, Lewiston, ME, USA. Personal Communication, 2011.
- Titus, V.R.; Green Mountain College, Poultney, VT, USA. Unpublished data. 2013.
- Semlitsch, R.D.; Wilbur, H.M. Effects of pond drying time on metamorphosis and survival in the salamander Ambystoma talpoideum. Copeia 1988, 1988, 978–983. [Google Scholar] [CrossRef]
- Madison, D.; Andrus, R. Tiger Salamander Habitat Use; Final Progress Report to NYS DEC. 76; Albany, NY, USA, 1993; pp. 1–137. [Google Scholar]
- New York State Department of Environmental Conservation. Article 24 Freshwater Wetlands Title 23 of Article 71 of the Environmental Conservation Law; New York State Department of Environmental Conservation: Albany, NY, USA, 1997.
- Rosenblatt, D.; New York State Department of Environmental Conservation, Albany, NY, USA. Personal Communication, 2012.
- Gamble, L.R.; McGarigal, K.; Jenkins, C.L.; Timm, B.C. Limitations of regulated “buffer zones” for the conservation of marbled salamanders. Wetlands 2006, 26, 298–306. [Google Scholar] [CrossRef]
- Gamble, L.R.; McGarigal, K.; Compton, B.W. Fidelity and dispersal in the pond-breeding amphibian, Ambystoma opacum: Implications for spatio-temporal population dynamics and conservation. Biol. Conserv. 2007, 139, 127–257. [Google Scholar]
- Peterman, W.E.; Locke, J.L.; Semlitsch, R.D. Spatial and temporal patterns of water loss in heterogeneous landscapes: Using plaster models as amphibian analogues. Can. J. Zool. 2013, 91, 135–140. [Google Scholar] [CrossRef]
- Osbourn, M.S.; Connette, G.M.; Semlitsch, R.D. Effects of fine-scale forest habitat quality on movement and settling decisions in juvenile pond-breeding salamanders. Ecol. Appl. 2014, 24, 1719–1729. [Google Scholar] [CrossRef]
- Madison, D.M.; Titus, V.R.; Lamoureux, V.S. Movement patterns and radiotelemetry. In Amphibian Conservation and Ecology: A Handbook of Techniques; Dodd, C.K., Ed.; Oxford University Press: Oxford, UK, 2010; pp. 185–202. [Google Scholar]
- Titus, V.R. Movements, connectivity, and management: Conserving the New York State endangered eastern tiger salamander. Ph.D. Thesis, Binghamton University, Binghamton, NY, USA, 2013. [Google Scholar]
- Cecala, K.K.; Price, S.J.; Dorcas, M.E. A comparison of the effectiveness of recommended doeses of MS-222 (tricain methanesulfonate) and Oragel (benzocaine) for amphibian anesthesia. Herpetol. Rev. 2007, 38, 63–66. [Google Scholar]
- Peterman, W.E.; Semlitsch, R.D. Effects of tricaine methanesulfonate (MS-222) concentrations on anesthetization and recovery in four plethodontid salamanders. Herpetol. Rev. 2006, 37, 303–304. [Google Scholar]
- Mineau, P.; Madison, D. Radio-tracking of Peromyscus. leucopus. Can. J. Zool. 1977, 55, 465–468. [Google Scholar] [CrossRef]
- Beyer, H.L. Hawth’s Analysis Tools for ArcGIS 2004. Available online: http://www.spatialecology.com/htools (accessed on 16 July 2008).
- Trivers, R. Parental investment and sexual selection. In Sexual Selection and the Descent of Man 1871–1971; Campbell, B., Ed.; Aldine Press: Chicago, IL, USA, 1972; pp. 139–179. [Google Scholar]
- Zimmer, K.D.; Hanson, M.A.; Butler, M.G. Factors influencing invertebrate communities in prairie wetlands: A multivariate approach. Can. J. Fish Aquat. Sci. 2000, 57, 76–85. [Google Scholar] [CrossRef]
- Neckles, H.A.; Murkin, H.R.; Cooper, J.A. Influences of seasonal flooding on macroinvertebrate abundance in wetland habitats. Freshw. Biol. 1990, 23, 311–322. [Google Scholar] [CrossRef]
- Chew, R.W. The water exchange of some small mammals. Ecol. Monogr. 1951, 21, 215–225. [Google Scholar] [CrossRef]
- Getz, L.L. Factors influencing the local distribution of shrews. Am. Midl. Nat. 1961, 65, 67–88. [Google Scholar] [CrossRef]
- Trenham, P.C.; Shaffer, H.B.; Koenig, W.D.; Stromberg, M.R. Life history and demographic variation in the California tiger salamander (Ambystoma californiense). Copeia 2000, 2000, 365–377. [Google Scholar] [CrossRef]
- Church, D.R.; Bailey, L.L.; Wilbur, H.M.; Kendall, W.L.; Hines, J.E. Iteroparity in the variable environment of the salamander Ambystoma tigrinum. Ecology 2007, 88, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Pittman, S.E.; Osbourn, M.J.; Drake, D.L.; Semlitsch, R.D. Predation of juvenile ringed salamanders (Ambystoma annulatum) during initial movement out of ponds. Herpetol. Conserv. Biol. 2013, 8, 681–687. [Google Scholar]
- Whiteman, H.H.; Wissinger, S.A.; Brown, W.S. Growth and foraging consequences of facultative paedomorphosis in the tiger salamander, Ambystoma tigrinum nebulosum. Evol. Ecol. 1996, 10, 429–442. [Google Scholar] [CrossRef]
- Phillips, C.A.; Johnson, J.R.; Drezlik, M.J.; Petzing, J.E. Effects of hydroperiod on recruitment of mole salamanders (Genus Ambystoma) at a temporary pond in Vermillion County, Illinois. Trans. Ill. Acad. Sci. 2002, 95, 131–139. [Google Scholar]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Titus, V.R.; Bell, R.C.; Becker, C.G.; Zamudio, K.R. Connectivity and gene flow among eastern tiger salamander (Ambystoma tigrinum tigrinum) populations in highly modified anthropogenic landscapes. Conserv. Genet. 2014, 15, 1447–1462. [Google Scholar] [CrossRef]
- Compton, B.W.; McGarigal, K.; Cushman, S.A.; Gamble, L.R. A resistant-kernel model of connectivity for amphibians that breed in vernal pools. Conserv. Biol. 2007, 21, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, K.R.; Gibbs, H.L.; Waite, H.A. Efficacy of land-cover models in predicting isolation of marbles salamander populations in a fragmented landscape. Conserv. Biol. 2009, 25, 1232–1241. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titus, V.; Madison, D.; Green, T. The Importance of Maintaining Upland Forest Habitat Surrounding Salamander Breeding Ponds: Case Study of the Eastern Tiger Salamander in New York, USA. Forests 2014, 5, 3070-3086. https://doi.org/10.3390/f5123070
Titus V, Madison D, Green T. The Importance of Maintaining Upland Forest Habitat Surrounding Salamander Breeding Ponds: Case Study of the Eastern Tiger Salamander in New York, USA. Forests. 2014; 5(12):3070-3086. https://doi.org/10.3390/f5123070
Chicago/Turabian StyleTitus, Valorie, Dale Madison, and Timothy Green. 2014. "The Importance of Maintaining Upland Forest Habitat Surrounding Salamander Breeding Ponds: Case Study of the Eastern Tiger Salamander in New York, USA" Forests 5, no. 12: 3070-3086. https://doi.org/10.3390/f5123070
APA StyleTitus, V., Madison, D., & Green, T. (2014). The Importance of Maintaining Upland Forest Habitat Surrounding Salamander Breeding Ponds: Case Study of the Eastern Tiger Salamander in New York, USA. Forests, 5(12), 3070-3086. https://doi.org/10.3390/f5123070