3.1. Influence of Genotype and Plantation Age on Poplar Allometry
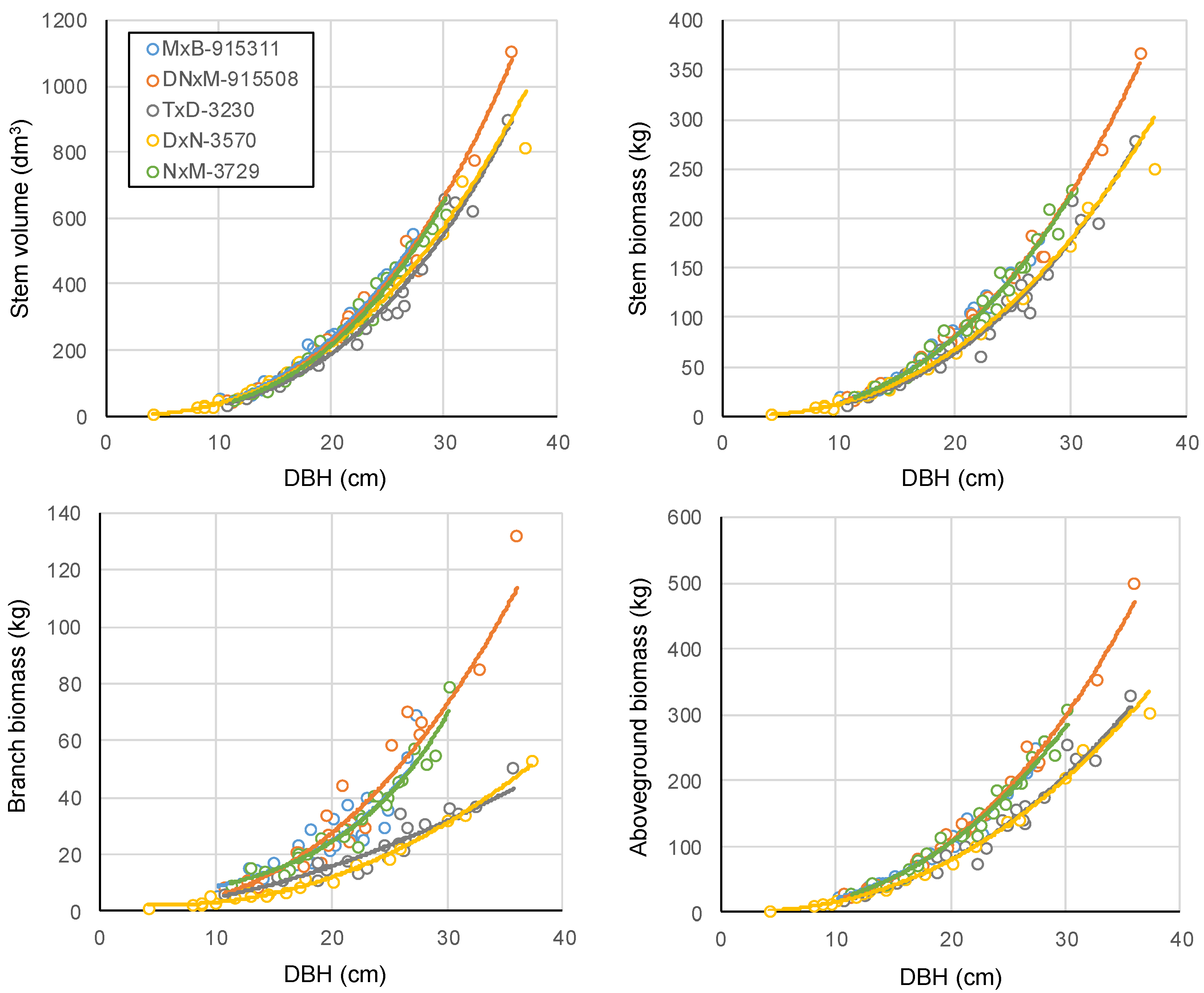
The allometric relationships developed between DBH and volume or biomass of the different tree compartments for 13 year-old hybrid poplars suggested an important influence of the genotype on allometry (
Figure 1,
Table 2). In addition, results from the analyses of covariance suggest a significant Genotype effect on the regression between DBH and volume or biomass of different tree compartments (
Table 4). For most tree compartments studied, allometry was found to be very similar for
P.
maximoviczii hybrids (
Figure 1). No significant Genotype effect was observed on stem, branch and aboveground biomass, when the three
P.
maximoviczii hybrids (clones MxB-915311, DNxM-915508 and NxM-3729) were used as different treatment levels in the analysis of covariance (
Table 4). Likewise, allometry was also found to be relatively similar for clones DxN-3570 and TxD-3230, which are both Euramerican hybrids (
Figure 1). No significant Genotype effect was observed on stem and aboveground woody biomass, when these two clones were used as different treatment levels in the analysis of covariance (
Table 4). More generally, for all tree compartments studied, highly significant Genotype effects (
p < 0.001) were also observed when the group formed by
P.
maximoviczii hybrids and the group formed by Euramerican hybrids were the treatment levels used in the analysis of covariance (
Table 4). These trends may be related to the fact that for a given tree DBH, Euramerican hybrids tended to have lower stem volume, stem biomass, branch biomass and aboveground biomass than
P.
maximoviczii hybrids (
Figure 1). This evidence suggests that accurate estimates of wood or biomass production and carbon storage in tree biomass for clones of different parentages will require the use of genotype-specific, or perhaps hybrid-specific allometric relationships. Similar conclusions have been drawn regarding biomass estimates in short-rotation coppices of willows [
35]. The implications for yield results of using clone-specific
vs. general allometric relationships will be further discussed in
Section 3.3.
Figure 1.
Clone-specific allometric relationships between diameter at breast height (cm) and stem volume (dm3), stem dry biomass (kg), branch dry biomass (kg), and woody aboveground dry biomass (kg). The blue line (clone MxB-915311) is sometimes overlapped by the green (clone NxM-3729) and orange (clone DNxM-915508) lines.
Figure 1.
Clone-specific allometric relationships between diameter at breast height (cm) and stem volume (dm3), stem dry biomass (kg), branch dry biomass (kg), and woody aboveground dry biomass (kg). The blue line (clone MxB-915311) is sometimes overlapped by the green (clone NxM-3729) and orange (clone DNxM-915508) lines.
Our results also confirm previous observations done in poplar agroforestry systems of the studied region (southern Québec, Canada). In 9 year-old poplar buffer strips, allometric relationships between DBH and branch biomass were similar for clones MxB-915311 and DNxM-915508, while clone DxN-3570 had much lower branch biomass at a given DBH [
34]. Yet, 7 year-old poplar clones of different parentages (D, DxB and DxN) showed little variation in allometry of woody biomass compartments [
50]. Thus, additional studies involving destructive sampling are needed to evaluate the allometry of clones that have different genetic assemblages. Eventually, general allometric relationships for different groups of clones or hybrids could be developed because some clones and group of clones showed a very similar allometry (
Figure 1,
Table 4).
Table 4.
Significance level (p-value) of the main effects for the analysis of covariance (ANCOVA) testing DBH, as a continuous covariate, and genotype or plantation age, as nominal main effects on the different growth variables (stem volume, stem biomass, branch biomass and aboveground woody biomass). The covariate was always significant at p < 0.001. The Genotype effect was tested on 13 year-old poplar data, while the Plantation age effect was tested on the data set combining data from 8 and 13 year-old poplars. Data were log transformed prior to analysis; NS = non significant.
Table 4.
Significance level (p-value) of the main effects for the analysis of covariance (ANCOVA) testing DBH, as a continuous covariate, and genotype or plantation age, as nominal main effects on the different growth variables (stem volume, stem biomass, branch biomass and aboveground woody biomass). The covariate was always significant at p < 0.001. The Genotype effect was tested on 13 year-old poplar data, while the Plantation age effect was tested on the data set combining data from 8 and 13 year-old poplars. Data were log transformed prior to analysis; NS = non significant.
| Main Effects and Treatment Levels | Stem Volume (dm3/tree) | Stem Biomass (kg/tree) | Branch Biomass (kg/tree) | Aboveground Biomass (kg/tree) |
|---|
| Genotype | | | | |
| MxB, NxM, DNxM, TxD and DxN | <0.001 | <0.001 | <0.001 | <0.001 |
| MxB, NxM and DNxM | <0.05 | NS | NS | NS |
| TxD and DxN | <0.01 | NS | <0.001 | NS |
| Euramerican and P. maximoviczii hybrids | <0.001 | <0.001 | <0.001 | <0.001 |
| Plantation age | |
| 8 and 13 years | <0.01 | <0.05 | <0.05 | |
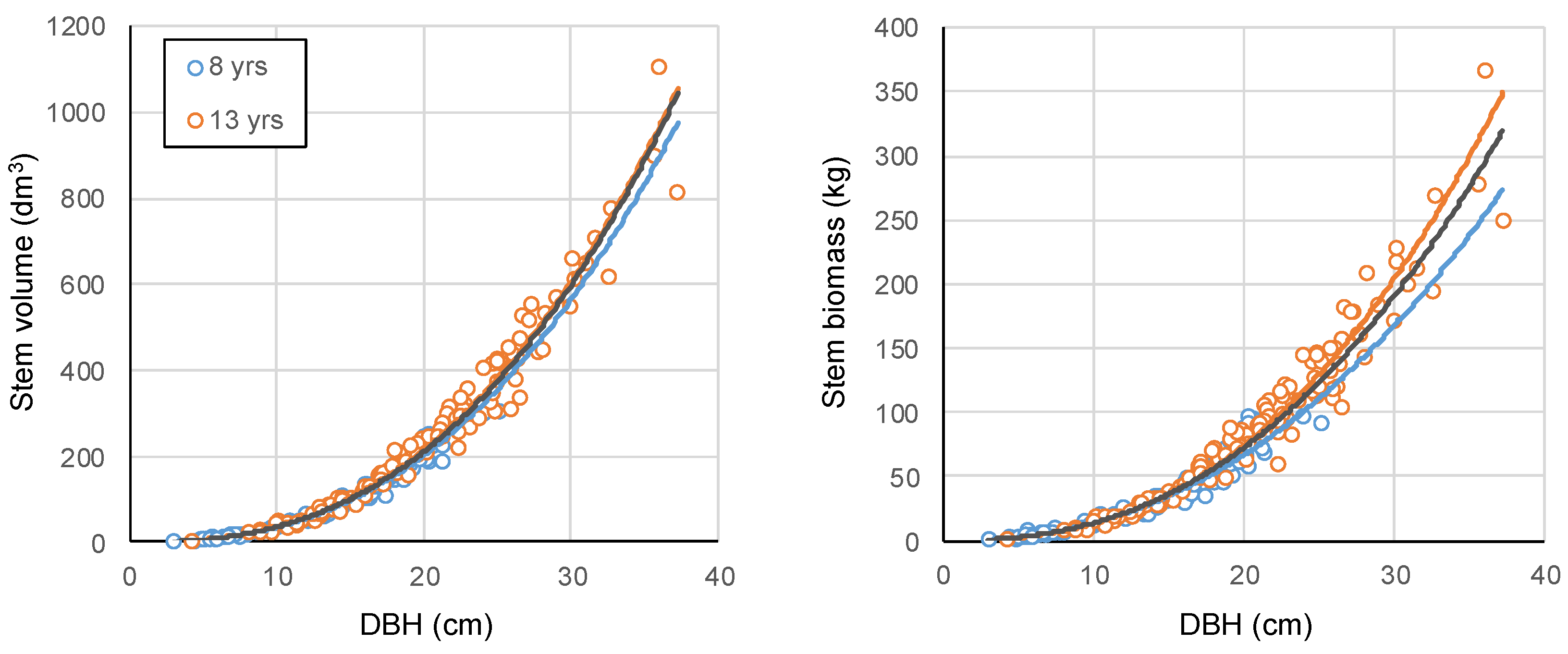
General models developed with data from the 5 clones (
n = 120) for stem volume and biomass suggest a significant Plantation age effect on poplar allometry, as revealed by the analysis of covariance (
Figure 2,
Table 3 and
Table 4). Although the size of the age effect seems more marginal for stem volume estimation, it was likely important for stem biomass, especially at larger DBH value. For example, a hybrid poplar with 30 cm of DBH would have an estimated stem volume of 563 dm
3 using the model developed with 8 year-old trees and of 605 dm
3 using the model developed with 13 year-old trees (7% difference between the two estimates). However, for the same tree, stem biomass estimates would be 167 and 203 kg, if model developed with 8 and 13 year-old trees were used respectively (18% difference between the two estimates). This trend may be the consequence of the influence of tree age on stem wood density. As observed by DeBell
et al. [
51], wood density of three clones with a
P.
trichocarpa parentage showed a continuous increase from year 5 to year 9. Therefore, using a biomass equation developed with juvenile poplars might result in an underestimation of the biomass of more mature poplars, and
vice versa, because of these possible wood density changes over the years.
Figure 2.
General (5 clones combined) allometric relationships between diameter at breast height (cm) and stem volume (dm3), and stem dry biomass (kg) in poplar plantations. The black line indicates the general model developed combining the 8 and 13 year data obtained from trees harvested in the poplar plantations.
Figure 2.
General (5 clones combined) allometric relationships between diameter at breast height (cm) and stem volume (dm3), and stem dry biomass (kg) in poplar plantations. The black line indicates the general model developed combining the 8 and 13 year data obtained from trees harvested in the poplar plantations.
3.2. Hybrid Poplar Yields in Mature Plantations
After 13 years, strong Site × Clone interactions were observed this study, suggesting that the 5 poplar genotypes had a different growth response to the plantation environment across the studied ecological gradient (
Table 1,
Table 5 and
Table 6). At the site level, very high yields can be achieved in poplar plantations in southern Québec, but moderate to high site fertility is imperative for achieving high productivity (
Table 6). In the same experimental design, Truax
et al. [
11] showed that soil P availability and elevation were strongly correlated with 8 year-old poplar productivity. Five years later, sites having the highest soil P availability and lower elevation (Bedford, Brompton and Ste-Catherine) were still the most productive. High yields were also observed at high elevation, at the fertile site of La Patrie (440 m), especially for
P.
maximoviczii hybrids (
Table 6), a trend that also emerged in our previous study.
Located at 80 m of elevation in the St. Lawrence Valley and having the highest soil P availability, the Bedford site was the most productive. The Bedford site was also recently abandoned and is the only site that is surrounded by intensive row crop agriculture, which is an indication of the high soil quality at this location. Depending on the clone used at this site, mean DBH reached 23.3–30.4 cm, while mean tree volume and mean annual yield reached 360–637 dm
3/tree and 20.5–31.4 m
3/ha/year or 9.0–14.3 t/ha/year (
Table 5 and
Table 6). In the province of Québec (Canada), these are the highest yields ever reported in the literature for poplar plantations with a relatively low stem density at planting (833 tree/ha) [
52]. Only narrow riparian buffers bordering fertile agricultural sites and short-rotation coppice systems, at much higher tree densities, were found to be more productive than the Bedford site in southern Québec [
34,
53].
Table 5.
Site × Clone interaction for mean tree DBH, stem volume per tree, stem biomass per tree, branch biomass per tree, and aboveground woody biomass per tree in 13 year-old poplar plantations.
Table 5.
Site × Clone interaction for mean tree DBH, stem volume per tree, stem biomass per tree, branch biomass per tree, and aboveground woody biomass per tree in 13 year-old poplar plantations.
| Sites | Clones | DBH (cm) | Stem Volume (dm3/tree) | Stem Biomass (kg/tree) | Branch Biomass (kg/tree) | Woody Biomass (kg/tree) |
|---|
| Bedford | MxB-915311 | 23.3 | 360 | 122 | 36.3 | 158 |
| DNxM-915508 | 27.8 | 553 | 189 | 61.9 | 251 |
| TxD-3230 | 28.9 | 531 | 170 | 30.2 | 200 |
| DxN-3570 | 30.4 | 637 | 197 | 33.9 | 231 |
| NxM-3729 | 28.0 | 549 | 190 | 58.4 | 248 |
| Brompton | MxB-915311 | 22.8 | 347 | 117 | 35.5 | 153 |
| DNxM-915508 | 23.8 | 374 | 130 | 43.3 | 174 |
| TxD-3230 | 27.7 | 467 | 150 | 27.9 | 178 |
| DxN-3570 | 23.9 | 342 | 108 | 18.9 | 126 |
| NxM-3729 | 26.0 | 451 | 158 | 47.3 | 205 |
| Ste-Catherine | MxB-915311 | 18.6 | 204 | 70 | 22.1 | 92 |
| DNxM-915508 | 21.1 | 272 | 96 | 32.4 | 129 |
| TxD-3230 | 22.2 | 280 | 92 | 19.5 | 111 |
| DxN-3570 | 16.2 | 156 | 50 | 9.1 | 59 |
| NxM-3729 | 21.8 | 290 | 105 | 31.7 | 136 |
| La Patrie | MxB-915311 | 22.0 | 312 | 106 | 31.8 | 138 |
| DNxM-915508 | 20.6 | 251 | 89 | 30.3 | 120 |
| TxD-3230 | 26.7 | 443 | 143 | 26.6 | 169 |
| DxN-3570 | 13.7 | 97 | 31 | 6.1 | 37 |
| NxM-3729 | 24.4 | 380 | 135 | 40.0 | 175 |
| Melbourne | MxB-915311 | 21.9 | 321 | 109 | 32.9 | 142 |
| DNxM-915508 | 21.5 | 281 | 99 | 33.5 | 133 |
| TxD-3230 | 23.4 | 299 | 98 | 20.9 | 119 |
| DxN-3570 | 15.8 | 130 | 42 | 7.7 | 50 |
| NxM-3729 | 21.4 | 263 | 96 | 28.7 | 125 |
| Fitch Bay | MxB-915311 | 18.7 | 212 | 73 | 23.1 | 96 |
| DNxM-915508 | 17.9 | 229 | 81 | 27.1 | 108 |
| TxD-3230 | 25.0 | 365 | 119 | 23.6 | 142 |
| DxN-3570 | 9.0 | 29 | 10 | 2.7 | 13 |
| NxM-3729 | 20.9 | 263 | 95 | 29.1 | 124 |
| Ham | MxB-915311 | 15.4 | 129 | 45 | 15.9 | 61 |
| DNxM-915508 | 15.4 | 120 | 44 | 15.4 | 59 |
| TxD-3230 | 17.4 | 146 | 50 | 12.7 | 62 |
| DxN-3570 | 9.4 | 39 | 13 | 3.3 | 16 |
| NxM-3729 | 15.5 | 113 | 44 | 15.6 | 59 |
| Stornoway | MxB-915311 | 13.3 | 83 | 30 | 12.2 | 42 |
| DNxM-915508 | 10.8 | 51 | 20 | 7.1 | 27 |
| TxD-3230 | 11.6 | 50 | 18 | 6.3 | 24 |
| DxN-3570 | 5.7 | 14 | 5 | 2.7 | 7 |
| NxM-3729 | 14.1 | 92 | 36 | 13.8 | 50 |
| SE | | 1.3 | 40 | 13 | 3.4 | 16 |
| p | | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Table 6.
Site × Clone interaction for survival and mean annual yield in 13 year-old poplar plantations. The branch biomass ratio is the percentage of woody aboveground biomass that is branch biomass. Woody biomass yield after 8 years is also indicated along with the yield variation between 8 and 13 year-old plantations. Key environmental variables (elevation and soil available P) are indicated for each site (see Truax
et al. [
11]).
Table 6.
Site × Clone interaction for survival and mean annual yield in 13 year-old poplar plantations. The branch biomass ratio is the percentage of woody aboveground biomass that is branch biomass. Woody biomass yield after 8 years is also indicated along with the yield variation between 8 and 13 year-old plantations. Key environmental variables (elevation and soil available P) are indicated for each site (see Truax et al. [11]).
| Sites and Key Environmental Variables | Clones | Survival (%) | Stem Volume (m3/ha/yr) | Stem Biomass (t/ha/yr) | Branch Biomass (t/ha/yr) | Branch Biomass Ratio (%) | Aboveground Woody Biomass (t/ha/yr) | Variation 8 vs. 13 Years (t/ha/yr) |
|---|
| | | 13 Years | 8 Years | 13 Years | |
|---|
| Bedford | MxB-915311 | 89 | 20.3 | 6.9 | 2.1 | 23 | 8.0 | 9.0 | +1.0 |
| 80 m | DNxM-915508 | 89 | 31.4 | 10.8 | 3.5 | 25 | 12.3 | 14.3 | +2.0 |
| 77 kg P/ha | TxD-3230 | 83 | 28.4 | 9.1 | 1.6 | 15 | 8.7 | 10.7 | +2.0 |
| | DxN-3570 | 81 | 30.9 | 9.6 | 1.7 | 15 | 7.6 | 11.2 | +3.6 |
| | NxM-3729 | 75 | 26.1 | 9.0 | 2.8 | 23 | 12.6 | 11.8 | −0.8 |
| Brompton | MxB-915311 | 92 | 20.2 | 6.9 | 2.1 | 23 | 6.8 | 9.0 | +2.1 |
| 170 m | DNxM-915508 | 83 | 19.6 | 6.8 | 2.3 | 25 | 6.3 | 9.1 | +2.8 |
| 60 kg P/ha | TxD-3230 | 92 | 27.4 | 8.8 | 1.6 | 16 | 7.0 | 10.5 | +3.5 |
| | DxN-3570 | 89 | 19.5 | 6.1 | 1.1 | 15 | 5.9 | 7.2 | +1.3 |
| | NxM-3729 | 92 | 26.5 | 9.3 | 2.8 | 23 | 9.6 | 12.1 | +2.4 |
| Ste-Catherine | MxB-915311 | 92 | 11.9 | 4.1 | 1.3 | 24 | 3.3 | 5.4 | +2.1 |
| 230 m | DNxM-915508 | 89 | 15.5 | 5.5 | 1.8 | 25 | 4.7 | 7.3 | +2.7 |
| 50 kg P/ha | TxD-3230 | 89 | 16.1 | 5.3 | 1.1 | 17 | 4.3 | 6.4 | +2.1 |
| | DxN-3570 | 89 | 9.0 | 2.9 | 0.5 | 15 | 2.8 | 3.4 | +0.6 |
| | NxM-3729 | 92 | 17.1 | 6.1 | 1.9 | 23 | 6.0 | 8.0 | +2.0 |
| La Patrie | MxB-915311 | 83 | 16.5 | 5.7 | 1.7 | 23 | 5.7 | 7.3 | +1.6 |
| 440 m | DNxM-915508 | 92 | 14.8 | 5.3 | 1.8 | 25 | 5.0 | 7.0 | +2.0 |
| 58 kg P/ha | TxD-3230 | 42 | 11.0 | 3.6 | 0.7 | 16 | 3.4 | 4.2 | +0.9 |
| | DxN-3570 | 33 | 2.1 | 0.7 | 0.1 | 16 | 0.8 | 0.8 | 0 |
| | NxM-3729 | 92 | 22.3 | 7.9 | 2.4 | 23 | 8.2 | 10.3 | +2.1 |
| Melbourne | MxB-915311 | 89 | 18.3 | 6.3 | 1.9 | 23 | 4.1 | 8.2 | +4.1 |
| 330 m | DNxM-915508 | 75 | 13.8 | 4.9 | 1.6 | 25 | 3.1 | 6.5 | +3.4 |
| 17 kg P/ha | TxD-3230 | 31 | 6.0 | 2.0 | 0.4 | 17 | 1.4 | 2.4 | +1.0 |
| | DxN-3570 | 50 | 4.4 | 1.4 | 0.3 | 16 | 1.1 | 1.7 | +0.6 |
| | NxM-3729 | 92 | 15.5 | 5.6 | 1.7 | 23 | 5.3 | 7.3 | +2.0 |
| Fitch Bay | MxB-915311 | 72 | 9.7 | 3.4 | 1.1 | 24 | 2.0 | 4.4 | +2.4 |
| 260 m | DNxM-915508 | 53 | 9.8 | 3.4 | 1.1 | 25 | 2.0 | 4.6 | +2.6 |
| 12 kg P/ha | TxD-3230 | 25 | 5.8 | 1.9 | 0.4 | 17 | 1.2 | 2.3 | +1.0 |
| | DxN-3570 | 8 | 0.2 | 0.1 | 0.0 | 22 | 0.1 | 0.1 | 0 |
| | NxM-3729 | 69 | 11.3 | 4.1 | 1.3 | 23 | 3.1 | 5.4 | +2.3 |
| Ham | MxB-915311 | 92 | 7.5 | 2.6 | 0.9 | 26 | 2.1 | 3.6 | +1.5 |
| 320 m | DNxM-915508 | 89 | 6.9 | 2.5 | 0.9 | 26 | 2.0 | 3.4 | +1.5 |
| 26 kg P/ha | TxD-3230 | 81 | 7.6 | 2.6 | 0.7 | 20 | 2.5 | 3.2 | +0.7 |
| | DxN-3570 | 64 | 1.6 | 0.5 | 0.1 | 20 | 0.6 | 0.7 | +0.1 |
| | NxM-3729 | 92 | 6.6 | 2.6 | 0.9 | 26 | 2.7 | 3.5 | +0.8 |
| Stornoway | MxB-915311 | 92 | 4.9 | 1.7 | 0.7 | 29 | 1.1 | 2.5 | +1.3 |
| 450 m | DNxM-915508 | 78 | 2.6 | 1.0 | 0.4 | 27 | 0.8 | 1.3 | +0.5 |
| 9 kg P/ha | TxD-3230 | 39 | 1.1 | 0.4 | 0.1 | 27 | 0.3 | 0.5 | +0.2 |
| | DxN-3570 | 17 | 0.3 | 0.1 | 0.0 | 27 | 0.1 | 0.1 | 0 |
| | NxM-3729 | 86 | 5.1 | 2.0 | 0.8 | 28 | 1.6 | 2.8 | +1.2 |
| SE | | 7 | 1.9 | 0.6 | 0.2 | - | 0.6 | 0.8 | - |
| p | | <0.001 | <0.001 | <0.001 | <0.001 | - | <0.001 | <0.001 | - |
High yields were also observed at the Brompton site, which had similar soil chemical characteristics (
Table 1), but slightly higher elevation (170 m) than the Bedford site. At this site, mean DBH reached 22.8–27.7 cm, while mean tree volume and mean annual yield reached 342–467 dm
3/tree and 19.5–27.4 m
3/ha/year or 7.2–12.1 t/ha/year, depending on the clone (
Table 5 and
Table 6). The yields observed at the Bedford and Brompton sites after 13 years were also comparable to the most productive plantations of Sweden and Denmark [
24,
26], which were grown under similar climatic conditions and silvicultural regime.
The two high soil fertility sites (Bedford and Brompton) also experienced the greatest increases in volume and biomass growth from year 8 to year 13. During this 5 year period, a net gain of 178 and 170 m
3/ha, or 69.5 and 67.3 t/ha, was observed at the Bedford and Brompton sites respectively (
Table 7). Consequently, the productivity gap between the high yielding and the low yielding sites (Fitch Bay, Ham and Stornoway) has widened from year 8 to year 13. For example, after 8 years, Bedford and Stornoway had cumulated 179 and 8 m
3/ha respectively, for a productivity gap of 171 m
3/ha between these two sites. After 13 years, the same two sites had cumulated 357 and 36 m
3/ha respectively, for a productivity gap of 321 m
3/ha. From year 8 to year 13, the productivity gap has also widened between sites with moderate yields (Ste-Catherine, La Patrie and Melbourne) and sites with high yields (Bedford and Brompton).
Our results also suggest that some poplar genotypes are generalists, while others appeared to be specialists, a trend that was equally observed in other studies [
13,
24]. In other words, clone selection is very important to optimize wood or biomass production along environmental gradients. Clones having a
P.
maximoviczii parentage (MxB-915311, DNxM-915508, NxM-3729) were generalists, because they had the smallest yield variation across the studied gradient, a trend equally observed in Denmark [
24]. In addition, when yield at the end of the 8th and 13th growing season were compared,
P.
maximoviczii hybrids had the greatest yield increase on higher elevation and/or poorer sites (Ste-Catherine, La Patrie, Melbourne, Fitch Bay, Ham and Stornoway) (
Table 1 and
Table 6). On the other hand, Euramerican hybrids (clones TxD-3230 and DxN-3570) were specialists, being only productive on low elevation fertile sites (Bedford and Brompton). In addition, at the Bedford site, the yield of clone DxN-3570 showed the greatest increase (+3.6 t/ha/year) when 8 and 13 years yield data were compared. Consequently, at this site, clone DxN-3570 was the least productive at year 8, but was among the most productive ones at year 13 (
Table 6). This suggests that important productivity gains can be obtained by managing clone DxN-3570 on longer rotations on bottomland sites. However, a longer rotation (13
vs. 8 years) only leads to marginal or nil yield gains for this clone on the other sites (
Table 6). The same could be said about clone TxD-3230, which experienced the second greatest productivity gain at Bedford (+2.0 t/ha/year) and the greatest productivity gain at Brompton, (+3.5 t/ha/year) (
Table 6), the other low elevation fertile site (
Table 1). However, on higher elevation sites gains for the TxD-3230 clone were marginal.
Table 7.
Site effect for total stem volume and woody biomass production and mean annual volume and biomass yield for 8 and 13 year-old poplar plantations. The volume and biomass increases from year 8 to 13 is indicated. Data for 8 year-old plantations were taken from Truax
et al. [
11].
Table 7.
Site effect for total stem volume and woody biomass production and mean annual volume and biomass yield for 8 and 13 year-old poplar plantations. The volume and biomass increases from year 8 to 13 is indicated. Data for 8 year-old plantations were taken from Truax et al. [11].
| Sites | Total Volume (m3/ha) | Increase (m3/ha) | Volume Yield (m3/ha/yr) | Total Woody Biomass (t/ha) | Increase (t/ha) | Woody Biomass Yield (t/ha/yr) |
|---|
| | 8 Years | 13 Years | | 8 Years | 13 Years | 8 Years | 13 Years | | 8 Years | 13 Years |
| Bedford | 179 | 357 | 178 | 22.4 | 27.5 | 78.7 | 148 | 69.5 | 9.8 | 11.4 |
| Brompton | 125 | 295 | 170 | 15.7 | 22.7 | 57.1 | 124 | 67.3 | 7.1 | 9.6 |
| Ste-Catherine | 68 | 181 | 113 | 8.5 | 13.9 | 33.7 | 79 | 45.8 | 4.2 | 6.1 |
| La Patrie | 75 | 174 | 99 | 9.4 | 13.4 | 36.9 | 77 | 40.3 | 4.6 | 5.9 |
| Melbourne | 46 | 151 | 105 | 5.8 | 11.6 | 23.9 | 68 | 43.8 | 3.0 | 5.2 |
| Fitch Bay | 24 | 96 | 72 | 3.1 | 7.4 | 13.4 | 44 | 30.1 | 1.7 | 3.3 |
| Ham | 27 | 79 | 51 | 3.4 | 6.1 | 15.7 | 37 | 21.6 | 2.0 | 2.9 |
| Stornoway | 8 | 36 | 28 | 1.1 | 2.8 | 6.4 | 19 | 12.4 | 0.8 | 1.4 |
| SE | 5 | 11 | - | 0.6 | 0.9 | 2.1 | 5 | - | 0.3 | 0.4 |
| p | <0.001 | <0.001 | - | <0.001 | <0.001 | <0.001 | <0.001 | - | <0.001 | <0.001 |
These trends may be related to the greater cold hardiness of
P.
maximoviczii hybrids compared to
P.
deltoides hybrids. In their natural habitats, poplars from the
Tacamahaca section such as
P.
maximoviczii and
P.
balsamifera, are widely distributed in the northern latitudes, growing to the latitudinal limits of trees [
54]. Conversely, poplars from the
Aigeiros section, such as
P.
deltoides, are better adapted to the riparian habitats of the bottomlands of temperate and arid regions [
54,
55]. This could explain the lower survival and yields of clones DxN-3570 and TxD-3230, compared to the
P.
maximoviczii hybrids, on high elevation sites such as Stornoway, Melbourne and La Patrie (
Table 6). Despite this lower survival, clone TxD-3230 has produced relatively large trees on these sites (
Table 5). However, many of these large trees were heavily damaged by sunscalds (B. Truax and J. Fortier, field observations), reflecting the vulnerability of this clone to cold damages. Clones DxN-3570 and TxD-3230 were also more heavily browsed by deer following planting, which may have reduced their survival and early growth [
11]. Consequently, when planted in more stressful environments, in terms of climate, soil fertility and herbivore pressure, these two clones were less productive than
P.
maximoviczii hybrids.
From a physiological perspective, the very low productivity of clone DxN-3570 on higher elevation sites suggests that it may be poorly adapted to soil conditions prevailing in colder sites (lower N mineralization rate). In a previous field experiment involving the same poplar genotypes, clone DxN-3570 had the greatest leaf nitrate reductase activity (NRA), and was the only clone that showed an increase in NRA in response to a soil increase in nitrate (NO
3) availability [
56]. This evidence suggested that this clone has a greater ability for NO
3 assimilation in its leaves than the other clones, potentially reflecting its preference for soil NO
3 over NH
4, as also observed for
P.
deltoides [
18,
57]. In temperate bottomland habitats, soil NO
3 is generally the dominant N-form in soils with a good drainage [
58,
59]. This could explain why clone DxN-3570 was among the most productive clones at the Bedford site, the only rich bottomland site in this study. On all the other sites, clone DxN-3570 was the least productive clone (
Table 5 and 6
Table 6), reflecting its specialization for warm and rich bottomland sites.
Although clone MxB-915311 was a generalist clone in this study, it may not be the best choice in rich bottomland sites since its yield at the Bedford site (20.5 m
3/ha/yr) was much lower than that of the other clones (26.1–31.4 m
3/ha/yr). Having both of its parental species belonging to the
Tacamahaca (balsam poplar) section, clone MxB-915311 may be more adapted to colder and wetter sites where the dominant soil N-form is often NH
4. Its parental species
P.
balsamifera was also found to prefer soil NH
4 over NO
3 [
15], but also to have the ability to short-circuit the mineralization step of decomposition by absorbing directly amino-acids in environments with low
N-mineralization rates and low inorganic N stocks (cold climates and wet soils) [
60,
61].
In addition to potential differences in cold hardiness and soil N-form preferences of the studied genotypes, other factors could explain why some clones performed better than others on lower fertility sites. Several endophytic bacteria from poplars can promote the growth of their host plant by providing N-fixation, growth regulators or by preventing the growth or activity of plant pathogens [
62,
63]. Poplar growth can also be promoted by the colonization of fungal endophytes, but also ectomycorrhizal and arbuscular mycorrhizal fungi [
64]. The effects of those symbiotic associations have been found to be strongly influenced by poplar genotypes, as well as environmental conditions [
64,
65]. Consequently, across the environmental gradients studied, some genotypes may have benefited from those symbiotic associations more than others.
After eight years, clone NxM-3729 had the greatest productivity and survival rate on all sites across the climate and site fertility gradient studied [
11]. Five years later, this clone was still among the most productive ones at most sites (
Table 5 and 6
Table 6). However, at the fertile site of Bedford, clone NxM-3729 fell in 2nd position for biomass yield and in 4th position for volume yield. At this site, several trees of this clone showed symptoms of
Septoria musiva (stem canker) infection (B. Truax and J. Fortier, field observations), which may have increased tree mortality and lowered its growth rate. At Bedford, clone NxM-3729 was the only clone to have a lower biomass yield at 13 years than at 8 years (
Table 6). This situation highlights the need for planting a diversity of clones, in mono-specific blocks, in order to increase plantation resilience, while avoiding interspecific competition [
66,
67].
3.4. Management Implications
In temperate regions, short rotation woody crop yields should ideally fall within the range of 10–30 m
3/ha/yr, depending on site quality, clones, local climate and cultivation methods [
68]. Taking these numbers as a bench mark, our study suggests that 5 of our study sites out of 8 had produced a satisfactory wood volume after 13 years. On these 5 best sites, wood and biomass production ranged from 151 m
3/ha (or 68 t/ha) at Melbourne up to 357 m
3/ha (or 148 t/ha) at Bedford (
Table 7). Higher yields could have been obtained on all of these 5 sites by selecting the clones that were better adapted to site characteristics, as revealed by the significant Site × Clone interactions (
Table 5 and
Table 6). Although some genotypes are generalists, being able to produce moderate to high yields on a variety of sites, clone selection is still very important to optimize yields across environmental gradients.
Despite the fact that the 3 low productivity sites (Fitch Bay, Ham, Stornoway) had increased their mean annual increments or yields from year 8 to year 13 (
Table 7), they still have a marginal productivity after 13 years. While some genotypes were more productive than others on these low yielding sites, clone selection alone cannot compensate for poor site selection, as equally observed in South Carolina, United States [
27]. In other words, not all abandoned farmland sites of southern Québec are suitable for poplar cultivation, which corroborates similar observations in France [
69].
It could be argued that poplar plantations should be managed on longer rotations on these low productivity sites. However, hybrid poplars are short-lived trees because they are highly susceptible to diseases and pest outbreak [
16]. In addition, hybrid poplars tend to have low mechanical strength properties [
70], which make them susceptible to stem breakages during severe climatic events such as windstorms or ice storms. Since those climatic events are episodically observed in the southern Québec region, a low risk management approach for the production of pulpwood and sawlogs would be the use of high quality sites, with intermediate rotation length (12–15 years) for planting densities around 833 stem/ha. At the beginning of the 15th growing season, we observed severe damages (broken crowns, bent trees, uprooted trees) at the La Patrie plantation caused by climatic events, which suggests that long rotations (more than 15 years) might be risky for the studied region. Besides, time did not play in favor of the low yielding sites since the productivity gap between the more and less productive sites has widened over the years (
Table 7). If biomass for bioenergy is also an objective, a planting density reaching up to 2500 stems/ha could be used, with half of the trees being thinned earlier in the rotation, allowing the remaining trees to reach pulpwood diameter for the final harvest, as recommended in Sweden [
26].
The allometric relationships developed in this study, as well as the branch biomass ratio calculated for the different clones at each site (
Figure 1 and
Table 6), suggest important genotypic variation in patterns of biomass allocation to branches. Our observations showed that
P.
maximoviczii hybrids allocated a larger proportion of their woody biomass to branches compared to clones DxN-3570 and TxD-3230. This trait of
P.
maximoviczii hybrids may not be desirable when the objective is to produce higher quality products (veneer or sawlogs) since pruning trees with greater branch biomass will be more time-consuming or expensive. On the other hand, the greater branch biomass proportion of
P.
maximovizcii hybrids favors rapid canopy closure, which is important to restore key forest understory attributes such as shade and low herbaceous cover [
6,
71,
72].
Finally, this study suggests that clone-specific and age-specific allometric relationships will provide more accurate estimates of wood volume and woody biomass. Increasing the accuracy of volume and biomass estimates of standing trees could be especially helpful in supporting management decisions regarding the timing of harvest or thinning operations.


{kind=link}
{kind=link}

