1. Introduction
Understanding a species’ habitat selection is a major prerequisite for wildlife management, concerning both conservation and forest management. Habitat selection can be analyzed at a very fine spatio-temporal resolution using GPS telemetry, which provides location data of cryptic organisms on an almost continuous scale [
1]. Indeed, the availability of environmental covariates at an adequate spatial resolution to resolve site selection at the scale of the GPS telemetry data is an important methodological challenge, as conventional field measurement techniques require a prohibitively high effort when covering large areas [
2]. High-resolution remote sensing, such as airborne Laser scanning (LiDAR, Light detection and ranging), promises to provide the unique possibility to describe forest structure in high detail and over large areas [
3].
Here, we analyze winter habitat selection of European roe deer using metrics of forest structure derived from airborne LiDAR. The European roe deer is a medium sized ungulate that is distributed throughout Europe and adapted to a wide variety of landscape types [
4], with a preference for wooded habitats [
5]. Roe deer are selective feeders that browse on a large number of plant species [
6,
7]. By selecting palatable species, roe deer can greatly influence the composition of tree regeneration, in particular in areas where their natural enemies are missing [
8,
9]. In northern latitudes and parts of Continental Europe, animals have to face extreme winter conditions that strongly influence roe deer survival [
10,
11]. Winter is, in general, a time of food shortage, both in quantity and quality and animals lose a lot of energy to compensate for the thermal loss, due to low temperatures or wind chill. Additionally, deep snow restricts food availability and moving abilities, especially when snow cover exceeds a height of 50 cm [
12].
LiDAR is an established remote sensing technique that provides accurate, high-resolution data for the assessment of three-dimensional vegetation structure over large areas [
13]. In addition to many applications relevant for forest management [
14], airborne LiDAR is increasingly used in wildlife-habitat studies [
15,
16,
17]. Although Coops
et al. [
18] underlined the utility of airborne LiDAR to map potential winter habitat of mule deer (
Odocoileus hemionus), to our knowledge, no study exists that utilized this technique to analyze movement behavior of forest habitat species.
Knowing the behavioral mode of an animal is a prerequisite to better understand an animals’ habitat use [
1]. For this purpose, acceleration sensors integrated in modern GPS collars are a promising technology for collecting behavioral data in wildlife habitat studies. These sensors are able to quantify activity intensity at short time intervals, and, thus, deliver information on the activity status of the animals,
i.e., if they are active or resting, on a quasi continuous basis [
19]. This technique has been shown to be able to distinguish between locomotion, foraging, and resting for roe deer and red deer (
Cervus elaphus) [
19,
20]. Here, we utilize this technique to investigate differential habitat selection between active and resting animals.
We used GPS telemetry data of 15 individuals, and analyzed site selection depending on LiDAR derived estimates of vegetation cover in different height strata, using step selection functions (SSF). SSF compare the environmental conditions at locations of realized movement steps to the conditions at locations the animal could have alternatively chosen at a given step [
21].
Specifically, our study aimed at utilizing LiDAR-derived measures of forest structure for testing three predictions on the winter habitat selection of roe deer in a mountain area with harsh weather conditions: We predicted that in winter: (1) Roe deer should prefer semi open habitats with short distances between vegetation structure providing suitable shelter and food; (2) With increasing harsh weather conditions (temperature, snow height) roe deer should prefer sites with dense canopy cover due to lower heat emission and lower snow levels; and (3) Sites used for resting should be characterized by higher canopy cover than sites visited by active roe deer due to higher thermal cover.
2. Experimental Section
2.1. Study Area
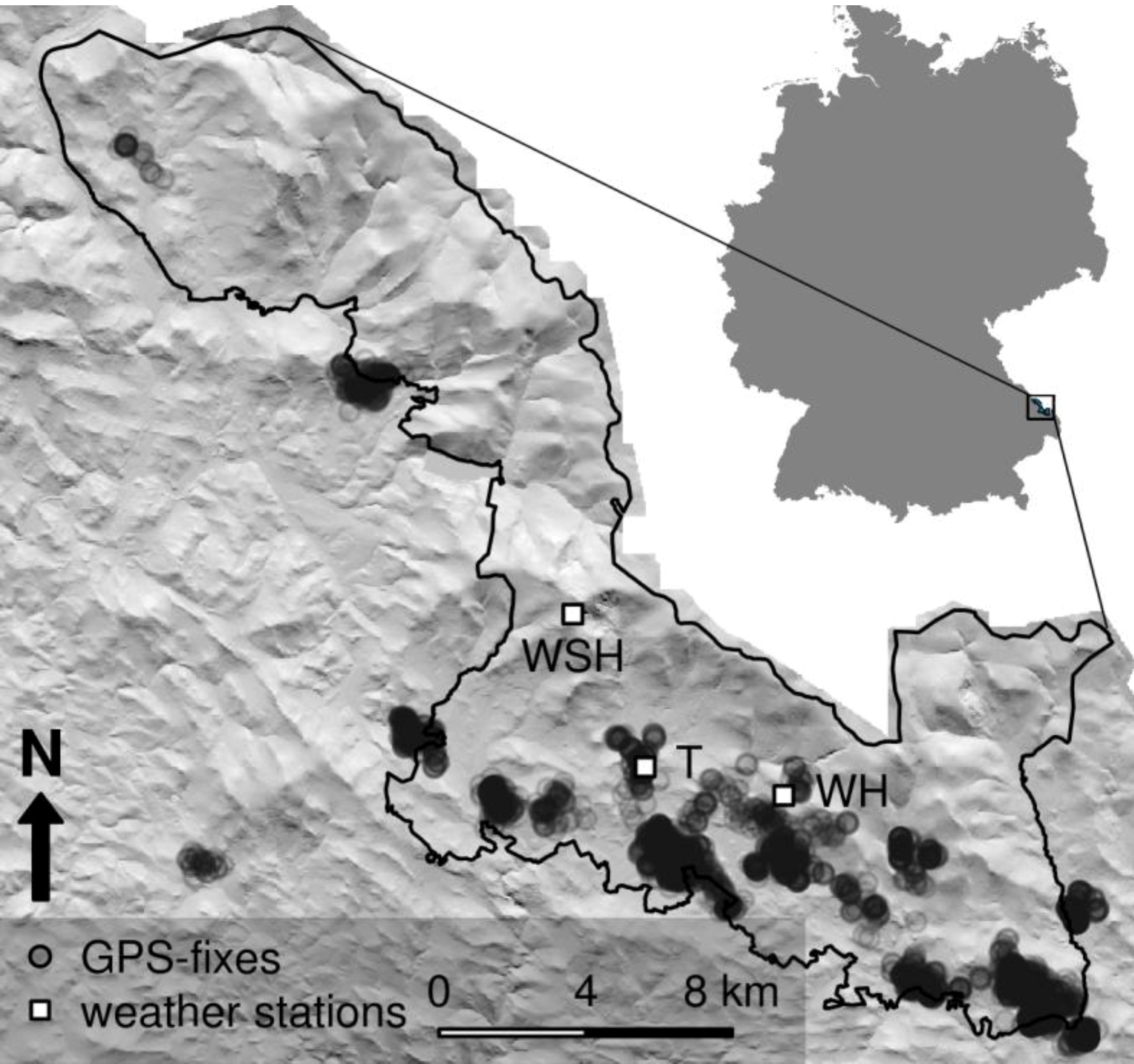
The study was conducted within the Bavarian Forest National Park (
Figure 1), situated in Southeastern Germany along the border of the Czech Republic (center coordinates 49°3′19″ N, 13°12′9″ E). It covers an area of 24,369 hectares; elevation ranges from 600 to 1453 m above sea level (a.s.l.). Three major forest types characterize the vegetation: High altitudes are dominated by Norway spruce (
Picea abies) forests, the slopes in medium elevations are dominated by mixed montane forests, mainly consisting of beech (
Fagus sylvatica), fir (
Abies alba), and Norway spruce, and depressions in low altitudes are dominated by alluvial spruce forests, mainly consisting of Norway spruce and birches (
Betula pendula and
Betula pubescens). Mean annual temperature ranges from 2 °C to 7.3 °C and annual precipitation from 830 to 2280 mm between valley bottoms and mountaintops. Snow cover lasts on average 139 days per year in medium altitudes with, on average, 50 days per year with snow depths higher than 50 cm [
22]. Roe deer use the whole area of the national park in summer. In winter, they move to lower altitudes, where snow is less deep, and some individuals use the adjacent areas of the national park where they can find artificial feeding places on private hunting grounds. Within the national park, artificial feeding places are available in and around traps that are used to catch individuals for research purposes. Food mainly consists of apple pomace, occasionally also maize and silage, and is provided daily to keep animals close to the traps. Hunting is prohibited within the national park. As a predator of roe deer, the Eurasian lynx (
Lynx lynx) occurs in the national park [
23].
Figure 1.
Study area and its location in Germany; The Bavarian Forest National Park is outlined in black; GPS-fixes of roe deer used in the analysis are shown as black circles, locations of weather stations as squares; T: Taferlrück, WH: Waldhäuser, WSH: Waldschmidthaus.
Figure 1.
Study area and its location in Germany; The Bavarian Forest National Park is outlined in black; GPS-fixes of roe deer used in the analysis are shown as black circles, locations of weather stations as squares; T: Taferlrück, WH: Waldhäuser, WSH: Waldschmidthaus.
2.2. Telemetry Data
Roe deer were caught using wooden box traps. After fitting of GPS-collars animals were quickly released. GPS Telemetry data was recorded using GPS-GSM collars with integrated activity sensors (series 3.000 from VECTRONIC Aerospace, Berlin, Germany). Locations of roe deer were recorded with different time intervals, mainly ranging from 15 min to two hours between successive fixes. Individual animals were tracked for time periods ranging from 14 days to six months during the wintertime. Data from fawns and individuals with more than 10% missing positions due to GPS-errors were excluded from the analysis. For the remaining data, we removed recordings if velocities between successive locations exceeded 6.5 m/s. In order to get an equally spaced time series of roe deer locations, we only retained GPS-fixes with a time interval of 4 h. We checked independence of locations between individuals by visually inspecting movement trajectories. Furthermore, we restricted recorded locations to the leaf-off period from November to the end of April. This resulted in 1418 locations of nine adult females and six adult males recorded between April, 2007, and December, 2010, for the analysis. GPS-collars were equipped with activity sensors that are able to capture the animals’ locomotion by measuring acceleration in forward/backward and sideward motion. Acceleration is measured six to eight times a second and averaged over a fixed time period, in this case over 5 min. Values were recorded on a linear scale between 0 and 255. In order to identify the activity status of individuals we used threshold levels for activity values that had been determined in a separate study [
20], conducted in the same study area. Individuals were considered as active when the sum of both acceleration values (forward/backward and sideward) was greater than 4, and as resting otherwise.
2.3. Weather Data
We used temperature and wind speed data from three weather stations Taferlruck (769 m a.s.l.), Waldhäuser (940 m a.s.l.), and Waldschmidthaus (1356 m a.s.l.) (
Figure 1) recorded in 10 min time intervals, and daily-recorded snow height data from the Waldhäuser weather station. Temperature and wind speed values for each roe deer position were taken from the nearest weather station. In order to exclude effects of daytime on habitat selection we calculated a generalized additive model (GAM) with temperature as response, and hour of day as predictor variable, and used the residuals for further analysis. This resulted in a variable representing the deviation from mean temperature at the corresponding daytime a GPS-fix was recorded.
2.4. LiDARData Acquisition
LiDAR data were provided by the “Bavarian State Office for Land Survey and Geo-information”. Data of the study area were acquired in leaf-off period between April, 2008, and November, 2009, using an airborne Riegl LMS-Q 560 (RIEGL Inc., Horn, Austria) system. The system operated with a wavelength of 1550 nm and recorded first and last return points with a vertical error of +/−0.16 m. Flight height was between 1194 and 2306 m a.s.l. (average height above ground: 776 m), and average flight velocity was 55 m/s. This resulted in a mean swath width of 832 m, a footprint with mean diameter of 38.8 cm, and an average point density of 9.8 points/m². Classification into ground and vegetation returns was performed using TerraScan (TerraSolid Ltd., Helsinki, Finland). A digital terrain model (DTM) in 1-m resolution was calculated with SCOP++ (inpho GmbH, Vienna, Austria) using “adaptable prediction” as interpolation method. The height of LiDAR returns above ground was calculated by subtracting the height value of the underlying DTM from the height of each point.
The resulting 3D point cloud, representing discrete returns of the land surface and vegetation cover, was used to calculate point statistics. Various metrics, describing different aspects of the vegetation structure, can be calculated [
24]. Estimates of fractional vegetation cover, defined as the projection of the tree crowns onto the ground divided by ground surface area, can be calculated from point ratios in different strata [
14]. Most applications of airborne LiDAR to derive fractional cover were focused on tree canopy cover, which can be estimated with high accuracy [
25,
26,
27]. Some attention has also been paid on the estimation of understory cover in forests [
28,
29,
30]. Wing
et al. [
31] received good results for estimating understory cover from high-resolution small-footprint LiDAR data.
We calculated fractional vegetation cover of three different strata; understory (0.5 m–2 m above ground), midstory (2 m–10 m above ground), and overstory vegetation cover (10 m–60 m above ground) (
Figure 2). For this purpose points were aggregated into a 5 m × 5 m grid and values were calculated for each grid cell using the following formula:
where
vch12 represents an estimate of vegetation cover between height above ground
h1 and
h2 (
h1 <
h2), while
nh1 and
nh2 represent the number of LiDAR return points below height
h1 and below height
h2, respectively. The variables
h1 and
h2 represent the lower and upper height border of a stratum.
The estimation of fractional cover of vegetation directly above the ground is subject to errors, due to the imprecision of the DTM. A previous study conducted in the Bavarian Forest National Park revealed a root mean square error of 0.3 m for the DTM [
32]; therefore, we did not estimate fractional cover below 0.5 m.
Figure 2.
Vertical cross section of LiDAR point cloud data and derived fractional cover values for under-, mid- and overstory along an illustrative 50 m transect of 5 m width.
Figure 2.
Vertical cross section of LiDAR point cloud data and derived fractional cover values for under-, mid- and overstory along an illustrative 50 m transect of 5 m width.
2.5. Statistical Analysis
To analyze habitat selection of roe deer, we applied a step selection function (SSF), a special form of a resource selection function (RSF) [
33,
34]. In the case of an SSF, control points are sampled based on empirical data of the monitored animals’ movement behavior. The direct spatial connection between two successive GPS-fixes is thereby denoted as a step. For each observed step, a certain number of control steps are generated using the empirical distribution of step-lengths and turning angles between successive steps of the monitored animals. To analyze habitat preferences, the characteristics of observed steps from an individual are compared with that of control steps sharing the same starting point. In this study, we compared different values of vegetation cover at the endpoint of each step. By using a conditional logistic regression model with random effects, we accounted for between-individual heterogeneity. Additionally, we tested whether site selection of individuals is influenced by the proximity to the nearest artificial feeding site. Data processing and statistical analysis were carried out using R version 2.14.0 [
35].
For the generation of random steps, we jointly sampled step lengths and turning angles from the distributions of observed steps lengths and angles. Step length is defined as the distance between the start and endpoint of a step and the turning angle as the angular deviation of a step’s direction relative to the direction of the previous step. As step lengths of male roe deer were significantly longer compared to females, values were calculated separately for these two groups. The length and turning angle of the observed step for which random steps had to be generated, were excluded from sampling. Following the framework of Fortin
et al. [
33], we generated 25 random steps for each observed step.
LiDAR-derived vegetation cover values were extracted from raster cells within a buffer of 5 m around observed and random locations. This buffer size was selected to compensate for possible imprecision in positioning of the animals. Extracted values were averaged for each location. To check the presence of collinearity between explanatory variables, we calculated Spearman’s rank correlation coefficient considering all possible pair wise variable combinations. All correlation coefficients were smaller than 0.5, indicating low collinearity between explanatory variables.
In order to compare characteristics of selected and random locations we fitted a mixed effects conditional logistic model with vegetation cover values as explanatory variables and individuals as random effects using the coxme function in the R package coxme [
36]. Four separate models were fitted, differing in whether (1) both weather variables (temperature deviation, snow height, and wind speed) and activity information; (2) only weather; (3) only activity; or (4) neither were included (
Table 1). Initial models included the linear and quadratic effects of vegetation cover, as well as their two-way interactions with the activity status. Models including weather conditions as co-variables additionally consisted of interaction terms between the terms described in the initial model and the three used weather variables. Furthermore, distance to the nearest feeding place was included as explanatory variable in all models.
In order to find minimum adequate models, an AIC (Akaike’s Information Criterion) based stepwise backward selection was performed.
Table 1.
Summary of calculated mixed conditional logistic regression models explaining habitat selection and associated Akaike’s Information Criterion(AIC) values; model 1 incorporates weather variables and activity status, model 2 only weather variables, model 3 only activity status, and model 4 neither of the two; all models incorporated individuals as random effect; dist_to_f: distance to nearest artificial feeding place, OC: overstory cover, MC: midstory cover, UC: understory cover, temp: deviation from mean temperature at corresponding daytime, act: activity status of roe deer, snow: snow height, wind: wind speed.
Table 1.
Summary of calculated mixed conditional logistic regression models explaining habitat selection and associated Akaike’s Information Criterion(AIC) values; model 1 incorporates weather variables and activity status, model 2 only weather variables, model 3 only activity status, and model 4 neither of the two; all models incorporated individuals as random effect; dist_to_f: distance to nearest artificial feeding place, OC: overstory cover, MC: midstory cover, UC: understory cover, temp: deviation from mean temperature at corresponding daytime, act: activity status of roe deer, snow: snow height, wind: wind speed.
| Model | Included Variables | Fixed effects | AIC |
|---|
| 1 | LiDAR + activity + weather | OC + UC + MC + Dist_to_f + OC² + MC² + OC:act + UC:act + MC²:act + MC:temp + OC²:temp + OC:snow + MC:snow + OC²:snow | 9346 |
| 2 | LiDAR + weather | OC + UC + MC + Dist_to_f + OC² + MC² + UC:temp + MC:temp + OC²:temp + OC:snow + MC:snow + OC²:snow + MC²:snow + UC:wind + MC:wind + MC²:wind | 9362 |
| 3 | LiDAR + activity | Dist_to_f + OC + MC + UC + OC² + OC²:act + MC:act + C:act | 9394 |
| 4 | LiDAR | OC + MC + dist_to_f + OC² | 9421 |
We used a re-sampling procedure to determine the importance of explanatory variables in the selected models [
37]: We compared the prediction of the SSF calculated from original data to the prediction from randomized values of the explanatory variables. For this purpose, we first calculated the predicted values of the SSF with given values of the explanatory variables for each observed location. Then, we calculated predicted values of the SSF randomizing each time one of the explanatory variables. Finally, the Pearson correlation between predictions, based on the randomized datasets and the original data, was calculated. Smaller correlation coefficients indicate a higher influence of the permuted variable.
3. Results
Male roe deer moved significantly longer distances (Wilcoxon test, p < 0.001, mean: 208 m in 4 h) than females (mean: 171 m in 4 h). Mean snow height for all GPS fixes used in the analysis was 26.3 cm. Maximum snow height was 105 cm, recorded in February 2010. Mean temperatures during the study period ranged from −4.7 °C in the night to 1.3 °C at noon. Mean observed wind speed was 1.5 m/s. Individuals stayed on average distances of 250 to 1200 m away from the nearest artificial feeding site.
Integration of weather conditions as well as including the activity status of the animals improved tested models on habitat selection of roe deer (
Table 1). Model 1 containing interactions of weather conditions and activity status performed best as measured by AIC,
i.e., it has the best expected predictive performance on new observations at the same environmental conditions (AIC: 9346), followed by the models 2 and 3 only considering weather conditions (9362) or activity status (9394) as interactions. All following results refer to model 1, including weather variables and activity status of the animals.
The activity status of roe deer influenced habitat selection patterns (
Table 2). Generally, active roe deer selected sites characterized by lower canopy (overstory and midstory) cover and higher understory cover than sites selected for resting (
Figure 3,
Figure 4 and
Figure 5).
Table 2.
Results of mixed conditional logistic regression with individuals as random effect (Model 1); bold values indicate significant effects (p< 0.05) of model terms; dist_to_f: distance to nearest artificial feeding place, OC: overstory cover, MC: midstory cover, UC: understory cover, temp: deviation from mean temperature at corresponding daytime, act: activity status of roe deer, snow: snow height.
Table 2.
Results of mixed conditional logistic regression with individuals as random effect (Model 1); bold values indicate significant effects (p< 0.05) of model terms; dist_to_f: distance to nearest artificial feeding place, OC: overstory cover, MC: midstory cover, UC: understory cover, temp: deviation from mean temperature at corresponding daytime, act: activity status of roe deer, snow: snow height.
| Variable | Coef. | SE | Z | P |
|---|
| Dist_to_f | −0.002 | <0.001 | −4.54 | <0.001 |
| OC | 2.721 | 0.544 | 5.000 | <0.001 |
| MC | 1.341 | 0.547 | 2.45 | 0.014 |
| UC | −1.216 | 0.531 | −2.29 | 0.022 |
| OC² | −2.902 | 0.584 | −4.97 | <0.001 |
| MC² | −1.805 | 0.789 | −2.29 | 0.022 |
| OC:act | −0.764 | 0.204 | −3.74 | <0.001 |
| UC:act | 1.812 | 0.717 | 2.53 | 0.011 |
| MC:act | −0.911 | 0.385 | −2.36 | 0.018 |
| OC²:temp | −0.092 | 0.024 | −3.81 | <0.001 |
| MC:temp | −0.088 | 0.039 | −2.25 | 0.024 |
| OC:snow | −0.038 | 0.014 | −2.68 | 0.008 |
| MC:snow | 0.030 | 0.007 | 4.27 | <0.001 |
| OC²:snow | 0.052 | 0.015 | 3.37 | <0.001 |
Figure 3.
Predicted relative preference values for overstory cover (2 m–10 m) and midstory cover (10 m–60 m) as a function of temperature deviation from mean temperature values at given daytime for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to their data set mean. Preference values were scaled between 0 and 1 to keep results between different temperatures comparable.
Figure 3.
Predicted relative preference values for overstory cover (2 m–10 m) and midstory cover (10 m–60 m) as a function of temperature deviation from mean temperature values at given daytime for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to their data set mean. Preference values were scaled between 0 and 1 to keep results between different temperatures comparable.
Figure 4.
Predicted relative preference values for overstory cover (10 m–60 m) and midstory cover (2 m–10 m) as a function of snow height for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to their data set mean. Preference values were scaled between 0 and 1 to keep results between different temperatures comparable.
Figure 4.
Predicted relative preference values for overstory cover (10 m–60 m) and midstory cover (2 m–10 m) as a function of snow height for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to their data set mean. Preference values were scaled between 0 and 1 to keep results between different temperatures comparable.
Figure 5.
Predicted preference values for understory cover for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to the corresponding data set mean.
Figure 5.
Predicted preference values for understory cover for active and resting roe deer; values represent expected values estimated by mixed conditional logistic regression; not-displayed variables were set to the corresponding data set mean.
While temperature and snow height influenced the preference for over- and midstory cover (
Table 2), we found no influence of wind speed on roe deer habitat selection. Roe deer preferred higher over- and midstory cover when temperature decreased (
Figure 3). At temperatures more than 5 °C above average daytime temperatures, animals preferred more open habitats characterized by low to medium over- and midstory cover (
Figure 3). With increasing snow height, roe deer also preferred increasing midstory cover (
Figure 4). Additionally, animals strongly selected sites characterized by high values of overstory cover when snow height exceeded 60 cm (
Figure 4).
Distance to the nearest artificial feeding place was the most influencing variable describing habitat selection of roe deer (relative importance: 0.53). The most influencing cover value was overstory cover (relative importance: 0.104) followed by midstory cover (0.088) and understory cover (0.031). Comparison of used weather variables and activity data revealed that activity status was the most influencing variable (relative importance: 0.098), followed by temperature (0.09) and snow height (0.062).
4. Discussion
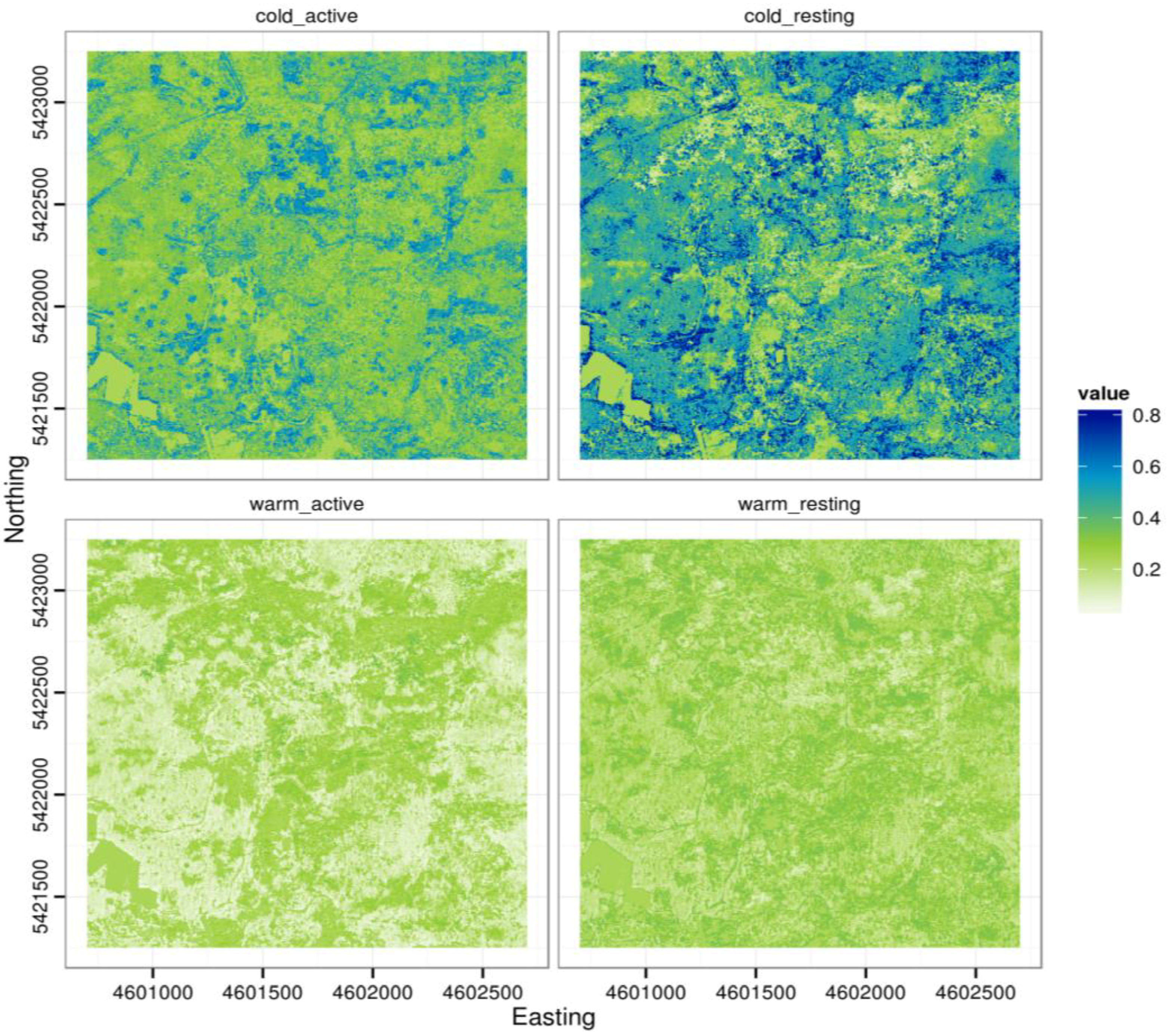
Winter habitat selection of roe deer strongly depended on the animal’s activity status and on weather conditions. Specifically, we found some general habitat selection patterns: Roe deer either tended to use sites characterized by at least some degree of canopy cover, or sites characterized by high understory cover (
Figure 6). Thus, roe deer avoided open sites in an area with harsh winter conditions. Roe deer primarily use open areas to forage and, under harsh winter conditions, we expect no food to be available there. Our findings are in line with Mysterud
et al. [
38], who also found roe deer avoiding open habitats in winter. Similarly, under high snow conditions, browse availability was found to be less important for relative patch use of white-tailed deer (
Odocoileus virginianus) [
39,
40], and only shelter-related features were predictive of variation in habitat use [
40].
Animals preferred higher canopy cover when it was cold compared to average temperatures or when snow heights of our reference weather station exceeded 60 cm. This result is consistent with previous studies on ungulates in general [
41,
42,
43,
44], and roe deer in particular [
45,
46]; these studies also observed a preference for higher canopy cover at low temperatures and in deep snow. The result confirms our expectations that roe deer use forests with high canopy cover as thermal shelter when temperatures are low.
We found no effect of wind speed on roe deer winter habitat selection. In a previous study, increased use of dense canopy cover was observed with increasing wind speed for roe deer [
46]. The weak response in our study is probably due to a lack of information in the used weather data. Most of the observed wind speeds, at the time of GPS-fixes, were below 5 m/s, so that little information was available on behavior at extreme wind conditions. Additionally, wind exposure strongly depends on terrain and thus can locally differ considerably from values measured at weather stations.
Figure 6.
Maps showing predicted values of roe deer habitat preference for different scenarios of weather conditions and activity status; prediction maps where calculated using model 1 (
Table 2), keeping the distance to the nearest feeding place constant at the dataset mean; cold_active: prediction for active roe deer at temperatures kept constant at the 10th percentile and snow height kept constant at the 90th percentile of observed values; cold_resting: resting roe deer, 10th percentile temperature value, 90th percentile snow height value; warm_active: active roe deer, 90th percentile temperature value, no snow; warm_resting: resting roe deer, 90th percentile temperature value, no snow. Map projection: DHDN, Gauss-Krüger Zone 4.
Figure 6.
Maps showing predicted values of roe deer habitat preference for different scenarios of weather conditions and activity status; prediction maps where calculated using model 1 (
Table 2), keeping the distance to the nearest feeding place constant at the dataset mean; cold_active: prediction for active roe deer at temperatures kept constant at the 10th percentile and snow height kept constant at the 90th percentile of observed values; cold_resting: resting roe deer, 10th percentile temperature value, 90th percentile snow height value; warm_active: active roe deer, 90th percentile temperature value, no snow; warm_resting: resting roe deer, 90th percentile temperature value, no snow. Map projection: DHDN, Gauss-Krüger Zone 4.
Overall, roe deer tended to use sites with lower canopy cover when active. This is concordant with the result of Ratikainen
et al. [
47], who observed that bed-sites were characterized by higher canopy cover than foraging sites. This suggests that especially resting individuals utilize the thermal cover of dense canopies. Additionally, we found roe deer preferring dense understory when active, and low understory cover when resting. The high preference of resting animals for forests, characterized by little undergrowth is reasonable to avoid encounters by potential predators. Especially lynx depend on the presence of understory vegetation to stalk their prey [
48]. On the other hand, active bouts of roe deer in winter are mainly used for foraging [
49]. Therefore, it is a reasonable result that individuals prefer sites characterized by higher understory cover, also indicating higher winter food supply, when active. Overall, our results underline the findings of Mysterud
et al. [
46], who could show that roe deer particularly in winter face a trade-off between food availability and thermal cover. However, our study shows that this finding should be extended by a second trade-off between food availability and protection from predators such as lynx.
Distance to the nearest artificial feeding place was the most important variable explaining winter movement behavior of adult roe deer. This result is in line with a study by Guillet
et al. [
50], who observed that roe deer tended to concentrate around artificial feeding places at extreme weather conditions in winter. This is not surprising as roe deer only store small fat reserves and individuals have to cover most of their energy by permanent food intake [
12]. Our finding underlines that low food availability is one of the major challenges roe deer face under harsh winter conditions. However, average distances to artificial feeding places in our study were fairly high compared to that observed by Guillet
et al. [
50]. Although most of the animals appeared at least once in close proximity to these places, animals typically kept a distance for most of the time. This finding can be explained by the small amount of food that is supplied only for the purpose of luring roe deer to the traps and that, thus, cannot be compared to a full winter-feeding.
Of the used LiDAR-variables, overstory cover was the most important variable, but did not differ strongly from midstory cover. Since roe deer showed no basic differences in usage of over- and midstory cover, we can assume that primarily the functional characteristics of these layers, such as reduced heat emission and snow interception, are important for the animals. In contrast, we found only little influence of understory cover. This result is in line with the findings by Torres
et al. [
51], who analyzed winter habitat selection in Norway and found no influence of shrub cover on habitat selection. In the Bavarian Forest National Park, the small influence may be explained by the presence of lynx. In this case, high understory cover serves as potential source of winter forage, but on the other hand enhances the risk of being killed.
In addition, the difference of importance between over- and understory could be influenced by the estimation accuracies. Using LiDAR with similar footprint and point density, as in this study, tree canopy cover can be estimated more accurately than understory cover [
25,
31].
5. Conclusions
Here we aimed at integrating high-resolution remote sensing data of forest structure and behavioral data on a forest-dwelling ungulate to study site selection, combining GPS-telemetry with a resource selection function approach. We could show that habitat selection of roe deer under rough winter conditions cannot be generalized, but is strongly dependent on external influences, like weather, and on the activity status of the animal. Thus, our study underlines the importance of animal behavioral data in habitat use studies. Moreover, our results show that roe deer in winter are dependent on heterogeneous forest structures. These may comprise forest regeneration with high understory cover, which provide potential forage for roe deer particularly at high snow, and older forest stands with dense canopy cover, which potentially mitigates the effect of cold temperatures and high snow.
High-resolution LiDAR data were well suited to depict forest structure relevant for habitat selection in general, and dependencies of site selection on activity status and weather variables in particular. This underlines the importance of physical forest structure for site selection of roe deer and suggests the increased application of LiDAR technology in wildlife-habitat studies of ungulates. Particularly, the use of continuous and area-wide available environmental covariates allows generating detailed prediction maps on habitat use which can be extrapolated beyond the borders of the study area and may inform forest conservation or wildlife management. Thus, we highly recommend the use of high-resolution remote sensing data and behavioral data in studies on animal habitat use.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



