
Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China


Abstract
:1. Introduction
2. Materials and Method
2.1. Study Area

2.2. Tree-Ring Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I.D. | Tree species | Topographic aspect | Latitude N | Longitude E | Elevation (m) | DBH a (cm) | Tree height a (m) |
|---|---|---|---|---|---|---|---|
| PN | P. meyeri | North-facing | 38°43′14.6″ | 111°51′16.2″ | 2600–2660 | 17.3 ± 4.8 | 10.0 ± 2.1 |
| LN | L. principis-rupprechtii | North-facing | 38°43′14.6″ | 111°51′16.2″ | 2600–2660 | 15.9 ± 4.5 | 8.8 ± 3.2 |
| PS | P. meyeri | South-facing | 38°42′44″ | 111°51′33.5″ | 2540–2620 | 19.3 ± 4.2 | 13.4 ± 3.4 |
| LS | L. principis-rupprechtii | South-facing | 38°42′44″ | 111°51′33.5″ | 2540–2620 | 26.2 ± 4.0 | 16.0 ± 3.4 |
2.3. Chronology Development
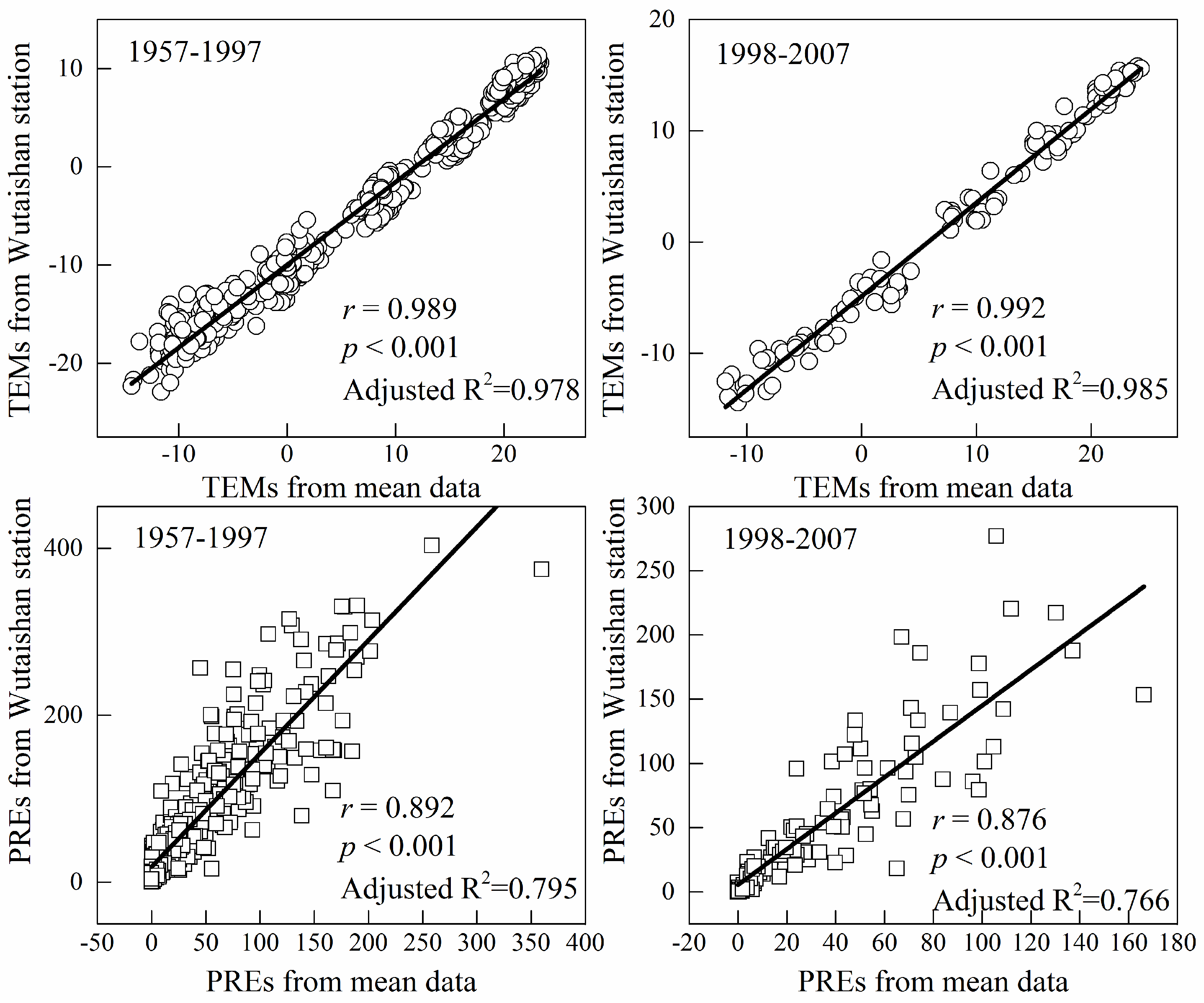
2.4. Climate Data

2.5. Data Analysis
3. Results
3.1. Characteristics of Tree-Ring Chronologies
| I.D. | Trees/Cores | Time span | MS | SD | Rbar | PC1% | SNR | EPS |
|---|---|---|---|---|---|---|---|---|
| PN | 18/36 | 1947–2007 | 0.166 | 0.134 | 0.409 | 49.0 | 15.679 | 0.940 |
| LN | 16/32 | 1940–2007 | 0.171 | 0.161 | 0.509 | 58.0 | 18.273 | 0.948 |
| PS | 17/34 | 1909–2007 | 0.140 | 0.120 | 0.365 | 46.6 | 18.807 | 0.950 |
| LS | 16/32 | 1931–2007 | 0.182 | 0.164 | 0.457 | 51.0 | 25.178 | 0.962 |
| PICEA | 35/70 | 1909–2007 | 0.140 | 0.120 | 0.334 | 41.6 | 26.999 | 0.964 |
| LARIX | 32/64 | 1931–2007 | 0.171 | 0.160 | 0.488 | 52.4 | 40.800 | 0.976 |
| NORTH | 34/68 | 1947–2007 | 0.146 | 0.126 | 0.326 | 38.3 | 19.381 | 0.951 |
| SOUTH | 33/66 | 1909–2007 | 0.133 | 0.119 | 0.319 | 37.5 | 27.386 | 0.965 |
3.2. Similarities and Differences in Radial Growth between Different Species and Topographic Aspects
| LN | LS | PN | PS | PICEA | LARIX | NORTH | SOUTH | |
|---|---|---|---|---|---|---|---|---|
| LN | 79.6 *** | 67.4 ** | 65.3 * | 69.4 ** | 89.8 *** | 75.5 *** | 75.5 *** | |
| LS | 63.3 * | 65.3 * | 65.3 * | 89.8 *** | 71.4 ** | 79.6 *** | ||
| PN | 81.6 *** | 93.9 *** | 65.3 * | 91.8 *** | 79.6 *** | |||
| PS | 87.8 *** | 67.3 ** | 77.6 *** | 85.7 *** | ||||
| PICEA | 67.3 ** | 89.8 *** | 85.7 *** | |||||
| LARIX | 73.5 *** | 81.6 *** | ||||||
| NORTH | 83.7 *** | |||||||
| SOUTH |
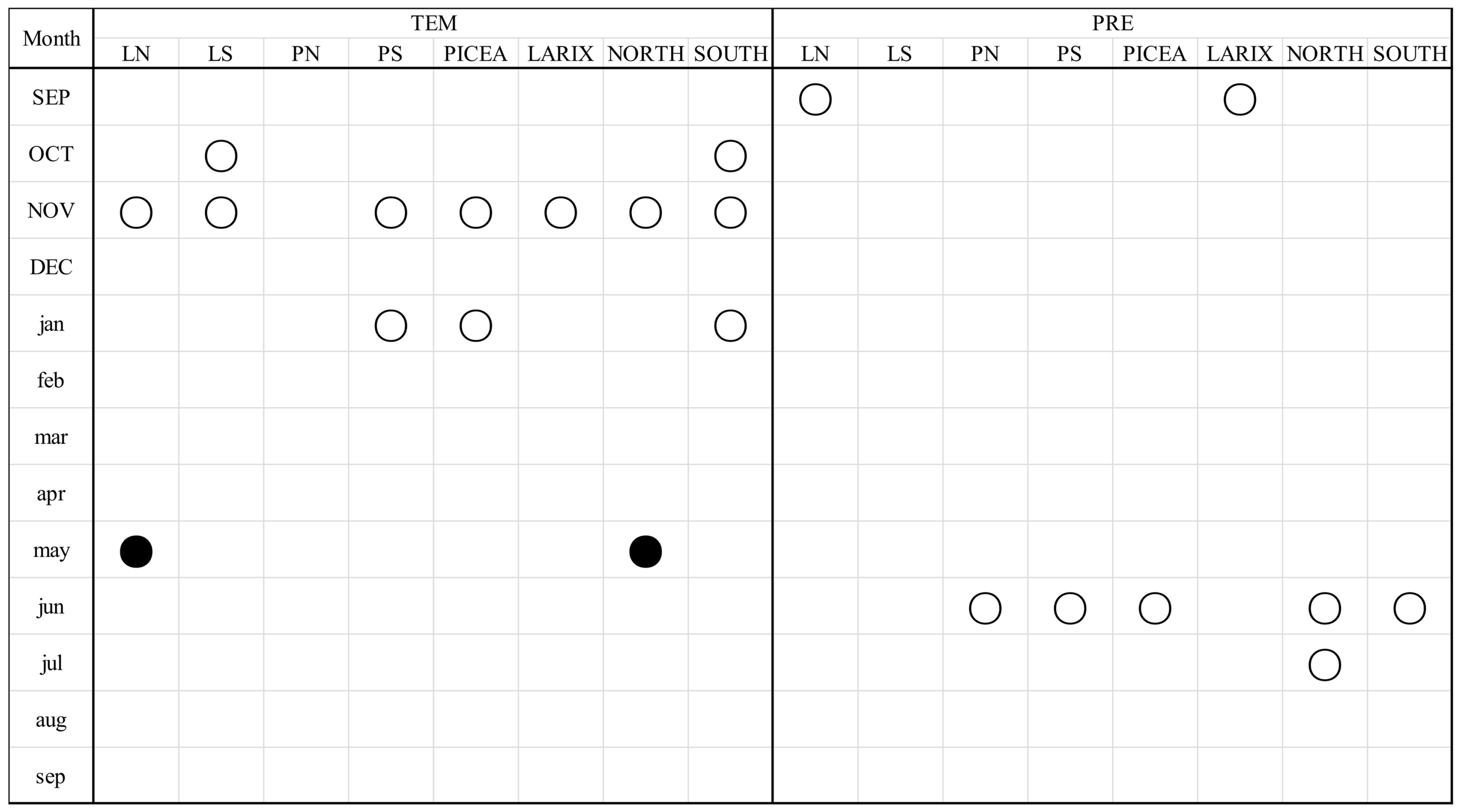
3.3. Correlations between Radial Growth Chronologies and Climatic Variables

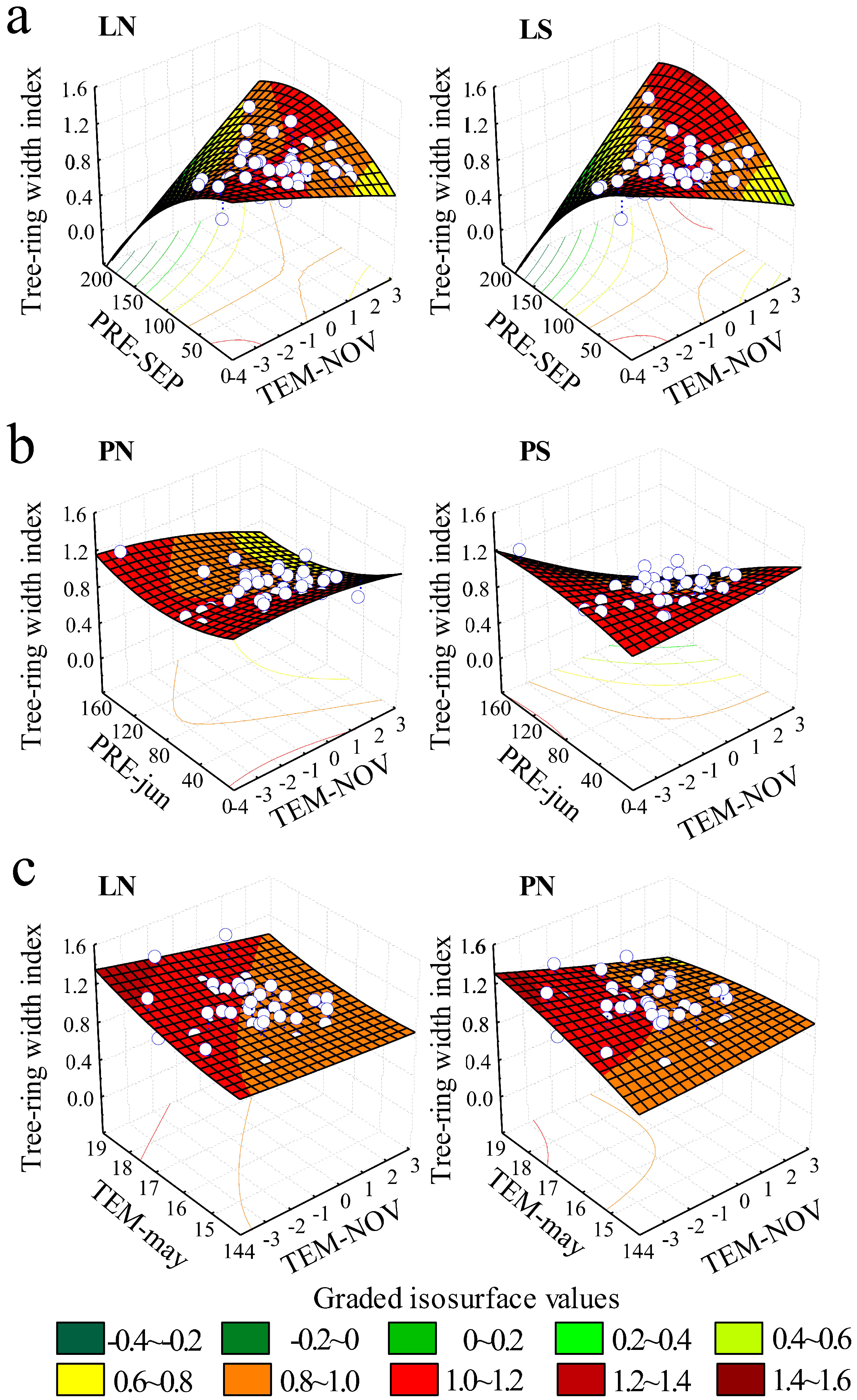
3.4. Variation in the Integrated Influence of Climate Factors on Radial Growth

4. Discussion
4.1. Common Relationships between Growth and Climate at Timberline
4.2. Species-Dependent Growth in Response to Climate
4.3. Topography-Dependent Growth in Response to Climate
4.4. Possible Errors in the Climate and Growth Relationships
4.5. Topography- and Species-Dependent Growth Trends under Climate Change


5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cullen, L.E.; Palmer, J.G.; Duncan, R.P.; Stewart, G.H. Climate change and tree-ring relationships of Nothofagus menziesii tree-line forests. Can. J. For. Res. 2001, 31, 1981–1991. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.; Jiang, Y.; Fang, X.; Wu, S. The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For. Ecol. Manag. 2013, 289, 333–340. [Google Scholar] [CrossRef]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the alpine timberline ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Liu, Y. Climatic response of three tree species growing at different elevations in the Luliang Mountains of Northern China. Dendrochronologia 2013, 31, 311–317. [Google Scholar] [CrossRef]
- Liang, E.; Shao, X.; Eckstein, D.; Huang, L.; Liu, X. Topography- and species-dependent growth responses of Sabina przewalskii and Picea crassifolia to climate on the northeast Tibetan Plateau. For. Ecol. Manag. 2006, 236, 268–277. [Google Scholar] [CrossRef]
- Fekedulegn, D.; Hicks, R.R., Jr.; Colbert, J.J. Influence of topographic aspect, precipitation and drought on radial growth of four major tree species in an Appalachian watershed. For. Ecol. Manag. 2003, 177, 409–425. [Google Scholar] [CrossRef]
- Leonelli, G.; Pelfini, M.; Battipaglia, G.; Cherubini, P. Site-aspect influence on climate sensitivity over time of a high-altitude Pinus cembra tree-ring network. Clim. Chang. 2009, 96, 185–201. [Google Scholar] [CrossRef]
- Elliott, G.P.; Kipfmueller, K.F. Multi-scale influences of slope aspect and spatial pattern on ecotonal dynamics at upper treeline in the southern Rocky Mountains, USA. Arct. Antarct. Alp. Res. 2010, 42, 45–56. [Google Scholar] [CrossRef]
- Fang, J.; Wang, Z.; Tang, Z. Atlas of Woody Plants in China, Distribution and Climate; Higher Education Press and Springer: Beijing, China, 2011. [Google Scholar]
- Liang, E.; Shao, X.; Hu, Y.; Lin, J. Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri) on a sandy substrate in semi-arid grassland, north China. Trees 2001, 15, 230–235. [Google Scholar] [CrossRef]
- Liu, Y.; Cai, Q.; Park, W.; An, Z.; Ma, L. Tree-ring precipitation records from Baiyinaobao, Inner Mongolia since A.D.1838. Chin. Sci. Bull. 2003, 48, 1140–1145. [Google Scholar] [CrossRef]
- Dai, J.; Pan, Y.; Cui, H.; Tang, Z.; Liu, H.; Cao, Y. Impactions of climate change on alpine vegetation on Wutai Mountains. Quat. Sci. 2005, 25, 216–223. [Google Scholar]
- Cai, Q.; Liu, Y. Climatic response of Chinese pine and PDSI variability in the middle Taihang Mountains, north China since 1873. Trees 2013, 27, 419–427. [Google Scholar] [CrossRef]
- He, S.; Xing, Q.; Yin, Z.; Jiang, X. Flora of Beijing; Beijing Press: Beijing, China, 1993. [Google Scholar]
- Ma, Z. Vegetation of Shanxi; China Science and Technology Press: Beijing, China, 2001. [Google Scholar]
- National Soil Survey Office. Annals of Soil Species in China; China Agricultural Press: Beijing, China, 1995. [Google Scholar]
- Wardle, P. An explanation for alpine timberline. New Zealand J. Bot. 1971, 9, 371–402. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R. A time series approach to tree-ring standardization. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Szeicz, J.M.; MacDonald, G.M. Age dependent tree ring growth responses of subartic white spruce to climate. Can. J. For. Res. 1994, 24, 120–132. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Application in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, Holland, 1990. [Google Scholar]
- Fritts, H.C. Reconstruction Large-Scale Climate Patterns from Tree-Ring Data; University of Arizona Press: Tucson, AZ, USA; London, England, 1991. [Google Scholar]
- Fritts, H.C.; Shatz, D.J. Selecting and characterizing the tree-ring chronologies for dendroclimatic analysis. Tree-ring Bull. 1975, 35, 31–46. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology; Kluwer Academic Publishers: Dordrecht, Holland, 1989. [Google Scholar]
- Schweingruber, F.H.; Briffa, K.R.; Nogler, P. A tree-ring densitometric transect from Alaska to Labrador. Int. J. Biometeorol. 1993, 37, 151–169. [Google Scholar] [CrossRef]
- Peterson, D.W.; Peterson, D.L. Effects of climate on radial growth of subalpine conifers in North Casade Mountains. Can. J. For. Res. 1994, 24, 1921–1932. [Google Scholar] [CrossRef]
- Fan, Z.; Bräuning, A.; Cao, K.; Zhu, S. Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern China. For. Ecol. Manag. 2009, 258, 306–313. [Google Scholar] [CrossRef]
- Chen, L.; Wu, S.; Pan, T. Variability of climate-growth relationships along an elevation gradient in the Changbai Mountain, northeastern China. Trees 2011, 25, 1133–1139. [Google Scholar] [CrossRef]
- Takahashi, K.; Tokumitsu, Y.; Yasue, K. Climatic factors affecting the tree-ring width of Betula ermanii at the timberline on Mount. Norikura, central Japan. Ecol. Res. 2005, 20, 445–451. [Google Scholar] [CrossRef]
- Wieser, G.; Tausz, M. Trees at Their Upper Limit: Treelife Limitation at the Alpine Timberline; Springer: Dordrecht, Holland, 2007. [Google Scholar]
- Pallardy, S.G. Physiology of Woody Plants; Academic Press: London, UK, 2008. [Google Scholar]
- Sun, Y.; Wang, L.; Chen, J.; Duan, J.; Shao, X.; Chen, K. Growth characteristics and response to climate change of Larix Miller tree-ring in China. Sci. Chin. Earth Sci. 2010, 53, 871–879. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Dong, M.; Kang, M.; Yang, H. Relationship between the radial growth of Picea meyeri and climate along elevations of the Luyashan Mountain in North-Central China. For. Ecol. Manag. 2012, 265, 142–149. [Google Scholar] [CrossRef]
- Bräuning, A.; Mantwill, B. Summer temperature and summer monsoon history on the Tibetan Plateau during the last 400 years recorded by tree rings. Geophys. Res. Lett. 2004, 31, L24205. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Wang, Y.; Zhou, W.; Ding, H.; Fang, X.; Jiang, S.; Dai, L. Climatic effects on radial growth of major tree species on Changbai Mountain. Ann. For. Sci. 2011, 68, 921–933. [Google Scholar] [CrossRef]
- Yi, L.; Liu, Y.; Song, H.; Li, Q.; Cai, Q.; Yang, Y.; Sun, J. Summer temperature variations since 1676 AD in Luya Mountain, Shanxi province of China, inferred from tree rings. J. Glaciol. Geocryol. 2006, 28, 330–336. [Google Scholar]
- Kimball, K.D.; Keifer, M.B. Climatic comparisons with tree-ring data from montane forests: Are the climatic data appropriate? Can. J. For. Res. 1988, 18, 385–390. [Google Scholar] [CrossRef]
- Liang, E.; Wang, Y.; Eckstein, D.; Luo, T. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytol. 2011, 190, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperature. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Wang, M.; Zhang, L.; Dong, M.; Guo, Y. Responses of radial growth to climate warming in Picea meyeri trees growing at different elevations on the southern slope of Luya Mountain. Chin. J. Plant Ecol. 2013, 37, 1142–1152. [Google Scholar] [CrossRef]
- Harsh, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.P.; Panigrahy, S.; Thapliyal, A.; Kimothi, M.M.; Soni, P.; Parihar, J.S. Monitoring the alpine treeline shift in parts of the Indian Himalayas using remote sensing. Curr. Sci. 2012, 102, 559–562. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Jiang, Y.; Wang, M.; Zhang, L.; Dong, M. Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China. Forests 2015, 6, 116-132. https://doi.org/10.3390/f6010116
Zhang W, Jiang Y, Wang M, Zhang L, Dong M. Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China. Forests. 2015; 6(1):116-132. https://doi.org/10.3390/f6010116
Chicago/Turabian StyleZhang, Wentao, Yuan Jiang, Mingchang Wang, Lingnan Zhang, and Manyu Dong. 2015. "Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China" Forests 6, no. 1: 116-132. https://doi.org/10.3390/f6010116
APA StyleZhang, W., Jiang, Y., Wang, M., Zhang, L., & Dong, M. (2015). Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China. Forests, 6(1), 116-132. https://doi.org/10.3390/f6010116