
Allometric Models Based on Bayesian Frameworks Give Better Estimates of Aboveground Biomass in the Miombo Woodlands



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site

{kind=link}
{kind=link}
{kind=link}
| District | Location | Altitude (m) | Rainfall (mm) * | Rainfall Period | Annual Temperature (°C) | |
|---|---|---|---|---|---|---|
| Minimum | Maximum | |||||
| Kasungu | 12°48′ S, 33°21′ E | 1020–1130 | 800–1000 | November–April | 12 | 24 |
| Salima | 13°40′ S, 34°17′ E | 520–640 | 1000–1200 | October–May | 19.6 | 29.2 |
| Neno | 15°31′ S, 34°41′ E | 320–730 | <800 | November–March | 8 | 32 |
2.2. Selection of Trees for Harvesting
2.3. Tree Measurements and Biomass Sampling
2.4. Sub-Sampling and Drying
| Site | No of Trees | DBH (cm) | Height (m) | Crown Area (m2) | Aboveground Biomass (kg) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Range | Mean | CV% | Range | Mean | CV% | Range | Mean | CV% | Range | Mean | CV% | ||
| Kasungu | 21 | 5.1–55.8 | 18.8 | 68% | 3.3–15.7 | 7.9 | 39% | 0.1–114.7 | 32.4 | 104% | 4.3–1967.6 | 254.0 | 174% |
| Salima | 47 | 6.5–105 | 38.9 | 65% | 4.1–24.2 | 11.1 | 45% | 2.1–397.2 | 100.1 | 95% | 6.2–5357.8 | 806.5 | 143% |
| Neno | 20 | 6.8–74.6 | 29.1 | 61% | 4.2–19.8 | 9.8 | 44% | 3.5–236.8 | 82.0 | 83% | 6–2554.5 | 548.1 | 125% |
2.5. Development and Evaluation of Biomass Models
| Author | Allometric Model | N | DBH | Country | MRE | RMSE | MAPE |
|---|---|---|---|---|---|---|---|
| Chamshama et al. [22] | AGB = 0.0625 × DBH2.553 | 30 | 1–50 | Tanzania | 17 | 937 | 36.4 |
| Chidumayo [8] Model 1 | AGB = 0.0446 × DBH2.765 | 113 | 2–39 | Zambia | 50 | 1491 | 58.3 |
| Chidumayo [8] Model 2 | AGB = 0.0799 × DBH2.5553 | 113 | 2–39 | Zambia | 73 | 2584 | 80.8 |
| Mugasha et al. [23] | AGB = 0.1027 × DBH2.4798 | 167 | 1.1–110 | Tanzania | 50 | 1278 | 56.5 |
| Brown [18] | AGB = 0.1359 × DBH2.2320 | 28 | 5–40 | Global dry forests | 17 | 506 | 32.7 |
| Ryan et al. [21] * | AGB = 0.0265 × DBH2.601 | 29 | 5–73 | Mozambique | 24 | 1169 | 41.6 |
3. Results and Discussion
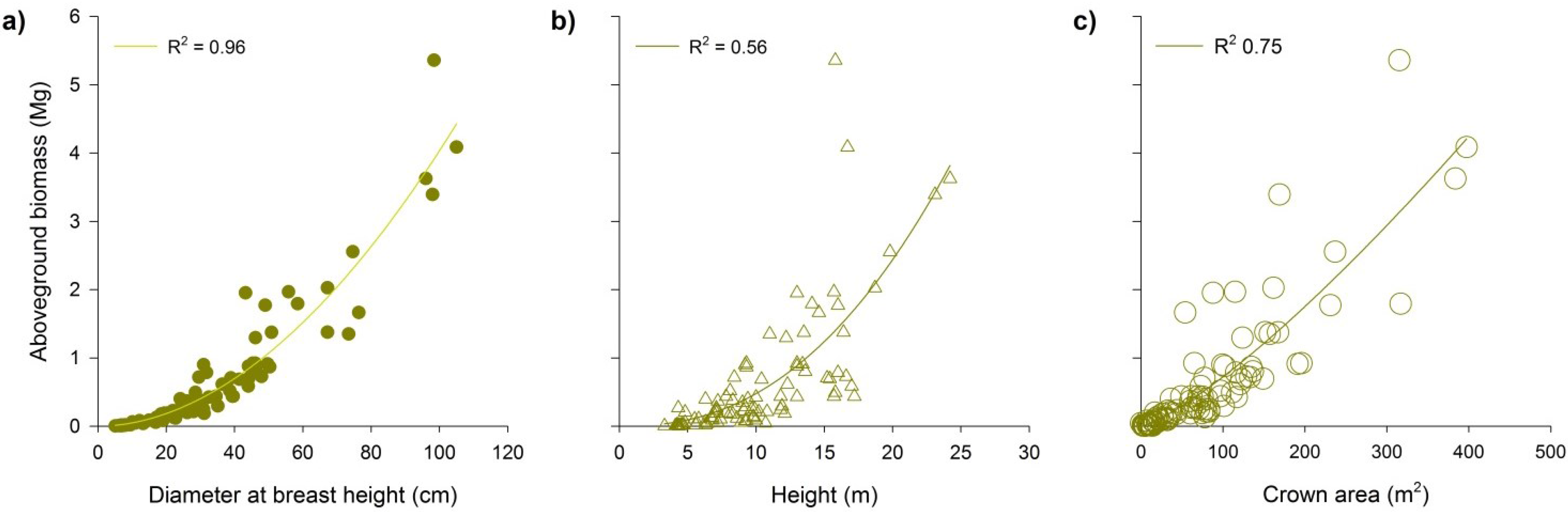
3.1. Relationship between Aboveground Biomass and Predictor Variables

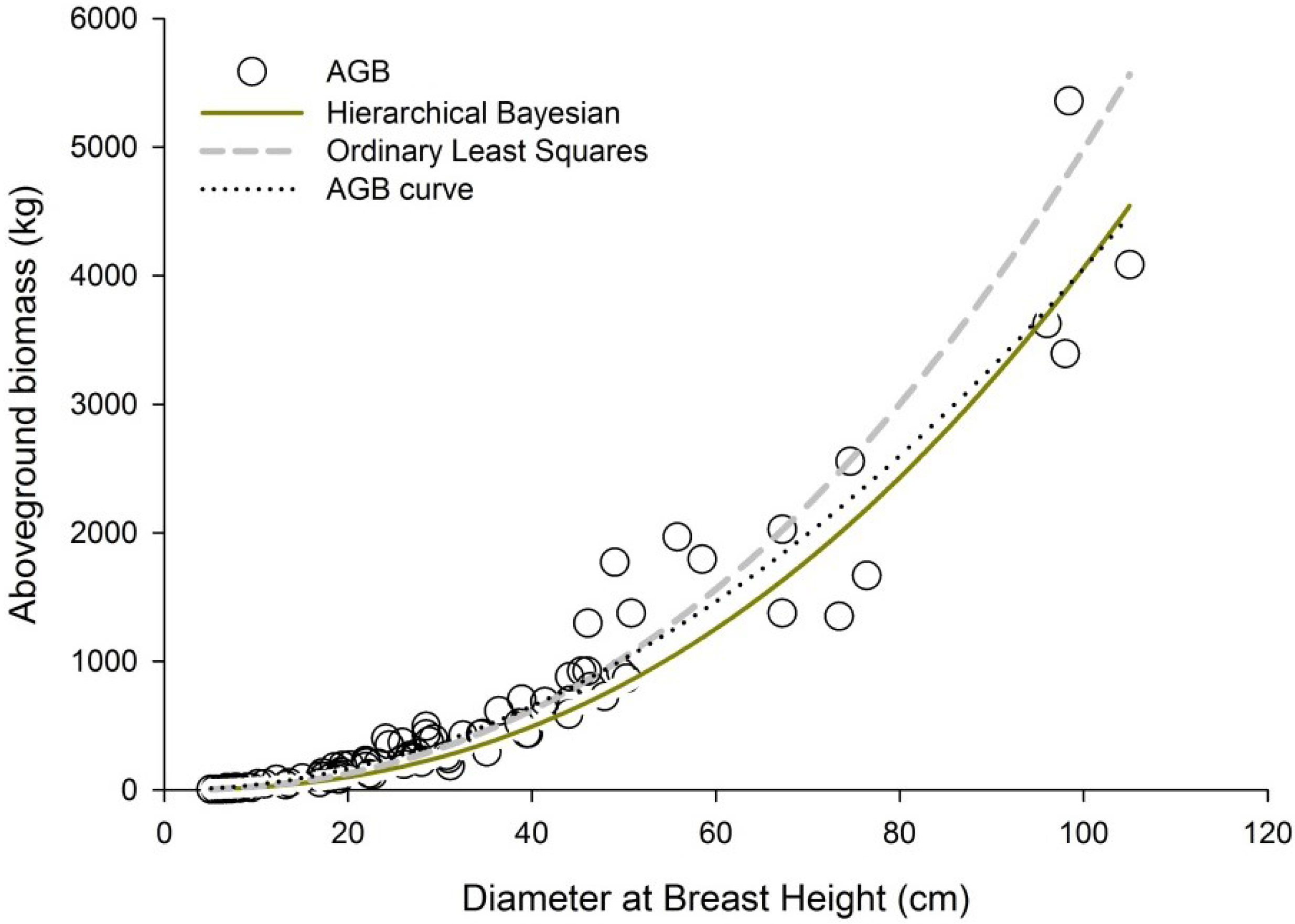
3.2. Performance of New Allometric Models
| Model | Parameter Estimate | DIC | MRE | MAPE | VIF | |||
|---|---|---|---|---|---|---|---|---|
| Intercept (CI) | lnDBH (CI ‡) | lnH (CI) | lnCrA (CI) | |||||
| 1 | −2.284 (−2.635, −1.933) | 2.300 (2.242, 2.438) | 51.04 | −17.34 | 27.20 | |||
| 2 | −1.946 (0.158) | 2.271 (0.048) | 59.02 | 5.34 | 27.98 | None | ||
| 3 | −2.013 (−2.350, −1.683) | 2.187 (2.00, 2.368) | 0.154 (−0.137, 0.452) NS | 59.96 | 5.41 | 27.94 | Acceptable | |
| 4 | −1.838 (−2.180, −0.494) | 2.150 (1.967, 2.334) | 0.0747 (−0.023, 0.173) NS | 58.76 | 5.31 | 27.54 | Acceptable | |
| 5 | −1.910 (−2.255, −1.565) | 2.019 (1.762, 2.276) | 0.203 (−0.084, 0.491) NS | 0.087 (−0.009, 0.183) NS | 58.84 | 5.07 | 27.24 | Moderate |

3.3. Performance of Published Allometric Models

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
| Site | Species Name | DBH (cm) | Height (m) | Crown Area (m2) | Biomass (kg) | |||
|---|---|---|---|---|---|---|---|---|
| Stem | Branch | Twigs | AGB | |||||
| Kasungu | Bauhinia thonningii | 5.1 | 4.3 | 5.11 | 3.41 | 0.45 | 0.45 | 4.31 |
| Zanha africana | 5.6 | 3.3 | 2.27 | 3.18 | 0.96 | 0.58 | 4.73 | |
| Bauhinia thonningii | 6.1 | 4.2 | 3.96 | 3.23 | 0.80 | 0.72 | 4.76 | |
| Antidesma venosum | 7.6 | 4.3 | 7.78 | 11.63 | 4.39 | 0.51 | 16.53 | |
| Bauhinia thonningii | 7.8 | 4.2 | 9.11 | 6.16 | 11.99 | 1.32 | 19.47 | |
| Diplorhynchus condylocarpon | 9.5 | 6.3 | 2.97 | 15.15 | 1.70 | 1.22 | 18.07 | |
| * Brachystegia utilis | 10.1 | 6.3 | 15.22 | 41.25 | 15.59 | 4.30 | 61.15 | |
| Julbernardia paniculata | 10.2 | 7.2 | 16.98 | 25.22 | 16.07 | 8.12 | 49.41 | |
| Brachystegia spiciformis | 10.9 | 10.7 | 0.09 | 34.33 | 8.10 | 3.32 | 45.75 | |
| Diplorhynchus condylocarpon | 12.2 | 9.2 | 6.83 | 22.30 | 40.00 | 16.60 | 78.90 | |
| Brachystegia spiciformis | 13.2 | 6.4 | 10.75 | 25.46 | 2.67 | 10.53 | 38.66 | |
| Brachystegia utilis | 19.1 | 9.8 | 18.73 | 70.64 | 35.62 | 14.65 | 120.91 | |
| S. cauescens | 19.4 | 6.9 | 37.32 | 92.81 | 89.47 | 6.61 | 188.89 | |
| Bauhinia petersiana | 21.9 | 8.1 | 26.41 | 106.67 | 67.18 | 7.65 | 181.50 | |
| Julbernardia paniculata | 21.9 | 9.1 | 32.68 | 134.58 | 73.80 | 16.56 | 224.93 | |
| Bauhinia thonningii | 22.0 | 10.2 | 80.11 | 92.47 | 107.51 | 10.14 | 210.13 | |
| Brachystegia utilis | 24.1 | 9.4 | 36.19 | 122.33 | 237.64 | 39.83 | 399.80 | |
| Bauhinia petersiana | 25.8 | 7.1 | 44.15 | 109.59 | 117.66 | 4.24 | 231.49 | |
| Bobgunnia madagascariensis | 25.9 | 7.2 | 59.97 | 82.58 | 265.23 | 19.69 | 367.50 | |
| Terminalia sericea | 27.5 | 9.2 | 62.67 | 130.40 | 135.51 | 16.66 | 282.58 | |
| * Khaya anthotheca | 30.9 | 9.2 | 99.35 | 314.56 | 556.79 | 27.60 | 898.95 | |
| * Senna siamea | 31.8 | 16.0 | 115.92 | 282.60 | 479.05 | 23.27 | 784.93 | |
| Ficus capensis | 44.1 | 13.0 | 102.10 | 219.14 | 626.66 | 32.66 | 878.47 | |
| Khaya anthotheca | 55.8 | 15.7 | 114.72 | 939.03 | 901.38 | 127.19 | 1967.60 | |
| Neno | Acacia nigrescens | 19.2 | 9.8 | 20.73 | 95.16 | 45.13 | 0.00 | 140.30 |
| Combretum fragrans | 6.8 | 5.6 | 10.15 | 15.50 | 0.89 | 16.39 | ||
| Diplorhynchus condylocarpon | 6.8 | 4.4 | 3.46 | 4.82 | 0.65 | 0.48 | 5.95 | |
| Combretum fragrans | 7.3 | 4.2 | 12.53 | 3.75 | 2.91 | 1.01 | 7.66 | |
| Philenoptera violacea | 8.2 | 4.5 | 9.01 | 7.13 | 3.86 | 2.56 | 13.55 | |
| Anisophyllea boehmii | 17.2 | 6.8 | 77.74 | 57.14 | 66.33 | 7.22 | 130.68 | |
| Combretum fragrans | 18.0 | 6.8 | 22.78 | 56.03 | 25.14 | 6.42 | 87.58 | |
| Combretum fragrans | 18.5 | 8.3 | 42.88 | 72.36 | 94.17 | 11.61 | 178.14 | |
| Anisophyllea boehmii | 20.2 | 7.1 | 74.66 | 65.29 | 90.15 | 35.67 | 191.11 | |
| Terminalia sericea | 26.7 | 7.1 | 65.74 | 117.81 | 127.91 | 19.63 | 265.36 | |
| Piliostigma thonningii | 28.8 | 8.3 | 69.39 | 99.84 | 277.94 | 377.77 | ||
| * Terminalia sericea | 29.5 | 8.4 | 133.24 | 185.02 | 463.52 | 67.92 | 716.46 | |
| Celtis durandii | 34.4 | 13.0 | 75.23 | 271.52 | 158.82 | 430.34 | ||
| Celtis durandii | 34.6 | 11.8 | 96.76 | 203.75 | 233.62 | 437.38 | ||
| Celtis durandii | 35.1 | 11.9 | 101.08 | 211.10 | 83.74 | 294.84 | ||
| Celtis durandii | 38.9 | 15.2 | 77.31 | 451.82 | 253.18 | 705.00 | ||
| Sclerocarya caffra | 41.4 | 10.4 | 122.52 | 275.12 | 345.71 | 64.86 | 685.68 | |
| * Acacia nigrescens | 43.2 | 13.0 | 87.65 | 456.88 | 666.88 | 827.52 | 1951.27 | |
| Sclerocarya caffra | 46.1 | 12.2 | 123.65 | 582.75 | 633.84 | 77.85 | 1294.44 | |
| Sclerocarya caffra | 49.0 | 16.0 | 230.99 | 570.34 | 1144.01 | 55.91 | 1770.26 | |
| Sclerocarya caffra | 50.8 | 13.5 | 167.13 | 485.08 | 833.14 | 56.10 | 1374.31 | |
| Celtis durandii | 74.6 | 19.8 | 236.80 | 1177.87 | 1376.60 | 2554.47 | ||
| * Sterculia africana | 76.0 | 11.9 | 86.56 | 603.34 | 637.29 | 1240.64 | ||
| Salima | Albizia harveyi | 6.5 | 4.1 | 13.95 | 4.13 | 2.04 | 6.16 | |
| Cassia spectabilis | 7.9 | 4.7 | 2.14 | 10.78 | 6.97 | 17.76 | ||
| Philenoptera violacea | 8.2 | 4.3 | 3.96 | 9.17 | 2.18 | 2.01 | 13.36 | |
| Piliostigma thonningii | 9.0 | 4.6 | 11.95 | 10.46 | 10.20 | 20.65 | ||
| Bauhinia spp | 13.6 | 4.8 | 16.24 | 19.94 | 19.73 | 14.51 | 54.17 | |
| Nchezime | 15.0 | 10.0 | 31.65 | 77.22 | 11.95 | 89.17 | ||
| Faidherbia albida | 16.9 | 8.9 | 19.23 | 42.35 | 13.88 | 56.24 | ||
| Combretum fragrans | 17.0 | 5.8 | 33.51 | 39.81 | 54.03 | 15.14 | 108.98 | |
| Combretum fragrans | 17.4 | 7.4 | 30.66 | 63.00 | 22.80 | 18.26 | 104.07 | |
| Faidherbia albida | 19.0 | 9.3 | 14.45 | 51.05 | 29.81 | 80.87 | ||
| Kigelia aethiopica | 19.0 | 5.5 | 20.40 | 52.05 | 32.88 | 19.33 | 104.26 | |
| Piliostigma thonningii | 22.3 | 6.7 | 30.68 | 67.96 | 41.34 | 14.62 | 123.92 | |
| Sclerocarya caffra | 22.7 | 8.6 | 26.64 | 67.96 | 34.07 | 13.90 | 115.93 | |
| Mangifera indica | 23.3 | 4.8 | 35.74 | 39.32 | 135.28 | 31.44 | 206.04 | |
| Bauhinia spp | 24.5 | 9.0 | 72.35 | 91.02 | 200.20 | 57.14 | 348.37 | |
| Philenoptera violacea | 26.1 | 7.8 | 16.82 | 128.96 | 43.68 | 25.54 | 198.17 | |
| Acacia polyacantha | 26.6 | 9.7 | 70.85 | 87.93 | 161.13 | 249.07 | ||
| Acacia polyacantha | 27.3 | 9.9 | 78.04 | 96.18 | 125.21 | 27.41 | 248.81 | |
| Albizia lebbeck | 28.0 | 10.8 | 83.20 | 125.49 | 70.53 | 17.60 | 213.62 | |
| Kigelia pinnata | 28.5 | 7.8 | 49.07 | 152.42 | 242.70 | 39.45 | 434.57 | |
| Nchezime | 28.5 | 15.8 | 111.23 | 289.11 | 182.50 | 22.23 | 493.83 | |
| Bauhinia spp | 29.3 | 6.3 | 80.58 | 125.97 | 219.23 | 51.88 | 397.09 | |
| Piliostigma thonningii | 30.7 | 4.3 | 61.50 | 99.18 | 127.56 | 39.65 | 266.39 | |
| Albizia lebbeck | 30.9 | 11.8 | 83.13 | 161.34 | 65.12 | 16.80 | 243.26 | |
| Faidherbia albida | 31.1 | 12.1 | 60.08 | 110.42 | 61.54 | 13.69 | 185.65 | |
| Acacia polyacantha | 32.5 | 10.0 | 116.77 | 199.62 | 188.98 | 35.04 | 423.64 | |
| Acacia polyacantha | 36.4 | 12.3 | 120.87 | 306.70 | 271.39 | 35.41 | 613.50 | |
| Sclerocarya caffra | 38.6 | 8.1 | 96.70 | 205.85 | 271.08 | 42.03 | 518.96 | |
| Faidherbia albida | 39.4 | 15.7 | 70.12 | 232.24 | 160.90 | 44.08 | 437.22 | |
| Faidherbia albida | 39.6 | 17.2 | 61.43 | 297.39 | 113.73 | 26.14 | 437.25 | |
| Faidherbia albida | 44.0 | 17.0 | 72.98 | 354.88 | 196.06 | 35.39 | 586.33 | |
| Faidherbia albida | 44.1 | 15.4 | 148.94 | 375.38 | 249.36 | 70.69 | 695.43 | |
| Albizia lebbeck | 45.4 | 13.0 | 195.78 | 279.41 | 579.44 | 62.78 | 921.62 | |
| Kigelia aethiopica | 46.0 | 9.3 | 65.05 | 324.12 | 531.16 | 67.24 | 922.52 | |
| Sclerocarya caffra | 46.4 | 13.6 | 136.46 | 578.18 | 156.67 | 66.22 | 801.07 | |
| Faidherbia albida | 47.9 | 16.6 | 128.21 | 389.47 | 260.11 | 77.01 | 726.59 | |
| Albizia lebbeck | 49.7 | 13.4 | 191.01 | 430.27 | 416.73 | 63.53 | 910.53 | |
| Sclerocarya caffra | 50.3 | 9.3 | 134.40 | 271.75 | 501.66 | 93.85 | 867.26 | |
| Acacia polyacantha | 58.5 | 14.1 | 316.86 | 814.47 | 942.17 | 34.80 | 1791.44 | |
| Faidherbia albida | 67.2 | 16.4 | 151.62 | 527.68 | 686.63 | 160.51 | 1374.82 | |
| Faidherbia albida | 67.2 | 18.7 | 161.57 | 790.38 | 1029.40 | 205.53 | 2025.30 | |
| Pterocarpus angolensis | 73.4 | 11.0 | 157.02 | 1309.04 | 40.40 | 1349.44 | ||
| Kigelia aethiopica | 76.4 | 14.6 | 53.98 | 728.05 | 889.06 | 47.48 | 1664.59 | |
| Faidherbia albida | 96.0 | 24.2 | 383.47 | 1989.51 | 1570.40 | 64.22 | 3624.12 | |
| Faidherbia albida | 98.0 | 23.1 | 168.96 | 2028.05 | 1156.51 | 207.61 | 3392.16 | |
| Sclerocarya caffra | 98.4 | 15.8 | 315.26 | 2166.89 | 3008.87 | 182.03 | 5357.80 | |
| Faidherbia albida | 105.0 | 16.7 | 397.22 | 1938.55 | 1986.92 | 159.30 | 4084.76 | |
References
- Campbell, B. The Miombo in Transition: Woodlands and Welfare in Africa; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 1996. [Google Scholar]
- Williams, M.; Ryan, C.M.; Rees, R.M.; Sambane, E.; Fernando, J.; Grace, J. Carbon sequestration and biodiversity of re-growing miombo woodlands in mozambique. For. Ecol. Manag. 2008, 254, 145–155. [Google Scholar] [CrossRef]
- Munishi, P.K.T.; Mringi, S.; Shirima, D.D.; Linda, S.K. The role of the miombo woodlands of the southern highlands of tanzania as carbon sinks. J. Ecol. Nat. Environ. 2010, 2, 261–269. [Google Scholar]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in miombo woodlands of zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making redd a reality. Environ. Res. Lett. 2007, 2, 13. [Google Scholar] [CrossRef]
- Frost, P. The ecology of miombo woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B., Ed.; Center for International Forestry Research (CIFOR): Bongor, Indonesia, 1996; pp. 11–57. [Google Scholar]
- Chidumayo, E.N. Changes in miombo woodland structure under different land tenure and use systems in central zambia. J. Biogeogr. 2012, 29, 1619–1626. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Forest degradation and recovery in a miombo woodland landscape in zambia: 22 years of observations on permanent sample plots. For. Ecol. Manag. 2013, 291, 154–161. [Google Scholar] [CrossRef]
- Harja, D.; Vincent, G.g.; Mulia, R.; van Noordwijk, M. Tree shape plasticity in relation to crown exposure. Trees 2012, 26, 1275–1285. [Google Scholar] [CrossRef]
- Shirima, D.D.; Munishi, P.K.T.; Lewis, S.L.; Burgess, N.D.; Marshall, A.R.; Balmford, A.; Swetnam, R.D.; Zahabu, E.M. Carbon storage, structure and composition of miombo woodlands in tanzania’s eastern arc mountains. Afr. J. Ecol. 2011, 49, 332–342. [Google Scholar] [CrossRef]
- MacFarlane, D.W.; Kuyah, S.; Mulia, R.; Dietz, J.; Muthuri, C.; van Noordwijk, M. Evaluating a non-destructive method for calibrating tree biomass equations derived from tree branching architecture. Trees 2014, 28, 807–817. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; Valentini, R.; Bernoux, M.; Saint-André, L. Estimating tree biomass of sub-saharan african forests: A review of available allometric equations. Silva Fenn. 2011, 45, 477–569. [Google Scholar] [CrossRef]
- Henry, M.; Bombelli, A.; Trotta, C.; Alessandrini, A.; Birigazzi, L.; Sola, G.; Vieilledent, G.; Santenoise, P.; Longuetaud, F.; Valentini, R.; et al. Globallometree: International platform for tree allometric equations to support volume, biomass and carbon assessment. iFor. Biogeosci. For. 2013, 6, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Kuyah, S.; Sileshi, G.W.; Njoloma, J.; Mng’Omba, S.; Neufeldt, H. Estimating aboveground tree biomass in three different miombo woodlands and associated land use systems in malawi. Biomass Bioenergy 2014, 66, 214–222. [Google Scholar] [CrossRef]
- Lowore, J.D.; Abbot, J.I.O.; Werren, M. Stackwood volume estimation for miombo woodlands in malawi. Commonw. For. Rev. 1994, 73, 193–197. [Google Scholar]
- Abbot, J.I.O.; Lowore, J.D.; Werren, M. Models for the estimation of single tree volume in four miombo woodland types. For. Ecol. Manag. 1997, 97, 25–37. [Google Scholar] [CrossRef]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 1997. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.N.; Colgan, M.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2015, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.M.; Williams, M.; Grace, J. Above- and belowground carbon stocks in a miombo woodland landscape of mozambique. Biotropica 2011, 43, 423–432. [Google Scholar] [CrossRef]
- Chamshama, S.A.O.; Mugasha, A.G.; Zahabu, E. Stand biomass and volume estimation for miombo woodlands at kitulangalo, morogoro, tanzania. South. Afr. For. J. 2004, 200, 59–70. [Google Scholar] [CrossRef]
- Mugasha, W.A.; Eid, T.; Bollandsås, O.M.; Malimbwi, R.E.; Chamshama, S.A.O.; Zahabu, E.; Katani, J.Z. Allometric models for prediction of above- and belowground biomass of trees in the miombo woodlands of tanzania. For. Ecol. Manag. 2013, 310, 87–101. [Google Scholar] [CrossRef]
- Vågen, T.-G.; Davey, F.A.; Shepherd, K.D. Land health surveillance: Mapping soil carbon in kenyan rangelands. In Agroforestry—The Future of Global Land Use, Advances in Agroforestry; Nair, P.K.R., Garrity, D., Eds.; Springer Science + Business Media: Dordrecht, The Nederland, 2012; Volume 9, pp. 455–462. [Google Scholar]
- Chaturvedi, R.K.; Raghubanshi, A.S. Aboveground biomass estimation of small diameter woody species of tropical dry forest. New For. 2012, 44, 509–519. [Google Scholar] [CrossRef]
- Preece, N.D.; Crowley, G.M.; Lawes, M.J.; van Oosterzee, P. Comparing above-ground biomass among forest types in the wet tropics: Small stems and plantation types matter in carbon accounting. For. Ecol. Manag. 2012, 264, 228–237. [Google Scholar] [CrossRef]
- Williamson, G.B.; Wiemann, M.C. Measuring wood specific gravity correctly. Am. J. Bot. 2010, 97, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Cuartas, M.; Sierra, C.A.; Alleman, L. Probability distribution of allometric coefficients and bayesian estimation of aboveground tree biomass. For. Ecol. Manag. 2012, 277, 173–179. [Google Scholar] [CrossRef]
- Price, C.A.; Ogle, K.; White, E.P.; Weitz, J.S. Evaluating scaling models in biology using hierarchical bayesian approaches. Ecol. Lett. 2009, 12, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Tredennick, A.T.; Bentley, L.P.; Hanan, N.P. Allometric convergence in savanna trees and implications for the use of plant scaling models in variable ecosystems. PLoS ONE 2013, 8, e58241. [Google Scholar] [CrossRef] [PubMed]
- Abbot, J.I.O.; Homewood, K. A history of change: Causes of miombo woodland decline in a protected area in malawi. J. Appl. Ecol. 1999, 36, 422–433. [Google Scholar] [CrossRef]
- Beauchamp, J.J.; Olson, J.S. Corrrections for bias in regression estimates after logarithmic transformation. Ecology 1973, 54, 1403–1407. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuria, C.; Jamnadassa, R.; Mwangi, P.; Coe, R.; Neufeldt, H. Allometric equations for estimating biomass in agricultural landscapes: I. Aboveground biomass. Agric. Ecosyst. Environ. 2012, 158, 216–224. [Google Scholar] [CrossRef]
- Mate, R.; Johansson, T.; Sitoe, A. Biomass equations for tropical forest tree species in mozambique. Forests 2014, 5, 535–556. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.L.; Baker, T.R. The importance of crown dimensions to improve tropical tree biomass estimates. Ecol. Appl. 2014, 24, 680–698. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.O.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef]
- Bastien-Henri, S.; Park, A.; Ashton, M.; Messier, C. Biomass distribution among tropical trees pecies grown under differing regional climates. For. Ecol. Manag. 2010, 260, 403–410. [Google Scholar] [CrossRef] [Green Version]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuyah, S.; Sileshi, G.W.; Rosenstock, T.S. Allometric Models Based on Bayesian Frameworks Give Better Estimates of Aboveground Biomass in the Miombo Woodlands. Forests 2016, 7, 13. https://doi.org/10.3390/f7020013
Kuyah S, Sileshi GW, Rosenstock TS. Allometric Models Based on Bayesian Frameworks Give Better Estimates of Aboveground Biomass in the Miombo Woodlands. Forests. 2016; 7(2):13. https://doi.org/10.3390/f7020013
Chicago/Turabian StyleKuyah, Shem, Gudeta W. Sileshi, and Todd S. Rosenstock. 2016. "Allometric Models Based on Bayesian Frameworks Give Better Estimates of Aboveground Biomass in the Miombo Woodlands" Forests 7, no. 2: 13. https://doi.org/10.3390/f7020013
APA StyleKuyah, S., Sileshi, G. W., & Rosenstock, T. S. (2016). Allometric Models Based on Bayesian Frameworks Give Better Estimates of Aboveground Biomass in the Miombo Woodlands. Forests, 7(2), 13. https://doi.org/10.3390/f7020013