Stand Dynamics and Biomass Increment in a Lucidophyllous Forest over a 28-Year Period in Central Japan

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
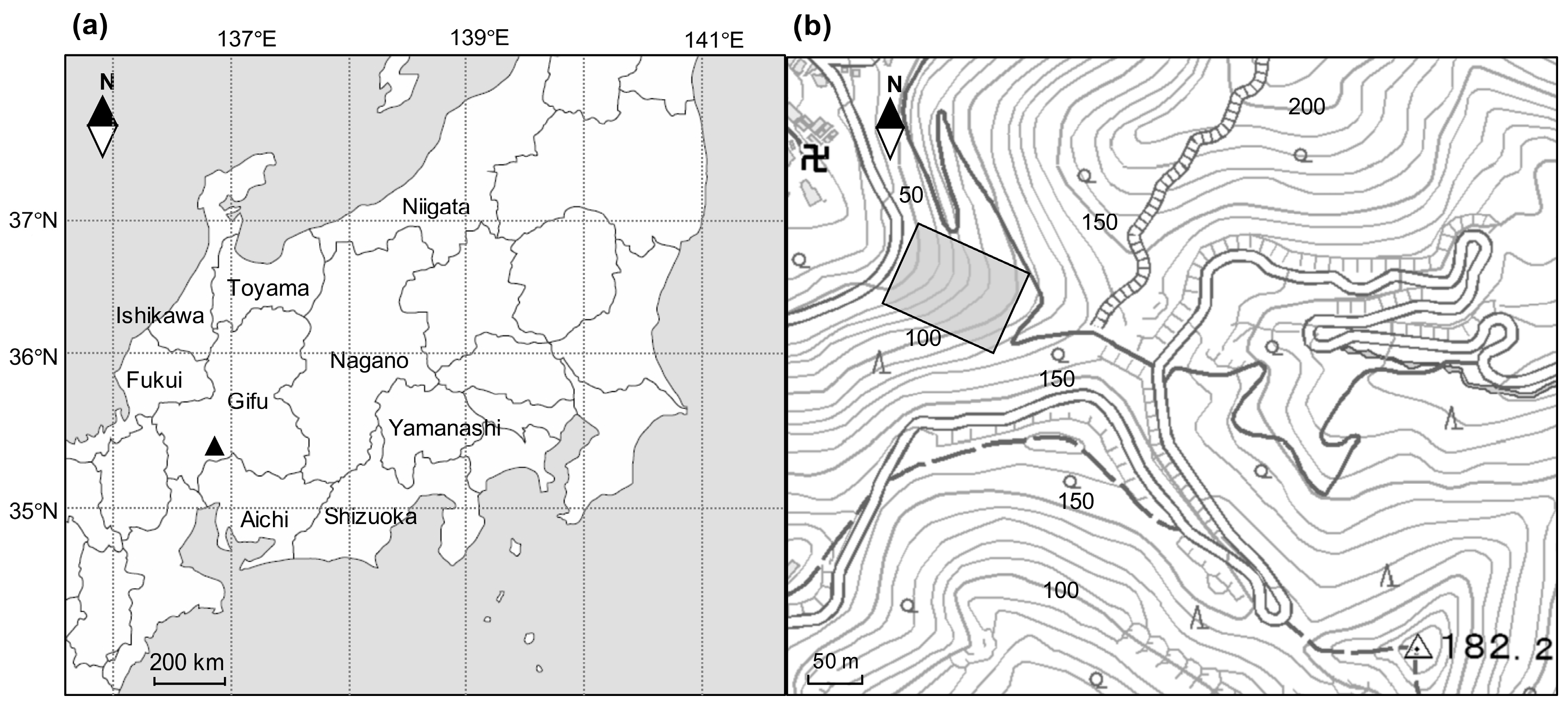
2.1. Study Site
2.2. Field Methods
2.3. Aboveground Biomass Increments and Losses
2.4. Statistical Analysis
3. Results
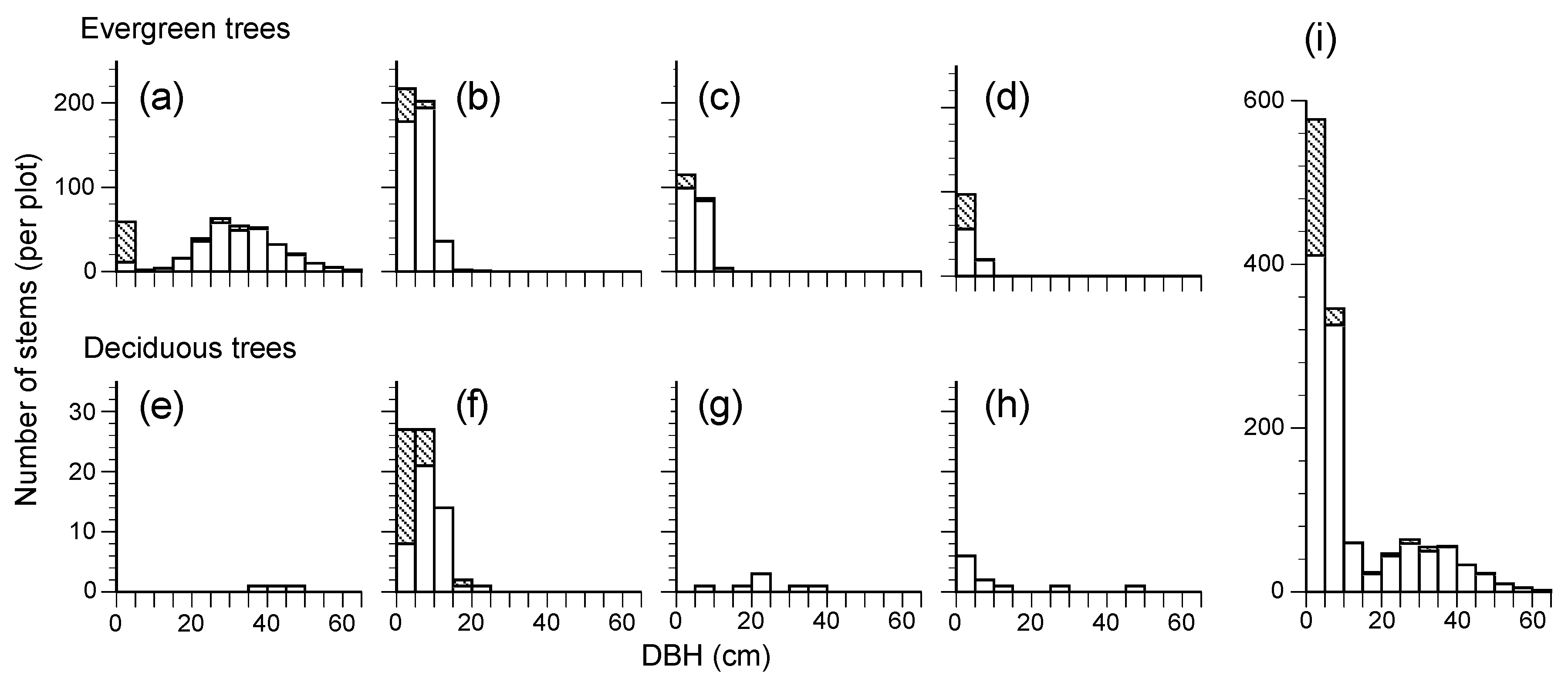
3.1. Structure and Species Composition of a Lucidophyllous Forest of Mt. Kinka
3.2. Temporal Changes of Forest Structure
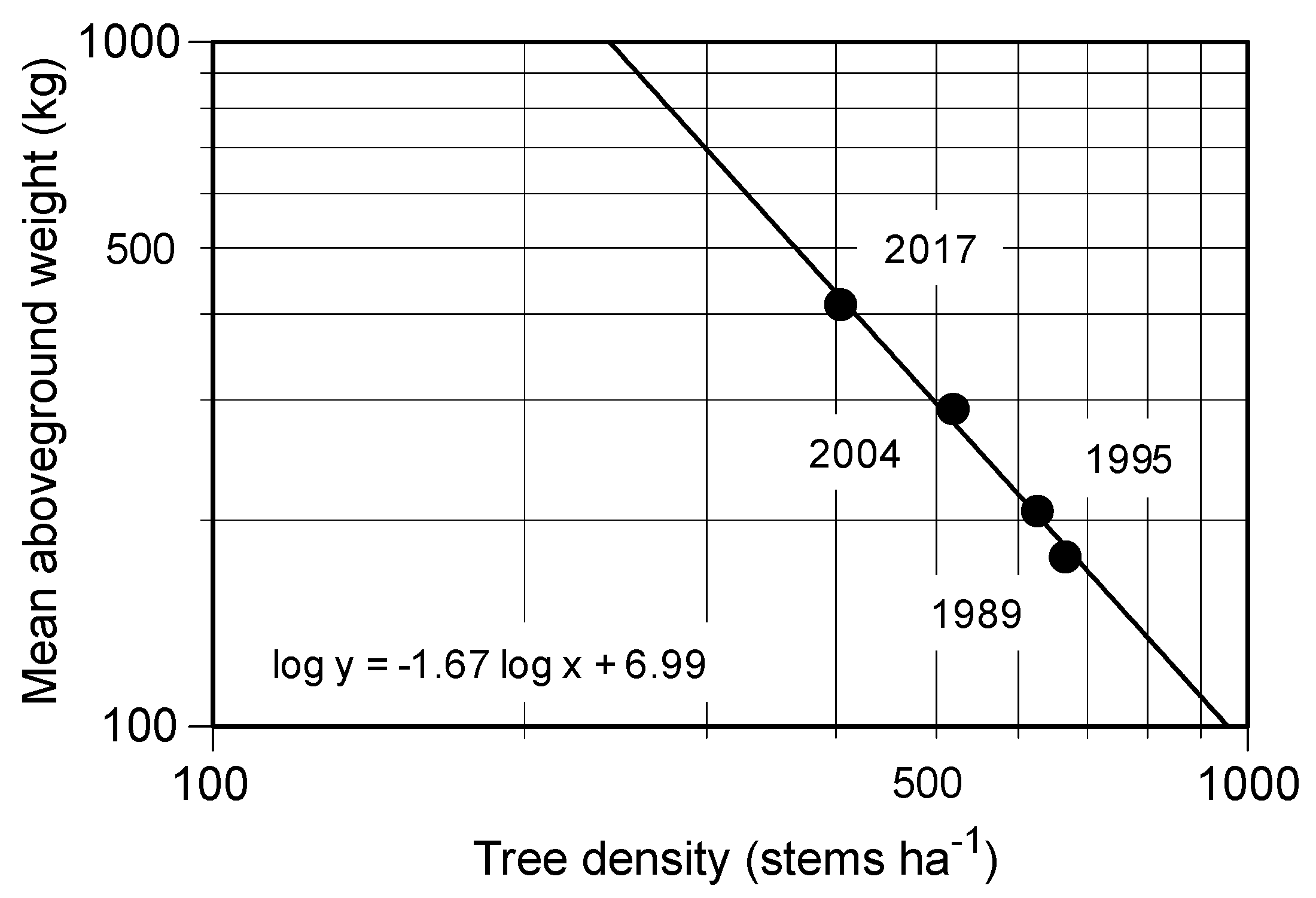
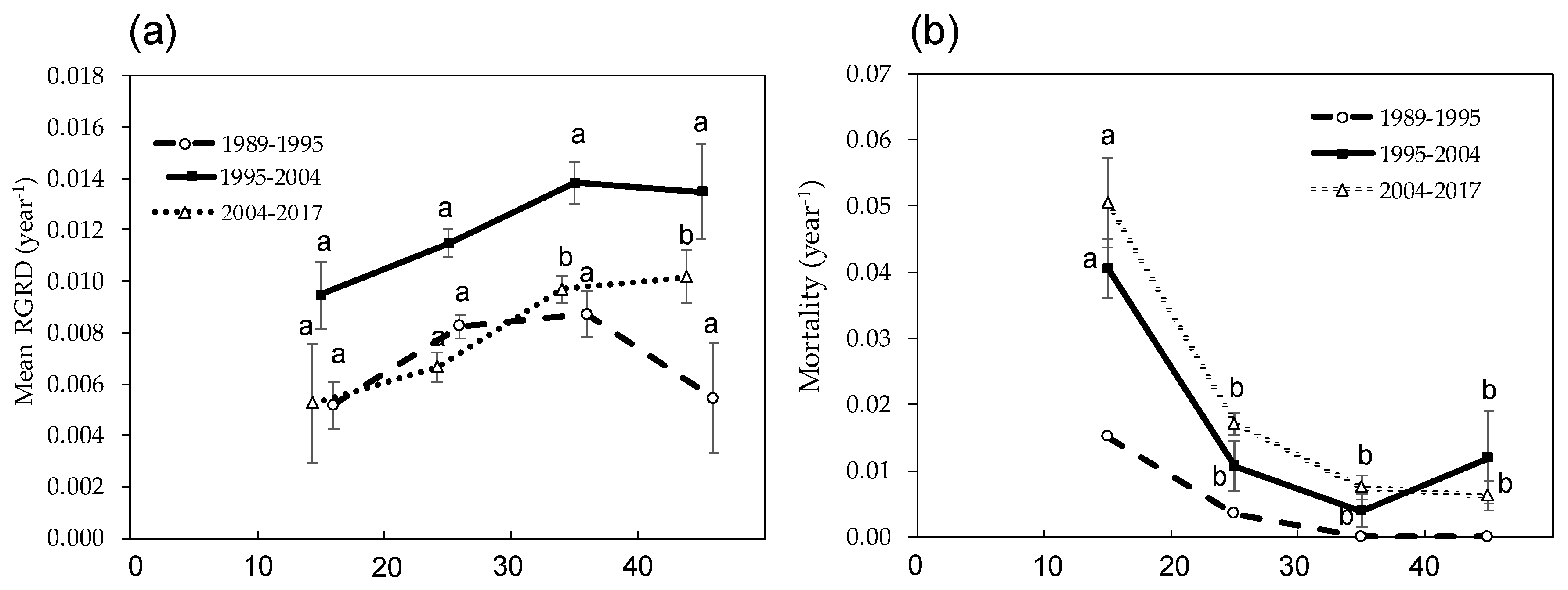
3.3. Change in Forest Biomass and Dynamics
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ohsawa, M. An interpretation of latitudinal patterns of forest limits in south and East Asian mountains. J. Ecol. 1990, 78, 326–339. [Google Scholar] [CrossRef]
- Tagawa, H. Distribution of lucidophll Oak-Laurel forest formation in Asia and other areas. Tropics 1995, 5, 1–40. [Google Scholar] [CrossRef]
- Ohsawa, M. Latitudinal pattern of mountain vegetation zonation in southern and eastern Asia. J. Veg. Sci. 1993, 4, 13–18. [Google Scholar] [CrossRef]
- Tanouchi, H.; Yamamoto, S. Structure and regeneration of canopy species in an old-growth evergreen broad-leaved forest in Aya district, southwestern Japan. Plant Ecol. 1995, 117, 51–60. [Google Scholar] [CrossRef]
- Sato, T.; Kominami, Y.; Saito, S.; Niiyama, K.; Manabe, T.; Tanouchi, H.; Noma, N.; Yamamoto, S. An introduction to the Aya Research Site, a long-term ecological research site, in a warm temperate evergreen broad-leaved forest ecosystems in southwestern Japan: Research topics and design. Bull. Kitakyushu Mus. Nat. His. 1999, 18, 57–180. [Google Scholar]
- Do, T.V.; Sato, T.; Saito, S.; Kozan, O.; Yamagawa, H.; Nagamatsu, D.; Nishimura, N.; Manabe, T. Effects of micro-topographies on stand structure and tree species diversity in an old-growth evergreen broad-leaved forest, southwestern Japan. Glob. Ecol. Conserv. 2015, 4, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Manabe, T.; Nishimura, N.; Miura, M.; Yamamoto, S. Population structure and spatial patterns for trees in a temperate old-growth evergreen broad-leaved forest in Japan. Plant Ecol. 2000, 151, 181–197. [Google Scholar] [CrossRef]
- Manabe, T.; Shimatani, K.; Kawasaki, S.; Aikawa, S.I.; Yamamoto, S.I. The patch mosaic of an old-growth warm-temperate forest: Patch-level descriptions of 40-year gap-forming processes and community structures. Ecol. Res. 2009, 24, 575–586. [Google Scholar] [CrossRef]
- Miura, M.; Manabe, T.; Nishimura, N.; Yamamoto, S. Forest canopy and community dynamics in a temperate old-growth evergreen broad-leaved forest, south-western Japan: A 7-year study of a 4-ha plot. J. Ecol. 2001, 89, 841–849. [Google Scholar] [CrossRef]
- Ishida, H.; Hattori, T.; Takeda, Y. Comparison of species composition and richness between primary and secondary lucidophyllous forests in two altitudinal zones of Tsushima Island, Japan. For. Ecol. Manag. 2005, 213, 273–287. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Shizu, Y.; Nishiwaki, A.; Yashiro, Y.; Koizumi, H. Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest. J. Plant Res. 2010, 123, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Kato, T.; Guo, Z.D.; Yang, Y.H.; Hu, H.F.; Shen, H.H.; Zhao, X.; Kishimoto-Mo, A.W.; Tang, Y.H.; Houghton, R.A. Evidence for environmentally enhanced forest growth. Proc. Natl. Acad. Sci. USA 2014, 111, 9527–9532. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, T.; Mo, W.; Satomura, T.; Inatomi, M.; Koizumi, H. Biometric based carbon flux measurements and net ecosystem production (NEP) in a temperate deciduous broad-leaved forest beneath a flux tower. Ecosystems 2007, 10, 324–334. [Google Scholar] [CrossRef]
- Gower, S.T.; McMurtrie, R.E.; Murty, D. Aboveground net primary production decline with stand age: Potential causes. Trends Ecol. Evol. 1996, 11, 378–382. [Google Scholar] [CrossRef]
- Magnani, F.; Mencuccini, M.; Borghetti, M.; Berbigier, P.; Berninger, F.; Delzon, S.; Grelle, A.; Hari, P.; Jarvis, P.G.; Kolari, P.; et al. The human footprint in the carbon cycle of temperate and boreal forests. Nature 2007, 447, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Aiba, S.; Hill, D.A.; Agetsuma, N. Comparison between old-growth stands and secondary stands regenerating after clear-felling in warm-temperate forests of Yakushima, southern Japan. For. Ecol. Manag. 2001, 140, 163–175. [Google Scholar] [CrossRef]
- Kubota, Y. Population structure and growth dynamics of Castanopsis sieboldii in secondary and old growth subtropical forests in southern Japan. Occas. Pap. 2003, 38, 47–64. [Google Scholar]
- Kubota, Y.; Katsuda, K.; Kikuzawa, K. Secondary succession and effects of clear-logging on diversity in the subtropical forests on Okinawa Island, southern Japan. Biodivers. Conserv. 2005, 14, 879–901. [Google Scholar] [CrossRef]
- Gifu City: Preservation and Management Plan of Gifu Castle Historic Site 2012. Available online: http://www.city.gifu.lg.jp/8881.htm (accessed on 24 April 2014).
- Kawanabe, S. A subtropical broad-leaved forest at Yona, Okinawa. In Primary Productivity of Japanese Forests—Productivity of Terrestrial Communities; Shidei, T., Kira, T., Eds.; University of Tokyo Press: Tokyo, Japan, 1977; pp. 268–279. [Google Scholar]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Saigusa, N.; Iimura, Y.; Muraoka, H.; Koizumi, H. Biometric-Based Estimations of Net Primary Production (NPP) in Forest Ecosystems. In Canopy Photosynthesis: From Basics to Applications Volume 42 of the Series Advances in Photosynthesis and Respiration; Hikosaka, K., Niinemets, Ü., Anten, N., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 333–351. [Google Scholar]
- Yoda, K.; Kira, T.; Ogawa, F.; Hozumi, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants. XI). J. Biol. Osaka City Univ. 1963, 14, 107–129. [Google Scholar]
- Ohtsuka, T.; Saigusa, N.; Koizumi, H. On linking multiyear biometric measurements of tree growth with eddy covariance-based net ecosystem production. Glob. Chang. Biol. 2009, 15, 1015–1024. [Google Scholar] [CrossRef]
- Kira, T.; Yabuki, K. Primary production rate in the Minamata forest. In Biological production in a warm-Temperate Evergreen Oak Forest of Japan; Kira, T., Ono, Y., Hosokawa, T., Eds.; JIBP Synthesis., 18; University of Tokyo Press: Tokyo, Japan, 1978; pp. 131–138. [Google Scholar]
- Tadaki, Y. Studies on the production structure of forest (XIV) The third report on the primary production of a young stand of Castanopsis cuspidata. Jpn. J. For. Res. 1968, 50, 60–65. [Google Scholar]
- Tadaki, Y. Studies on the production structure of forest (VII) The primary production of a young stand of Castanopsis cuspidata. Jpn. J. Ecol. 1965, 15, 142–147. [Google Scholar]
- Dang, C.L.; Wu, Z.L. Studies on the net primary production for Castanopsis echidnocarpa community of monsoon evergreen broad-leaved forest. J. Yunnan Univ. 1992, 14, 108–117. [Google Scholar]
- Ryan, M.G.; Binkley, D.; Fownes, J.H. Age-related decline in forest productivity: Pattern and process. Adv. Ecol. Res. 1997, 27, 213–262. [Google Scholar]
- Berger, U.; Hildenbrandt, H.; Grimm, V. Age-related decline in forest production: Modelling the effects of growth limitation, neighbourhood competition and self-thinning. J. Ecol. 2004, 92, 846–853. [Google Scholar] [CrossRef]
- Sato, T.; Kominami, Y.; Saito, S.; Niiyama, K. Spatial variations of net primary production in an old-growth lucidophyllous forest, Aya, southwestern Japan. In Proceedings of the Japanese Forestry Society Conference, Morioka, Japan, 31 March 2003. [Google Scholar]
- Do, T.V.; Sato, T.; Saito, S.; Kozan, O. Fine-root production and litterfall: Main contributions to net primary production in an old-growth evergreen broad-leaved forest in southwestern Japan. Ecol. Res. 2015, 30, 921–930. [Google Scholar]
- Sato, T.; Kominami, Y.; Saito, S.; Niiyama, K.; Tanouchi, H.; Nagamatsu, D.; Nomiya, H. Temporal dynamics and resilience of fine litterfall in relation to typhoon disturbances over 14 years in an old-growth lucidophyllous forest in southwestern Japan. Plant Ecol. 2010, 208, 187–198. [Google Scholar] [CrossRef]
- Kimura, M. Primary production of the warm-temperate laurel forest in the southern part of Osumi peninsula, Kyushu, Japan. Misc. Rep. Res. Inst. Nat. Resour. 1960, 52–53, 36–47. [Google Scholar]
- Yang, T.H.; Song, K.; Da, L.J.; Li, X.P.; Wu, J.P. The biomass and aboveground net primary productivity of Schima superba—Castanopsis carlesii forests in east China. Sci. China Life Sci. 2010, 53, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.M.; Lin, P.; Li, Z.J.; He, J.Y.; Liu, C.D. Biomass and productivity of Castanopsis eyrei community in Wuyi Mountains. J. Xiamen Univ. (Nat. Sci.) 1996, 35, 269–275. [Google Scholar]
- Dang, C.L.; Wu, Z.L. Studies on the net primary production of Castanopsis orthacantha community. J. Yunnan Univ. 1994, 16, 200–204. [Google Scholar]
- Dang, C.L.; Wu, Z.L. Studies on the biomass of Castanopsis orthacantha community. J. Yunnan Univ. 1994, 16, 195–199. [Google Scholar]
- Dang, C.L.; Wu, Z.L.; Zhang, Z. Studies on the biomass of Cyclobalanopsis delavayi community. J. Yunnan Univ. 1994, 16, 205–209. [Google Scholar]
- Dang, C.L.; Wu, Z.L.; Zhang, Z. Studies on the net primary production of Cyclobalanopsis delavayi community. J. Yunnan Univ. 1994, 16, 210–219. [Google Scholar]
- Dang, C.L.; Wu, Z.L. Studies on the biomass for Castanopsis echidnocarpa Community of monsoon evergreen broad-leaved forest. J. Yunnan Univ. 1992, 14, 95–107. [Google Scholar]
- Zhang, L.; Luo, T.X.; Deng, K.M.; Dai, Q.; Huang, Y.; Jiang, Z.F.; Tao, M.Y.; Zeng, K.Y. Biomass and net primary productivity of secondary evergreen broadleaved forest in Huangmian Forest Farm, Guangxi. Chin. J. Appl. Ecol. 2004, 15, 2029–2033. [Google Scholar]
- Xie, S.C.; Liu, W.Y.; Li, S.C.; Yang, G.P. Preliminary studies on the biomass of middle-mountain moist evergreen broadleaved forests in Ailao Mountain, Yunnan. Acta Phytoecol. Sin. 1996, 20, 167–176. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
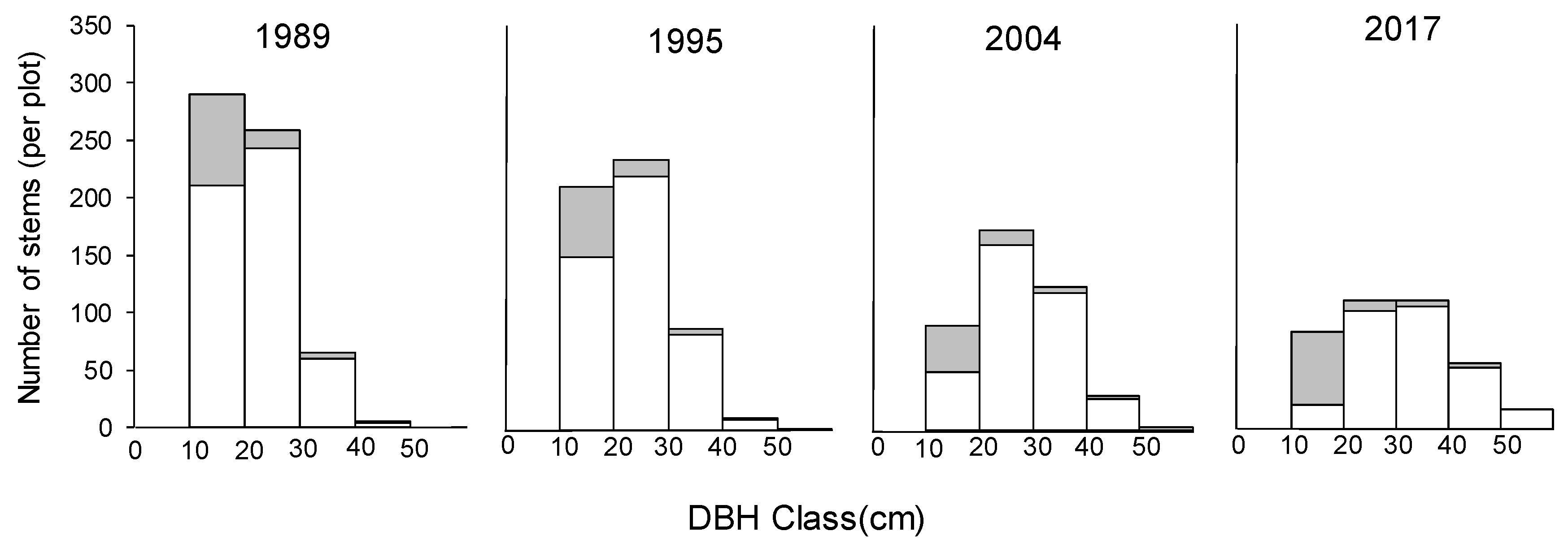{kind=link}
| Species | Basal Area | DBH (cm) | No. of Stems | |||
|---|---|---|---|---|---|---|
| cm2 ha–1 | % | Mean | Maximum | Per Plot | % | |
| Evergreen trees | ||||||
| Castanopsis cuspidata | 404,424 | 87.76 | 28.0 | 63.9 | 359 | 27.6 |
| Cleyera japonica | 20,592 | 4.47 | 5.6 | 23.0 | 458 | 35.2 |
| Eurya japonica | 6580 | 1.43 | 5.0 | 12.6 | 206 | 15.8 |
| Quercus glauca | 3321 | 0.72 | 3.3 | 37.4 | 118 | 9.1 |
| Ilex rotunda | 350 | 0.08 | 3.2 | 9.7 | 23 | 1.8 |
| Prunus spinulosa | 78 | 0.02 | 3.0 | 4.8 | 6 | 0.5 |
| Illicium anisatum | 56 | 0.01 | 4.9 | 6.1 | 2 | 0.2 |
| Aucuba japonica | 54 | 0.01 | 1.9 | 3.7 | 10 | 0.8 |
| Ilex latifolia | 18 | < 0.01 | 2.2 | 3.0 | 3 | 0.2 |
| Cinnamomum tenuifolium | 16 | < 0.01 | 3.8 | 3.8 | 1 | 0.08 |
| Gardenia jasminoides | 13 | < 0.01 | 1.9 | 2.2 | 3 | 0.2 |
| Photinia glabra | 6 | < 0.01 | 1.3 | 1.8 | 3 | 0.2 |
| Ligustrum japonicum | 5 | < 0.01 | 2.2 | 2.2 | 1 | 0.08 |
| Subtotal | 435,514 | 94.51 | - | - | 1193 | 91.7 |
| Deciduous trees | ||||||
| Ilex micrococca | 6213 | 1.35 | 42.7 | 48.4 | 3 | 0.2 |
| Ilex macropoda | 5974 | 1.30 | 7.2 | 25.0 | 71 | 5.5 |
| Magnolia obovata | 4716 | 1.02 | 22.9 | 38.7 | 7 | 0.5 |
| Eleutherococcus sciadophylloides | 3585 | 0.78 | 10.2 | 46.2 | 11 | 0.8 |
| Rhus sylvestris | 2283 | 0.50 | 15.8 | 22.6 | 7 | 0.5 |
| Quercus serrata | 1618 | 0.35 | 38.0 | 38.0 | 1 | 0.08 |
| Padus grayana | 696 | 0.15 | 24.9 | 24.9 | 1 | 0.08 |
| Clethra barbinervis | 95 | 0.02 | 5.5 | 9.0 | 2 | 0.2 |
| Diospyros kaki | 45 | 0.01 | 6.3 | 6.3 | 1 | 0.08 |
| Hamamelis japonica | 25 | 0.01 | 4.7 | 4.7 | 1 | 0.08 |
| Carpinus laxiflora | 22 | < 0.01 | 4.4 | 4.4 | 1 | 0.08 |
| Aphananthe aspera | 9 | < 0.01 | 2.9 | 2.9 | 1 | 0.08 |
| Styrax japonica | 8 | < 0.01 | 2.6 | 2.6 | 1 | 0.08 |
| Subtotal | 25,290 | 5.49 | - | - | 108 | 8.3 |
| Total | 460,804 | 100 | 11.7 | 63.9 | 1301 | 100 |
| Species | Number of Stems (ha–1) | BA (m2 ha–1) | ||||||
|---|---|---|---|---|---|---|---|---|
| 1989 (May) | 1995 (May) | 2004 (October) | 2017 (January) | 1989 (May) | 1995 (May) | 2004 (October) | 2017 (January) | |
| Evergreen trees | ||||||||
| Castanopsis cuspidata | 666 ± 13 a | 626 ± 13 a | 519 ± 14 b | 404 ± 10 c | 29.18 ± 1.84 a | 31.42 ± 2.46 ab | 35.70 ± 2.06 ab | 38.71 ± 2.22 b |
| Other species | 19 ± 5 a | 30 ± 6 a | 20 ± 5 a | 56 ± 7 b | 0.33 ± 0.10 a | 0.40 ± 0.11 ab | 0.43 ± 0.10 ab | 0.84 ± 0.14 b |
| Subtotal | 684 ± 15 a | 656 ± 18 a | 539 ± 11 b | 460 ± 7 c | 29.51 ± 1.80 a | 31.81 ± 2.41 a | 36.13 ± 2.03 b | 39.54 ± 2.24 c |
| Deciduous trees | 91 ± 30 a | 79 ± 34 a | 56 ± 19 a | 46 ± 14 a | 2.76 ± 0.85 a | 2.90 ± 0.94 a | 2.45 ± 0.59 a | 2.21 ± 0.77 a |
| Total | 779 ± 37 a | 733 ± 38 a | 591 ± 21 b | 510 ± 16 b | 33.23 ± 1.88 a | 34.99 ± 2.04 ab | 38.31 ± 1.83 ab | 42.13 ± 1.82 b |
| Species | 1989 (May) | 1995 (May) | 2004 (October) | 2017 (January) | |
|---|---|---|---|---|---|
| Evergreen species | |||||
| 1 | Castanopsis cuspidata | 518 | 463 | 361 | 298 |
| 2 | Cleyera japonica | 12 | 14 | 15 | 39 |
| 3 | Quercus glauca | 2 | 2 | 2 | 1 |
| 4 | Eurya japonica | 1 | 1 | 1 | 4 |
| 5 | Ilex pedunculosa | 1 | 1 | 0 | 0 |
| Subtotal | 534 | 481 | 379 | 342 | |
| Deciduous species | |||||
| 1 | Magnolia obovata | 15 | 13 | 10 | 6 |
| 2 | Eleutherococcus sciadophylloides | 24 | 14 | 5 | 3 |
| 3 | Ilex micrococca | 7 | 7 | 5 | 3 |
| 4 | Ilex macropoda | 16 | 16 | 16 | 17 |
| 5 | Rhus sylvestris | 14 | 9 | 6 | 6 |
| 6 | Quercus serrata | 2 | 2 | 1 | 1 |
| 7 | Padus grayana | 2 | 1 | 1 | 1 |
| 8 | Cerasus jamasakura | 1 | 1 | 0 | 0 |
| 9 | Hamamelis japonica | 1 | 1 | 0 | 0 |
| 10 | Gamblea innovans | 1 | 0 | 0 | 0 |
| 11 | Carpinus laxiflora | 1 | 0 | 0 | 0 |
| 12 | Styrax japonica | 1 | 0 | 0 | 0 |
| Subtotal | 85 | 64 | 44 | 37 | |
| Total | 619 | 545 | 423 | 379 | |
| Number of species | 17 | 14 | 11 | 11 | |
| Parts of Biomass | Period | Castanopsis cuspidata | Other Tree Species | All Trees | |||
|---|---|---|---|---|---|---|---|
| SI | D | SI | D | SI | D | ||
| Aboveground | 1989–1995 | 2.30 ± 0.18 a | 1.09 * | 0.22 ± 0.06 a | 0.28 * | 2.53 ± 0.17 a | 1.37 * |
| 1995–2004 | 3.84 ± 0.38 b | 1.59 ± 0.43 ab | 0.21 ± 0.05 a | 0.32 ± 0.17 a | 4.05 ± 0.34 b | 1.91 ± 0.39 b | |
| 2004–2017 | 3.15 ± 0.21 ab | 1.74 ± 0.21 b | 0.20 ± 0.06 a | 0.27 ± 0.03 a | 3.35 ± 0.17 ab | 2.01 ± 0.20 b | |
| Woody parts (stems & branches) | 1989–1995 | 2.23 ± 0.18 a | 1.05 * | 0.17 ± 0.06 a | 0.26 * | 2.40 ± 0.13 a | 1.31 * |
| 1995–2004 | 3.73 ± 0.37 b | 1.53 ± 0.41 a | 0.20 ± 0.04 a | 0.30 ± 0.16 a | 3.93 ± 0.33 b | 1.83 ± 0.37 | |
| 2004–2017 | 3.06 ± 0.21 ab | 1.67 ± 0.21 a | 0.20 ± 0.06 a | 0.26 ± 0.02 a | 3.26 ± 0.16 ab | 1.93 ± 0.19 b | |
| Year | 10–20 cm | 20–30 cm | 30–40 cm | ≥40 cm | Total |
|---|---|---|---|---|---|
| 1989 | 23.28 ± 2.59 a | 68.61 ± 2.40 a | 33.56 ± 7.96 a | 6.25 ± 2.64 a | 131.70 ± 7.17 a |
| 1995 | 19.34 ± 2.40 a | 67.18 ± 3.26 a | 43.03 ± 6.74 ab | 13.14 ± 4.46 a | 142.67 ± 8.89 a |
| 2004 | 8.79 ± 2.09 b | 49.26 ± 1.02 b | 69.94 ± 3.95 b | 34.85 ± 8.40 a | 162.83 ± 8.32 ab |
| 2017 | 6.05 ± 1.50 b | 31.60 ± 3.82 c | 63.16 ± 7.13 b | 79.83 ± 13.23 b | 180.64 ± 8.68 b |
| Latitude and Longtitude | Study Site | Dominant Species | No. of Stems (ha–1) | Aboveground Biomass (ton ha–1 ) | Woody (Stem and Branch) NPP (ton ha–1 year–1) | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Leaf | Stem and Branch | Total | ||||||
| 35°26′ N 136°47′ E | Mt. Kinka, Gifu, Japan | Castanopsis cuspidata | 884 | 5.4 | 133.3 | 138.7 | - | This study |
| 779 | 5.6 | 139.9 | 145.5 | 2.2 | ||||
| 604 | 5.9 | 156.3 | 162.2 | 3.6 | ||||
| 541 | 6.6 | 180.3 | 186.9 | 3.1 | ||||
| 32°47′ N 130°44′ E | Kumamoto City, Japan | Castanopsis cuspidata | 24,667 | 8.4 | 72.1 | 80.5 | 14.1 | [28] |
| 32°12′ N 130°24′ E | Minamata City, Japan | Castanopsis cuspidata | - | 7.8 | 335.3 | 343.1 | 4.6 | [27] |
| - | 7.8 | 369.8 | 377.6 | 3.4 | ||||
| - | 6.8 | 346.7 | 353.5 | 4.5 | ||||
| 32°04′ N 131°09′ E | Aya Research site, Japan | Distylium racemosum; Persea thunbergii; Quercus acuta, salicina, and gilva | 1391 | - | - | 446.4 | 4.3 | [33,34,35] |
| 31°12′ N 130°55′ E | Mt. Aranishi, Japan | Distylium racemosum | - | 11.4 | 312.0 | 323.4 | 7.4 | [36] |
| 29°48′ N 121°47′ E | Tiantong National Forest Park, China | Schima superba; Castanopsis carlesii | - | 4.9 | 157.4 | 162.3 | 3.5 | [37] |
| 27°42′ N 117°41′ E | Wuyi Mountain, China | Castanopsis eyrei | 1140 | 12.9 | 335.1 | 348.0 | 8.8 | [38] |
| 26°45′ N 122°13′ E | Yona, Okinawa Island, Japan | C. cuspidata var. sieboldii | 2900 | 7.7 | 185.6 | 193.3 | 11.4 | [21] |
| 25°24′ N 102°45′ E | Songming, China | Castanopsis orthacantha | 3910 | 7.8 | 192.5 | 200.4 | 11.3 | [39,40] |
| 25°19′ N 103°33′ E | Fuming, China | Cyclobalanopsis delavayi | - | 6.1 | 86.7 | 92.8 | 8.4 | [41,42] |
| 25°19′ N 103°33′ E | Puer, China | Castanopsis echidnocarpa | - | 7.0 | 60.1 | 67.1 | 11.8 | [31,43] |
| - | 6.9 | 124.2 | 131.1 | 11.6 | ||||
| 24°51′ N 109°51′ E | Huangmian, China | Schima superba; Engelhardtia chrysolepis; Castanopsis fissa; Cyclobalanopsis glaucoides | 2050 | 4.9 | 64.2 | 69.1 | 7.5 | [44] |
| 24°10′ N 101°25′ E | Ailao Mountain, China | Lithocarpus xylocarpus; L. chintungensis and Castanopsis wattii | 1100 | 7.9 | 343.6 | 351.5 | 6.6 | [45] |
| 512 | 7.3 | 201.4 | 208.7 | 4.3 | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Komiyama, A.; Kato, S.; Cao, R.; Yoshitake, S.; Ohtsuka, T. Stand Dynamics and Biomass Increment in a Lucidophyllous Forest over a 28-Year Period in Central Japan. Forests 2017, 8, 397. https://doi.org/10.3390/f8100397
Chen S, Komiyama A, Kato S, Cao R, Yoshitake S, Ohtsuka T. Stand Dynamics and Biomass Increment in a Lucidophyllous Forest over a 28-Year Period in Central Japan. Forests. 2017; 8(10):397. https://doi.org/10.3390/f8100397
Chicago/Turabian StyleChen, Siyu, Akira Komiyama, Shogo Kato, Ruoming Cao, Shinpei Yoshitake, and Toshiyuki Ohtsuka. 2017. "Stand Dynamics and Biomass Increment in a Lucidophyllous Forest over a 28-Year Period in Central Japan" Forests 8, no. 10: 397. https://doi.org/10.3390/f8100397
APA StyleChen, S., Komiyama, A., Kato, S., Cao, R., Yoshitake, S., & Ohtsuka, T. (2017). Stand Dynamics and Biomass Increment in a Lucidophyllous Forest over a 28-Year Period in Central Japan. Forests, 8(10), 397. https://doi.org/10.3390/f8100397