Elevational Shifts in the Topographic Position of Polylepis Forest Stands in the Andes of Southern Peru

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
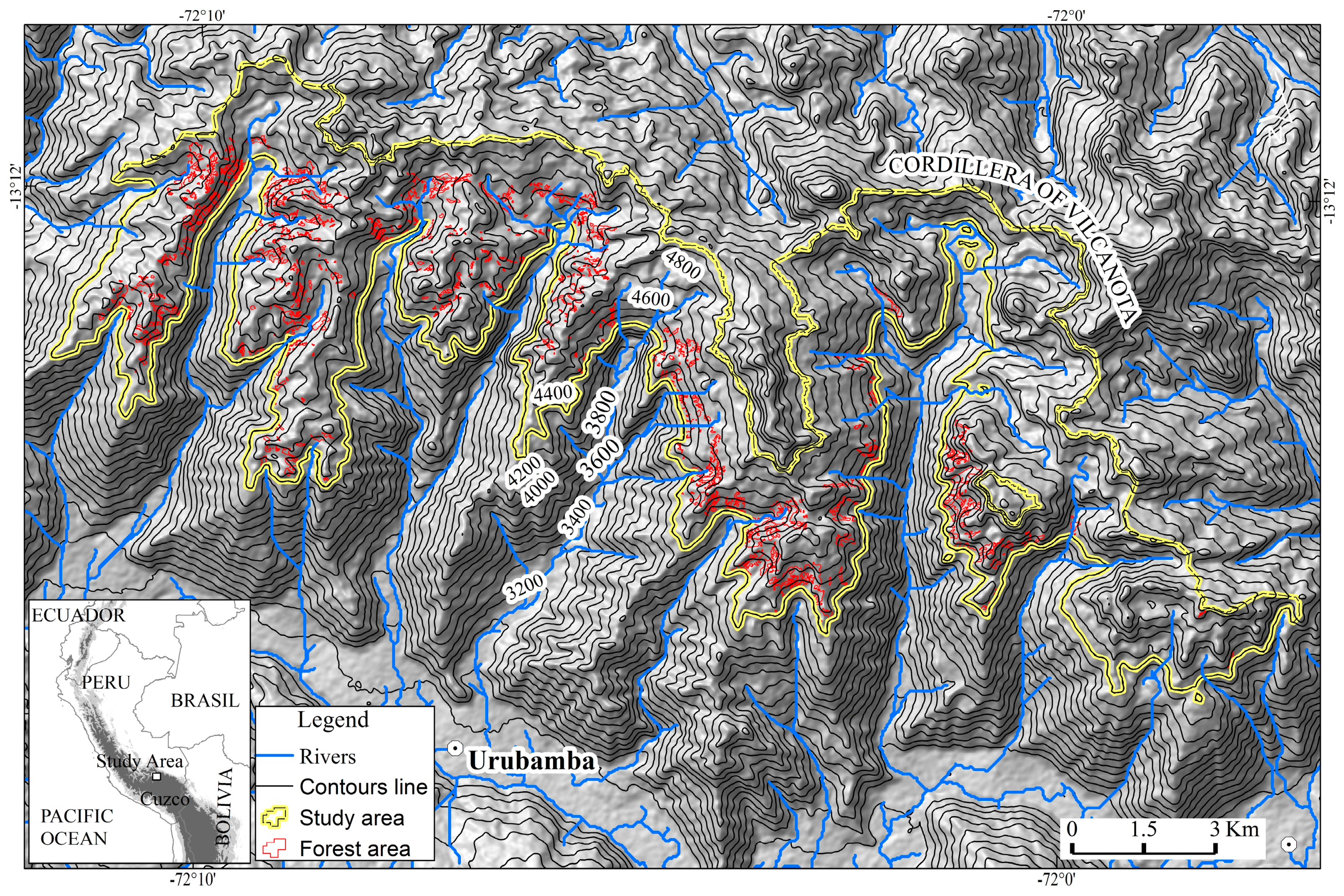
2.1. Study Area
2.2. Data Acquisition and Processing
2.3. Analyses
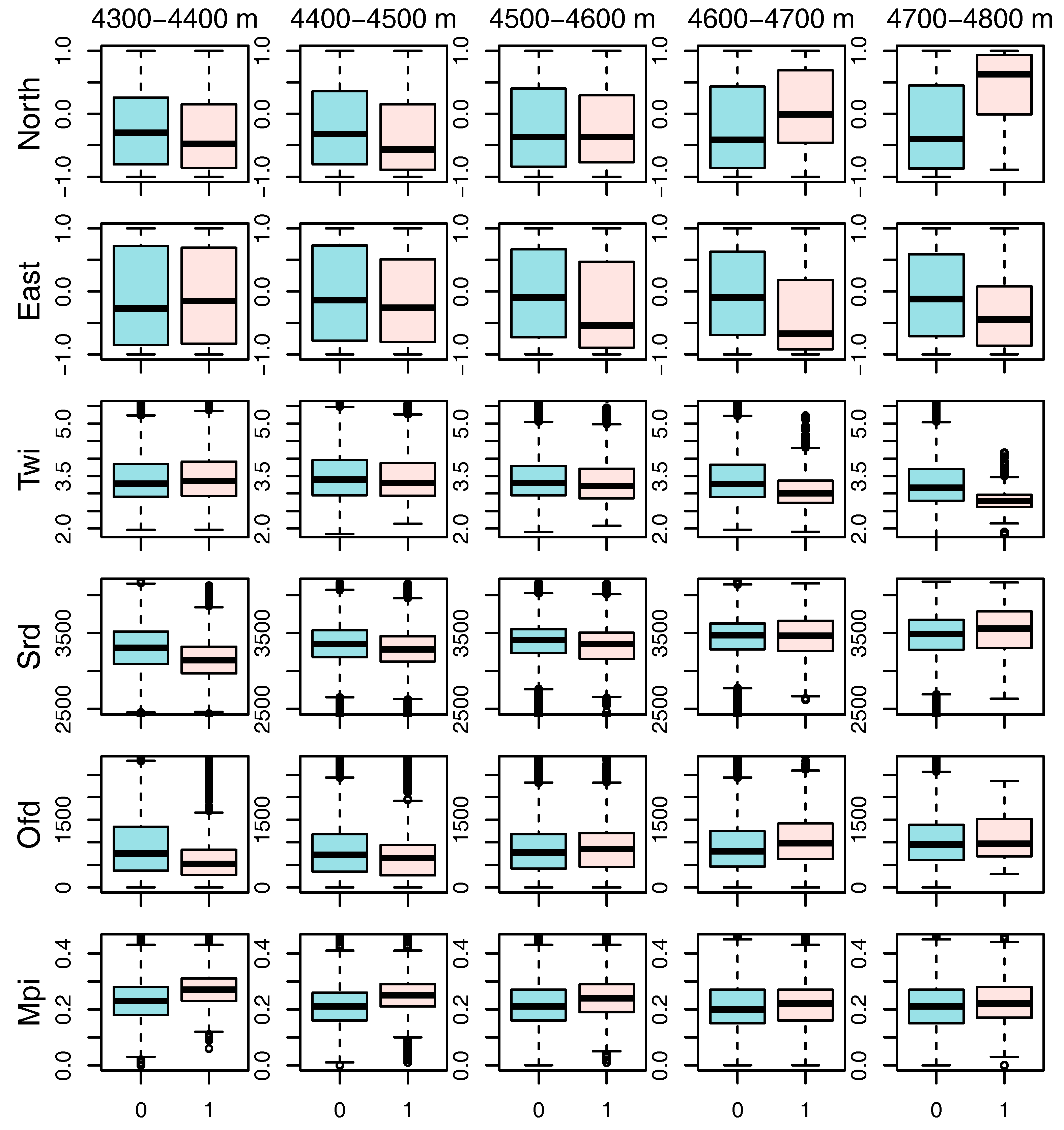
3. Results
4. Discussion
4.1. Topographic Preferences
4.2. Factors other than Topography Determining Polylepis Forest Distribution
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ellenberg, H. Man’s influence on tropical mountain ecosystems in South America. J. Ecol. 1979, 67, 401–416. [Google Scholar] [CrossRef]
- Kessler, M. Present and potential distribution of Polylepis (Rosaceae) forests in Bolivia. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; New York Botanical Garden: Bronx, NY, USA, 1995; pp. 281–294. [Google Scholar]
- Lloyd, H.; Sevillano Ríos, S.; Marsden, S.J.; Valdés-Velásques, A. Bird community composition across Andean tree-line ecotone. Aust. Ecol. 2010, 37, 470–478. [Google Scholar] [CrossRef]
- Toivonen, J.M. Determinants of Polylepis (Rosaceae) Forest Distribution and Treeline Formation in the High Andes. Ph.D. Thesis, University of Turku, Turku, Finland, 27 May 2014. [Google Scholar]
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Kessler, M.; Toivonen, J.M.; Sylvester, S.; Kluge, J.; Hertel, D. Elevational patterns of Polylepis (Rosaceae) tree height in the high Andes of Peru: Role of human impact and climatic conditions. Front. Plant Sci. 2014, 5, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coblentz, D.; Keating, P.L. Topographic controls on the distribution of tree islands in the high Andes of south-western Ecuador. J. Biogeogr. 2008, 35, 2026–2038. [Google Scholar] [CrossRef]
- Zutta, B.R.; Rundel, P.W. Modeled Shifts in Polylepis Species Ranges in the Andes from the Last Glacial Maximum to the Present. Forests 2017, 8, 232. [Google Scholar] [CrossRef]
- Kessler, M.; Böhner, J.; Kluge, J. Modelling tree height to assess climatic conditions at tree lines in the Bolivian Andes. Ecol. Model. 2007, 207, 223–233. [Google Scholar] [CrossRef]
- Gosling, W.D.; Hanselman, J.A.; Knox, C.; Valencia, B.G.; Bush, M.B. Long-term drivers of change in Polylepis woodland distribution in the central Andes. J. Veg. Sci. 2009, 20, 1041–1052. [Google Scholar] [CrossRef]
- Valencia, B.G.; Matthews-Bird, F.; Urrego, D.H.; Williams, J.J.; Gosling, W.D.; Bush, M. Andean microrefugia: Testing the Holocene to predict the Anthropocene. New Phytol. 2016, 212, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Kessler, M. The “Polylepis problem”: Where do we stand? Ecotropica 2002, 8, 97–110. [Google Scholar]
- Renison, D.; Chartier, M.P.; Menghi, M.; Marcora, P.I.; Giorgis, M.; Hensen, I.; Cingolani, A.M. Spatial variation in tree demography associated to domestic herbivores and topography: Insights from a seeding and planting experiment. For. Ecol. Manag. 2015, 335, 139–146. [Google Scholar] [CrossRef]
- Lauer, W. Zur Ökoklimatologie der Kallawaya-Region (Bolivien). Erkunde 1982, 36, 223–247. [Google Scholar] [CrossRef]
- Sarmiento, G. Ecological features of climate in high tropical mountains. In High Altitude Tropical Biogeography; Vuilleumier, F., Monasterio, M., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 11–45. [Google Scholar]
- Dirnböck, T.; Dullinger, S.; Gottfried, M.; Ginzler, C.; Grabherr, G. Mapping alpine vegetation based on image analysis, topographic variables and Canonical Correspondence Analysis. Appl. Veg. Sci. 2003, 6, 85–96. [Google Scholar] [CrossRef]
- Hörsch, B. Modelling the spatial distribution of montane and subalpine forests in the central Alps using digital elevation models. Ecol. Model. 2003, 168, 267–282. [Google Scholar] [CrossRef]
- Brown, D.G. Predicting vegetation types at treeline using topography and biophysical disturbance variables. J. Veg. Sci. 1994, 5, 641–656. [Google Scholar] [CrossRef]
- Braun, G. The use of digital methods in assessing forest patterns in an Andean environment: The Polylepis example. Mt. Res. Dev. 1997, 17, 253–262. [Google Scholar] [CrossRef]
- Bader, M.Y.; Ruijten, J.J.A. A topography-based model of forest cover at the alpine tree line in the tropical Andes. J. Biogeogr. 2008, 35, 711–723. [Google Scholar] [CrossRef]
- Bader, M.Y.; van Geloof, I.; Rietkerk, M. High solar radiation hinders tree establishment above the alpine treeline in northern Ecuador. Plant Ecol. 2007, 191, 33–45. [Google Scholar] [CrossRef]
- Sylvester, S.P.; Heitkamp, F.; Sylvester, M.D.P.V.; Jungkunst, H.F.; Sipman, H.J.M.; Toivonen, J.M.; Gonzales Inca, C.A.; Ospina, J.C.; Kessler, M. Relict high-Andean ecosystems challenge our concepts of naturalness and human impact. Sci. Rep. 2017, 7, 3334. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Kessler, M.; Ruokolainen, K.; Hertel, D. Accessibility predicts structural variation of Andean Polylepis forests. Biodivers. Conserv. 2011, 20, 1789–1802. [Google Scholar] [CrossRef]
- Chepstow-Lusty, A.J.; Bennett, K.D.; Fjeldså, J.; Kendall, A.; Galiano, W.; Tupayachi Herrera, A. Tracing 4000 Years of Environmental History in the Cuzco Area, Peru, from the Pollen Record. Mt. Res. Dev. 1998, 18, 159–172. [Google Scholar] [CrossRef]
- Mosblech, N.A.S.; Chepstow-Lusty, A.; Valencia, B.G.; Bush, M. Anthropogenic control of late-Holocene landscapes in the Cuzco region, Peru. Holocene 2012, 22, 1361–1372. [Google Scholar] [CrossRef]
- Hengl, T.; Reuter, H.I. (Eds.) Geomorphometry: Concepts, Software, Applications; Developments in Soil Science Series; Elsevier: Amsterdam, The Netherlands, 2008; Volume 33, p. 772. [Google Scholar]
- Dormann, C.F.; McPherson, J.M.; Araujo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D.; et al. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Record, S.; Fitzpatrick, M.C.; Finley, A.O.; Veloz, S.; Ellison, A.M. Should species distribution models account for spatial autocorrelation? A test of model projections across eight millennia of climate change. Glob. Ecol. Biogeogr. 2013, 22, 760–771. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Springer: New York, NY, USA, 2007; p. 672. [Google Scholar]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012; p. 220. [Google Scholar]
- Rundel, P.W. Tropical Alpine Climates. In Tropical Alpine Environments: Plant Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 21–44. [Google Scholar]
- Rada, F.; García-Núñez, C.; Boero, C.; Gallardo, M.; Hilal, M.; González, J.; Prado, F.; Liberman-Cruz, M.; Azócar, A. Low-temperature resistance in Polylepis tarapacana, a tree growing at the highest altitudes in the world. Plant Cell Environ. 2001, 24, 377–381. [Google Scholar] [CrossRef]
- Azócar, A.; Rada, F.; García-Núñez, C. Functional characteristics of the arborescent genus Polylepis along a latitudinal gradient in the high Andes. Interciencia 2007, 32, 663–668. [Google Scholar]
- Ball, M.C.; Hodges, V.S.; Laughlin, G.P. Cold-induced photoinhibition limits regeneration of snow gum at tree-line. Funct. Ecol. 1991, 5, 663–668. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K. Sky exposure, crown architecture, and low-temperature photoinhibition in conifer seedlings at alpine treeline. Plant Cell Environ. 1999, 22, 407–415. [Google Scholar] [CrossRef]
- Young, K.R. Tropical timberlines: Changes in forest structure and regeneration between two Peruvian timberline margins. Arct. Antarct. Alp. Res. 1993, 25, 167–174. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Acronym | Variable | Description | Ecological Importance |
|---|---|---|---|
| # North | Northern exposure | Relation to north (1 to −1) | Summer vs. winter solar radiation |
| # East | Eastern exposure | Relation to east (1 to −1) | Morning vs. afternoon solar radiation |
| Sl | Slope | Slope angle in degrees | Solar radiation, terrain stability, soil moisture |
| Pro | Profile curvature | Terrain surface curvature described parallel to slope steepness (convex, linear or concave) | Soil moisture and erosion |
| Pla | Planar curvature | Terrain surface curvature described perpendicularly to the slope steepness (convex, linear or concave). | Solar radiation, wind exposure, soil erosion |
| Convg | Convergenceindex | Overland flow convergence and divergence | Soil hydrological condition and soil erosion |
| # Twi | Topographic wetness index | Soil moisture and surface saturation index, calculation based on the control of local topography on hydrological processes, (ln(As/tanβ) * | Soil moisture, water logging, cold air accumulation |
| # Srd | Annual solar radiation hours | Duration of potential annual direct incoming solar radiation | Solar radiation, annual temperature sum |
| # Ofd | Overland flow distance | Overland distance to the nearest river measured using the DTM-based potential river network | Soil hydrological conditions, topographic protection |
| Vfd | Vertical flow distance | Vertical distance to the nearest river measured using the DTM-based potential river network | Soil hydrological conditions |
| # Mpi | Morphometric protection index | Topographic sheltering, calculation based on pixel values within 300 m radius | Sheltering, solar radiation, temperature extremes |
| Elevation (m) | Polylepis Forest Cover (km2) | Total Landscape (km2) | Polylepis Forest Cover (%) |
|---|---|---|---|
| 4300–4400 | 2.79 | 18.16 | 15.34 |
| 4400–4500 | 3.57 | 20.91 | 17.08 |
| 4500–4600 | 3.26 | 21.42 | 15.22 |
| 4600–4700 | 1.84 | 19.86 | 9.29 |
| 4700–4800 | 0.37 | 15.07 | 2.47 |
| Total | 11.84 | 95.43 | 12.40 |
| 4300–4400 m | Mean Estimate ± S.E. | Mean Std Error | Mean z-Value ± S.E. | Mean Probability |
| (Intercept) | −1.88 ± 0.001 | 0.07 | −26.35 ± 0.008 | 0.000 |
| East | −0.01 ± 0.002 | 0.07 | −0.20 ± 0.028 | 0.526 |
| North | −0.19 ± 0.002 | 0.07 | −2.78 ± 0.029 | 0.041 |
| Twi | −0.02 ± 0.002 | 0.07 | −0.37 ± 0.028 | 0.512 |
| Ofd | −0.30 ± 0.003 | 0.09 | −3.44 ± 0.028 | 0.010 |
| Srd | −0.10 ± 0.003 | 0.10 | −0.99 ± 0.030 | 0.366 |
| Mpi | 0.46 ± 0.003 | 0.09 | 4.94 ± 0.027 | 0.000 |
| 4400–4500 m | Mean Estimate ± S.E. | Mean Std Error | Mean z-Value ± S.E. | Mean Probability |
| (Intercept) | −1.71 ± 0.001 | 0.06 | −27.90 ± 0.004 | 0.000 |
| East | −0.12 ± 0.002 | 0.06 | −1.98 ± 0.028 | 0.130 |
| North | −0.31 ± 0.002 | 0.06 | −5.09 ± 0.028 | 0.000 |
| Twi | −0.14 ± 0.002 | 0.06 | −2.30 ± 0.027 | 0.081 |
| Ofd | −0.31 ± 0.002 | 0.07 | −4.37 ± 0.026 | 0.001 |
| Srd | 0.23 ± 0.003 | 0.08 | 2.77 ± 0.031 | 0.046 |
| Mpi | 0.57 ± 0.002 | 0.08 | 6.94 ± 0.027 | 0.000 |
| 4500–4600 m | Mean Estimate ± S.E. | Mean Std Error | Mean z-Value ± S.E. | Mean Probability |
| (Intercept) | −1.82 ± 0.001 | 0.06 | −29.27 ± 0.005 | 0.000 |
| East | −0.29 ± 0.002 | 0.06 | −4.64 ± 0.029 | 0.001 |
| North | −0.07 ± 0.002 | 0.06 | −1.24 ± 0.028 | 0.310 |
| Twi | −0.10 ± 0.002 | 0.06 | −1.61 ± 0.029 | 0.216 |
| Ofd | 0.08 ± 0.002 | 0.06 | 1.45 ± 0.030 | 0.252 |
| Srd | 0.15 ± 0.003 | 0.09 | 1.76 ± 0.031 | 0.189 |
| Mpi | 0.49 ± 0.003 | 0.09 | 5.56 ± 0.029 | 0.000 |
| 4600–4700 m | Mean Estimate ± S.E. | Mean Std Error | Mean z-Value ± S.E. | Mean Probability |
| (Intercept) | −2.61 ± 0.002 | 0.10 | −27.26 ± 0.017 | 0.000 |
| East | −0.47 ± 0.003 | 0.09 | −5.44 ± 0.029 | 0.001 |
| North | 0.38 ± 0.002 | 0.08 | 4.85 ± 0.025 | 0.000 |
| Twi | −0.55 ± 0.003 | 0.10 | −5.23 ± 0.024 | 0.000 |
| Ofd | 0.25 ± 0.002 | 0.07 | 3.33 ± 0.030 | 0.015 |
| Srd | 0.29 ± 0.004 | 0.12 | 2.38 ± 0.028 | 0.073 |
| Mpi | 0.20 ± 0.003 | 0.12 | 1.63 ± 0.029 | 0.211 |
| 4700–4800 m | Mean Estimate ± S.E. | Mean Std Error | Mean z-Value ± S.E. | Mean Probability |
| (Intercept) | −4.84 ± 0.008 | 0.33 | −14.64 ± 0.032 | 0.000 |
| East | −0.43 ± 0.008 | 0.21 | −1.92 ± 0.031 | 0.155 |
| North | 0.89 ± 0.005 | 0.21 | 4.22 ± 0.012 | 0.000 |
| Twi | −1.32 ± 0.009 | 0.31 | −4.18 ± 0.016 | 0.000 |
| Ofd | 0.10 ± 0.005 | 0.18 | 0.62 ± 0.027 | 0.471 |
| Srd | 0.36 ± 0.008 | 0.29 | 1.21 ± 0.026 | 0.320 |
| Mpi | 0.16 ± 0.008 | 0.28 | 0.57 ± 0.028 | 0.470 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toivonen, J.M.; Gonzales-Inca, C.A.; Bader, M.Y.; Ruokolainen, K.; Kessler, M. Elevational Shifts in the Topographic Position of Polylepis Forest Stands in the Andes of Southern Peru. Forests 2018, 9, 7. https://doi.org/10.3390/f9010007
Toivonen JM, Gonzales-Inca CA, Bader MY, Ruokolainen K, Kessler M. Elevational Shifts in the Topographic Position of Polylepis Forest Stands in the Andes of Southern Peru. Forests. 2018; 9(1):7. https://doi.org/10.3390/f9010007
Chicago/Turabian StyleToivonen, Johanna M., Carlos A. Gonzales-Inca, Maaike Y. Bader, Kalle Ruokolainen, and Michael Kessler. 2018. "Elevational Shifts in the Topographic Position of Polylepis Forest Stands in the Andes of Southern Peru" Forests 9, no. 1: 7. https://doi.org/10.3390/f9010007
APA StyleToivonen, J. M., Gonzales-Inca, C. A., Bader, M. Y., Ruokolainen, K., & Kessler, M. (2018). Elevational Shifts in the Topographic Position of Polylepis Forest Stands in the Andes of Southern Peru. Forests, 9(1), 7. https://doi.org/10.3390/f9010007